
Abstrak
- Keanekaragaman hayati dipengaruhi oleh faktor lokal dan lanskap, namun faktor mana yang paling penting tampaknya bervariasi tergantung pada kelompok spesies dan konteksnya. Memahami bagaimana faktor lingkungan memengaruhi spesies pada skala spasial yang berbeda sangat penting untuk perencanaan konservasi yang efektif. Di sini, kami meneliti pengaruh karakteristik lokal dan lanskap pada populasi kumbang saproksilik dan kumbang yang menyukai api setelah pembakaran yang direncanakan.
- Kumbang ( Coleoptera ) dikumpulkan menggunakan perangkap pencegat terbang di 23 lokasi di tiga wilayah di Swedia. Pembakaran terencana telah dilakukan di lokasi tersebut 4–7 tahun sebelumnya. Kami menganalisis hubungan antara kekayaan spesies, kelimpahan dan komposisi komunitas, dan variabel lingkungan pada skala lokal (lokasi pembakaran) dan lanskap (radius 2, 5, 10 dan 20 km di sekitar lokasi pembakaran). Variabel lokal adalah volume kayu mati, tutupan tajuk dan luas kebakaran. Variabel lanskap meliputi area hutan yang dilindungi, tua (>120 tahun), ditebang habis dan dibakar, dan volume kayu tegakan pinus Skotlandia, cemara Norwegia dan pohon peluruh.
- Secara total, kami mencatat 3094 kumbang saproksilik, yang termasuk dalam 188 spesies. Dari jumlah tersebut, 1153 individu (37 spesies) diklasifikasikan sebagai kumbang yang menyukai api. Tidak ada variabel lokal yang memiliki pengaruh signifikan terhadap kekayaan atau kelimpahan kumbang. Untuk hubungan lanskap, skala 5 dan 10 km paling relevan untuk menganalisis respons kumbang saproksilik. Pada skala ini, kekayaan kumbang saproksilik berasosiasi positif dengan kawasan hutan lindung, sedangkan kelimpahan menunjukkan hubungan negatif. Sebaliknya, kekayaan kumbang saproksilik berasosiasi negatif dengan area tebang habis, sementara kelimpahan spesies saproksilik dan yang menyukai api berasosiasi positif. Kekayaan spesies serupa di seluruh wilayah, sedangkan komposisi spesies bervariasi. Proporsi hutan tua merupakan variabel penting yang berkontribusi terhadap perbedaan regional dalam komposisi spesies.
- Sintesis dan aplikasi. Studi kami menyoroti pentingnya lanskap sekitar untuk membentuk komunitas kumbang saproksilik setelah pembakaran terencana. Pengelolaan hutan dan tindakan konservasi dalam radius 5–10 km dari pembakaran terencana dapat memengaruhi kumpulan kumbang pascapembakaran dan harus dipertimbangkan saat merencanakan pembakaran yang bertujuan untuk mendorong perkembangan kumbang saproksilik. Selain itu, karena komposisi spesies bervariasi di berbagai wilayah, hal ini menunjukkan bahwa pembakaran terencana di berbagai wilayah saling melengkapi.
1. PENDAHULUAN
Bahasa Indonesia: Di ekosistem hutan rawan kebakaran di seluruh dunia, api yang direncanakan digunakan sebagai alat manajemen dengan berbagai aplikasi (Burrows & McCaw, 2013 ; Lindberg et al., 2020 ; Ryan et al., 2013 ). Aplikasi utama di banyak wilayah adalah pengurangan bahaya kebakaran melalui pembakaran yang disengaja untuk mengurangi beban bahan bakar (Boer et al., 2009 ; Hunter & Robles, 2020 ). Di hutan boreal Fennoscandia, aplikasi umum adalah persiapan lokasi untuk regenerasi hutan dalam kehutanan produksi dan dalam konservasi alam sebagai alat restorasi (Cogos et al., 2020 ; Lindberg et al., 2020 ; Ramberg et al., 2018 ).
Pembakaran untuk tujuan konservasi di hutan boreal bertujuan untuk memberi manfaat bagi keanekaragaman hayati dengan memulihkan struktur dan dinamika yang telah berkurang atau terdegradasi akibat praktik pengelolaan kehutanan yang intensif dan penanggulangan kebakaran yang efektif (Granström, 2001 ; Lindberg et al., 2020 ). Luas dan lokasi pembakaran terencana ini terutama ditentukan oleh pertimbangan praktis dan keselamatan. Namun, agar efektif, penting juga untuk mengidentifikasi area di lanskap tempat restorasi akan memberikan manfaat terbesar bagi spesies sasaran dan mengurangi dampak negatif pada spesies yang sensitif terhadap kebakaran.
Tujuan penting dari pembakaran yang direncanakan adalah untuk menciptakan kayu mati, karena kekurangan substrat ini di hutan produksi merupakan ancaman bagi banyak spesies saproksilik (yakni tergantung pada kayu mati) (Siitonen, 2001 ). Kumbang saproksilik bergantung pada kayu mati untuk perkembangan larvanya (Stokland et al., 2012 ), dan banyak dari mereka disukai oleh kebakaran (Bell, 2023 ). Di antara kumbang yang disukai api, beberapa bersifat pirofilik, yakni bergantung pada dan beradaptasi dengan api, sementara yang lain lebih umum beradaptasi dengan gangguan, dengan preferensi untuk habitat terbuka yang disinari matahari (Bell, 2023 ; Wikars, 1997 ). Penelitian sebelumnya telah mengonfirmasi bahwa kebakaran hutan dan pembakaran yang direncanakan mendorong kumbang saproksilik dan kumbang yang disukai api (Hjältén et al., 2018 ; Johansson et al., 2011 ; Wikars, 1997 ). Namun, pengetahuan mengenai bagaimana lanskap sekitar memengaruhi kemunculan spesies setelah kebakaran masih terbatas (lihat Kouki et al., 2012 ; Ranius et al., 2014 ; Rubene et al., 2017 ).
Peningkatan jumlah dan heterogenitas kayu mati di lokasi setempat telah terbukti meningkatkan kelimpahan dan kekayaan kumbang saproksilik, dan memengaruhi komposisi spesies (Larsson Ekström et al., 2021 ; Ranius et al., 2014 ; Seibold et al., 2016 ). Hal ini sesuai dengan hipotesis energi spesies, yang menghubungkan kekayaan dan kelimpahan spesies yang lebih tinggi dengan peningkatan jumlah habitat, karena lebih banyak energi tersedia (Wright, 1983 ), dan dengan hipotesis heterogenitas habitat, yang mengaitkan peningkatan kekayaan spesies dengan peningkatan keanekaragaman habitat, karena lebih banyak relung yang ada (MacArthur & MacArthur, 1961 ; Seibold et al., 2016 ).
Semakin banyak dukungan untuk hipotesis jumlah habitat (HAH), yang menyatakan bahwa jumlah habitat total dalam lanskap lebih penting daripada konfigurasi spasialnya (Fahrig, 2013 ; Watling et al., 2020 ), tetapi lihat juga Gonçalves-Souza et al. ( 2025 ) dan sumber di dalamnya. Lanskap dengan habitat yang lebih sesuai diprediksi akan mendukung lebih banyak spesies, menghasilkan jumlah spesies yang lebih tinggi di lokasi di dalamnya dibandingkan dengan lokasi di lanskap dengan habitat yang kurang sesuai. Untuk kumbang saproksilik, jumlah habitat lanskap setidaknya sama pentingnya dengan jumlah habitat lokal dalam menjelaskan komposisi dan kekayaan spesies (Hallinger et al., 2018 ; Rubene et al., 2017 ; Seibold et al., 2017 ). Hutan yang dilindungi dan hutan tua umumnya memiliki volume kayu mati yang lebih besar daripada hutan yang dikelola dan lebih muda (Kyaschenko et al., 2022 ; Siitonen, 2001 ) dan pada skala lanskap, tipe hutan yang kaya kayu mati ini umumnya berkorelasi positif dengan kekayaan dan kelimpahan spesies saproksilik (Hämäläinen et al., 2023 ; Olsson et al., 2012 ; Stenbacka et al. , 2010 ). Jumlah area tebang habis mungkin juga penting, karena penebangan habis mengubah kondisi habitat bagi kumbang saproksilik, yang menguntungkan spesialis habitat terbuka (Stenbacka et al., 2010 ). Jumlah spesies pohon tertentu di lanskap juga dapat memengaruhi spesies saproksilik (Hallinger et al., 2018 ). Bagi spesies yang menyukai kebakaran, area hutan yang terbakar di lanskap kemungkinan merupakan habitat sumber yang penting, karena banyak spesies ini bereproduksi di pohon-pohon yang sekarat atau mati akibat kebakaran (Wikars, 2006 ). Pengetahuan tentang dampak karakteristik lanskap terhadap kumbang saproksilik setelah pembakaran terencana terbatas, sehingga tidak pasti seberapa penting hal ini bagi keanekaragaman hayati kumbang, dan yang harus dipertimbangkan saat merencanakan lokasi pembakaran konservasi.
Selain itu, untuk membuat keputusan konservasi yang terinformasi tentang pembakaran yang direncanakan, penting untuk memahami skala spasial di mana variabel lanskap memengaruhi spesies yang ditargetkan. Namun, menentukan skala yang tepat untuk kumbang saproksilik merupakan tantangan. Misalnya, skala spasial efek jumlah habitat sangat bervariasi di antara kumbang saproksilik, berkisar dari <100 m (Ranius et al., 2011 ) hingga setidaknya 30 km (Bell, 2023 ; Ranius et al., 2014 ). Skala efek adalah radius di mana komposisi lanskap paling baik memprediksi respons spesies (Jackson & Fahrig, 2012 ). Menguji berbagai skala kemungkinan akan meningkatkan peluang mengidentifikasi skala efek yang relevan. Dalam skala yang lebih besar, perbedaan regional dapat membatasi distribusi spesies, membentuk kumpulan spesies regional. Di Swedia, wilayah sangat bervariasi dalam iklim (Ahti et al., 1968 ) dan riwayat pengelolaan (Josefsson & Östlund, 2011 ; Linder & Östlund, 1998 ), yang juga memengaruhi area hutan alam sisa saat ini (Svensson et al., 2019 ). Secara umum, kekayaan spesies diperkirakan menurun dengan meningkatnya garis lintang (Hillebrand, 2004 ). Untuk kumbang saproksilik, suhu dapat membatasi distribusinya, menciptakan perbedaan regional (Gossmann et al., 2024 ). Perbedaan dalam penggunaan lahan juga dapat menjadi penting, dan wilayah yang kurang dipengaruhi oleh pengelolaan hutan dapat menampung lebih banyak spesies yang masuk daftar merah dan yang disukai kebakaran (Kouki et al., 2012 ). Di Swedia, wilayah utara memiliki sejarah pengelolaan hutan intensif yang lebih pendek dan lebih banyak hutan alami dan hutan yang hampir alami dibandingkan dengan wilayah selatan. Dengan demikian, wilayah utara cenderung memiliki spesies yang lebih langka dan lebih rentan terhadap kebakaran. Tujuan dari penelitian ini adalah untuk mendapatkan pemahaman yang lebih mendalam tentang bagaimana variabel lokal dan lanskap membentuk komunitas kumbang saproksilik setelah terbakar. Kami mengambil sampel kumbang saproksilik di 23 lokasi di Swedia yang telah terbakar 4–7 tahun sebelumnya. Kami memperkirakan kekayaan dan kelimpahan spesies di lokasi terbakar akan meningkat dengan: (1) jumlah habitat lokal (volume kayu mati dan luas kebakaran) dan (2) luas hutan lindung, hutan tua (>120 tahun) dan hutan terbakar—kategori ini tidak saling eksklusif—dan volume pohon hidup per ha di lanskap sekitarnya. Untuk penebangan habis, kami mengharapkan hubungan positif dengan spesies yang disukai api, karena mereka beradaptasi dengan gangguan, tetapi tidak untuk spesies saproksilik secara umum. Kami membandingkan tiga wilayah di Swedia yang berbeda dalam iklim dan riwayat penggunaan lahan. Kami memperkirakan bahwa kekayaan dan kelimpahan spesies saproksilik akan lebih tinggi di wilayah selatan, sementara kelimpahan dan kekayaan spesies yang disukai api akan lebih tinggi di utara.
2 BAHAN DAN METODE
Studi ini dilakukan pada tahun 2022 di tiga wilayah di Swedia: selatan, tengah, dan utara (Gambar 1 ). Delapan lokasi yang terbakar 4–7 tahun sebelum inventarisasi dipilih per wilayah. Semua lokasi berada di dalam kawasan lindung (cagar alam, taman nasional, atau Natura 2000) dan luasnya berkisar antara 6 hingga 40 ha (lihat Tabel S1 ). Satu lokasi di utara luluh lantak selama kerja lapangan sehingga menyisakan 23 lokasi.
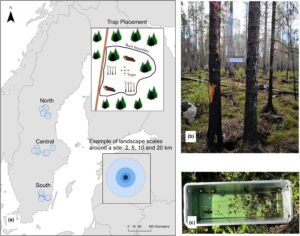
2.1 Survei dan identifikasi spesies
Perangkap intersepsi terbang digunakan karena perangkap tersebut sesuai untuk mengambil sampel kumbang saproksilik, dan perangkap tersebut khususnya berguna ketika tujuannya adalah untuk membandingkan lokasi dan bukan untuk mempelajari item kayu mati tertentu (Ranius & Jansson, 2002 ; Wikars et al., 2005 ). Izin untuk menempatkan perangkap di dalam cagar alam diberikan oleh masing-masing dewan Kabupaten (Gävleborg, Kronoberg, Jönköping, Väster Norrland, Kalmar, Örebro dan Västmanland). Studi ini tidak memerlukan persetujuan etis. Empat hingga lima perangkap ditempatkan di tengah setiap pembakaran, berjarak sekitar 25 m dan di arah yang berbeda (Gambar 1 ; Tabel S1 ). Perangkap tersebut terdiri dari lembaran plastik berukuran 30 × 60 cm dengan baki aluminium (1,8 L) yang dipasang di bawah lembaran tersebut. Baki tersebut setengahnya diisi dengan propilen glikol dan air (50:50) untuk mengawetkan serangga yang jatuh. Perangkap diikat di empat titik dengan tali ke pohon terdekat pada ketinggian sekitar 1,5 m untuk menghindari gangguan satwa liar (Gambar 1 ).
Perangkap tersebut dipasang selama kurang lebih 6 minggu (akhir Mei hingga awal Juli), bertepatan dengan musim puncak aktivitas untuk kumbang saproksilik (Wikars et al., 2005 ). Kami mengecualikan perangkap yang tidak berisi cairan pengawet (dan tidak ada serangga) dari analisis, karena perangkap tersebut dianggap tidak berfungsi, kemungkinan karena angin kencang (Tabel S2 dan S3 ). Kumbang disortir secara manual dari sisa isi perangkap yang distandarisasi menjadi 20 menit per perangkap. Ahli kumbang Hans-Erik Wanntorp, Stockholm, mengidentifikasi kumbang hingga tingkat spesies (Daftar spesies: Tabel S4 ). Spesies kumbang ( Coleoptera ) disortir sebagai saproksilik berdasarkan daftar yang tidak dipublikasikan oleh Mats Jonsell (berdasarkan Hansen, 1964 ; Koch, 1989 ; Palm, 1959 ) dan dikonfirmasi oleh Lars-Ove Wikars. Klasifikasi yang disukai api didasarkan pada Rubene et al. ( 2017 ), dan mencakup spesies yang memiliki hubungan signifikan secara statistik dengan habitat yang terbakar atau telah divalidasi sebagai spesies yang disukai kebakaran oleh para ahli.
2.2 Variabel lokal
Data variabel lokal dikumpulkan pada tahun 2020. Kami mengukur kayu mati kasar yang berdiri dan tumbang (diameter pada tinggi dada ≥10 cm) pada transek sepanjang 50 m, yang disusun dengan jarak 50 m dalam pola kisi yang meliputi lokasi tersebut. Metode transek garis digunakan untuk kayu mati yang tumbang (Van Wagner, 1968 ), yang memerlukan pengukuran diameter kayu mati yang berpotongan dengan garis transek. Volume (dalam m3 / ha) kayu mati yang tumbang dihitung menggunakan persamaan:
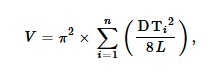
dimana DT i = diameter (dalam dm) item kayu i pada lintasan transek, dan L = panjang lintasan transek (dalam m) (Van Wagner, 1968 ).
Diameter dan tinggi kayu mati yang berdiri diukur dalam jarak 2,5 m pada setiap sisi transek yang sama yang digunakan untuk memperkirakan kayu mati yang tumbang. Volume kayu mati yang berdiri dihitung menggunakan persamaan untuk silinder. Volume kayu mati total, yang berdiri, dan yang tumbang dimasukkan sebagai variabel dalam uji statistik. Tutupan tajuk (%) diukur di atas setiap perangkap menggunakan CanopyApp (Untuk Apple iOS, Versi 1.0.3, University of New Hampshire) karena keterbukaan dapat memengaruhi kumpulan kumbang saproksilik (Seibold et al., 2016 ). Luas dan tahun kebakaran, serta wilayah, juga dimasukkan dalam analisis (Tabel S1 ).
2.3 Variabel lanskap
Pada tingkat lanskap, kami mengumpulkan data mengenai luas hutan yang terbakar (kebakaran hutan liar dan yang ditentukan), hutan lindung, hutan tebang habis, hutan yang berusia lebih dari 120 tahun, dan volume tegakan per hektar pohon hidup. Terjadi tumpang tindih dalam jenis hutan; misalnya, beberapa hutan tua dan hutan yang terbakar juga dilindungi.
Koordinat dan luas semua kebakaran hutan yang dilaporkan bersumber dari Badan Kontingensi Sipil Swedia. Poligon kebakaran hutan (melingkar) dibuat menggunakan koordinat dan luas untuk memperoleh estimasi area kebakaran hutan terkini dalam daerah penyangga. Data pembakaran yang direncanakan diambil dari dewan daerah dan Life Taiga (proyek yang didanai UE yang melakukan pembakaran yang direncanakan), serta pemilik lahan hutan terbesar di Swedia (Sveaskog, Holmen, SCA, dan Stora Enso). Banyak kumbang yang menyukai api dapat ditemukan di lokasi kebakaran langsung setelah kebakaran, tetapi umumnya berkurang seiring waktu (Ranius et al., 2014 ; Wikars, 2006 ). Kami menyertakan data hutan yang terbakar selama tahun 2012–2021, karena kami mempertimbangkan bahwa kebakaran di sekitar lokasi kami (terbakar 2015–2018) berpotensi menjadi sumber penyebaran serangga.
Kawasan hutan lindung (taman nasional, cagar alam, Natura 2000 dan habitat kunci hutan) bersumber dari Badan Perlindungan Lingkungan Swedia dan Badan Kehutanan Swedia. Kawasan hutan yang telah ditebang habis antara tahun 2012 dan 2021 diperoleh dari Badan Kehutanan Swedia. Kawasan hutan yang berusia lebih dari 120 tahun, dan volume tegakan per hektar cemara Norwegia ( Picea abies ), pinus Skotlandia ( Pinus sylvestris ) dan pohon peluruh (resolusi raster 25 × 25 m) didasarkan pada kombinasi citra satelit dan validasi di lapangan oleh program Inventarisasi Hutan Nasional Swedia, dan diunduh dari Peta Hutan SLU ( 2010 ).
Untuk memperhitungkan variasi dalam jumlah habitat dan skala spasial efek, variabel lanskap dihitung untuk lanskap melingkar pada empat skala spasial (radius 2, 5, 10 dan 20 km dari perimeter lokasi kebakaran; Gambar 1 ). ArcGIS (ESRI, 2020 ) digunakan untuk mensintesis dan menghitung data untuk setiap variabel lingkungan pada setiap skala. Untuk lapisan raster (volume pohon per ha dan umur hutan), alat statistik zonal sebagai tabel (ESRI, 2020 ) digunakan untuk menghitung rata-rata per hektar untuk volume pohon dan total luas dalam hektar untuk umur. Untuk lapisan vektor (kebakaran, hutan lindung dan area tebang habis), alat tabulasi area (ESRI, 2020 ) digunakan untuk menghitung luas dalam hektar.
2.4 Analisis statistik
Variabel respons kekayaan dan kelimpahan spesies dihitung dan dianalisis per lokasi untuk semua spesies saproksilik dan khususnya untuk spesies yang menyukai api. Untuk mengatasi masalah apa pun terkait upaya dan efisiensi pengambilan sampel, pengenceran berbasis kelimpahan (paket iNEXT; Hsieh et al., 2024 ) digunakan untuk memperkirakan kekayaan spesies di setiap lokasi, menggunakan lokasi dengan kelimpahan terendah sebagai titik perbandingan (saproksilik: 61, menyukai api: 18). Kelimpahan di setiap lokasi dihitung sebagai kelimpahan rata-rata per perangkap.
Kami meneliti efek variabel lokal dan lanskap (Tabel S5 ) pada kekayaan dan kelimpahan spesies per lokasi menggunakan model linear umum (GLM: Mass; Venables & Ripley, 2002 ). Untuk kekayaan spesies, kami menggunakan model Poisson dengan fungsi log-link. Karena overdispersi, kami menggunakan GLM binomial negatif untuk kelimpahan spesies. Plot diagnostik digunakan untuk menilai kecocokan dan asumsi model (paket DHARMa; Hartig, 2022 ). Korelasi antara beberapa variabel dan skala membuat model inklusif semua tidak dapat diandalkan (Gambar S1–S4 ). Dengan demikian, setiap variabel dan skala dimodelkan secara terpisah. Di satu lokasi, kelimpahan yang disukai api sangat tinggi (Tabel S3 ), yang secara tidak proporsional memengaruhi model, sehingga semua model untuk kelimpahan yang disukai api dijalankan dengan dan tanpa lokasi ini untuk menghindari hasil yang menyesatkan. Model AIC diekstraksi untuk setiap model, dan ∆AIC dihitung untuk model jika beberapa skala signifikan pada p < 0,05 untuk mengidentifikasi skala efek (Jackson & Fahrig, 2012 ). Model dengan ΔAIC < 2 dianggap sama-sama masuk akal (Burnham & Anderson, 2002 ). Luas buffer tumpang tindih terutama pada skala 20 km (Tabel S6 ). Dengan demikian, untuk menguji autokorelasi spasial, koefisien autokorelasi Moran’s I untuk residual model dihitung (paket spdep; Bivand & Wong, 2018 ). Tidak ditemukan autokorelasi (Tabel S7 ).
Pola dalam komposisi spesies untuk semua spesies saproksilik, dan khususnya untuk spesies yang menyukai api, dieksplorasi menggunakan penskalaan multidimensi nonmetrik (NMDS: paket vegan; Oksanen et al., 2020 ). NMDS sangat cocok untuk analisis yang fokusnya adalah pada posisi relatif individu statistik, daripada jarak absolutnya. Jarak Euklides pada data transformasi Hellinger digunakan, yang sesuai untuk data kelimpahan spesies dan mengurangi pengaruh spesies langka (Legendre & Gallagher, 2001 ). Asumsi homogenitas penyebaran diuji (paket Vegan; Oksanen et al., 2020 ). Kecocokan dinilai menggunakan fungsi ‘stressplot’ (paket Vegan; Oksanen et al., 2020 ). Analisis varians multivariat permutasi (PERMANOVA) digunakan untuk menguji perbedaan antarwilayah (paket Vegan; Oksanen et al., 2020 ). Autokorelasi spasial dinilai menggunakan Moran’s I pada sumbu NMDS dan pada residual model PERMANOVA. Sementara sumbu NMDS menunjukkan autokorelasi spasial yang signifikan (Moran’s I , p < 0,05), tidak ada autokorelasi spasial yang terdeteksi dalam residual PERMANOVA, yang menunjukkan bahwa model tersebut secara efektif menangkap variasi spasial (Tabel S7 ). Karena kami juga ingin memeriksa bagaimana variabel lokal dan lanskap dapat memengaruhi komposisi spesies, variabel ini dipasang dalam NMDS sebagai vektor dan signifikansinya dinilai dengan uji permutasi.
Semua analisis statistik dilakukan dengan R 4.3.3 (Tim Inti R, 2024 ).
3 HASIL
Secara total, kami mengumpulkan 3094 individu yang termasuk dalam 188 spesies kumbang saproksilik (Tabel S2 dan S4 ). Dari jumlah tersebut, 1153 individu yang termasuk dalam 37 spesies diklasifikasikan sebagai kumbang yang disukai api (Tabel S3 dan S4 ). Kami mencatat 12 spesies (39 individu) yang masuk dalam daftar merah di Swedia (SLU Artdatabanken, 2020 ) (Tabel S4 ). Empat spesies pirofilik dicatat ( Stephanopachys linearis N = 7, Stephanopachys substriatus N = 1, Euracmaeops septentrionis N = 7 dan Denticollis borealis N = 1).
3.1 Dampak lokal dan lanskap terhadap kekayaan dan kelimpahan
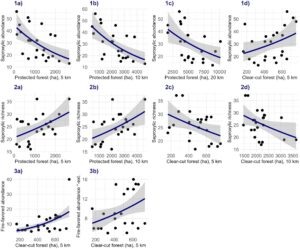
Untuk semua variabel yang berhubungan signifikan dengan variabel respon, skala efek spasial adalah 5 atau 10 km, dengan kedua skala menerima dukungan statistik yang setara (ΔAIC < 2; Tabel 1 ). Kelimpahan saproksilik berhubungan positif dengan area hutan tebang habis tetapi berhubungan negatif dengan area hutan lindung (Gambar 2 (1a–d); Tabel 1 ). Sebaliknya, kekayaan spesies saproksilik berhubungan positif dengan area hutan lindung tetapi berhubungan negatif dengan area hutan tebang habis (Gambar 2 (2a–d); Tabel 1 ). Kelimpahan spesies yang disukai api berhubungan positif dengan area hutan tebang habis, sementara tidak ada hubungan antara kekayaan spesies yang disukai api dan variabel lingkungan apa pun yang diuji. Untuk variabel lokal, tidak ada hubungan signifikan (berdasarkan α = 0,05) yang terdeteksi (Tabel S8 ).
| Variabel respons | Variabel lingkungan | df | penyimpangan | Sisa DF | Deviasi residual | nilai p | Model AIC (∆AIC) |
|---|---|---|---|---|---|---|---|
| Kelimpahan saproksilik | Hutan lindung
Jarak 5 kilometer |
1 | 11.9 | 21 | 23.1 | <0,001 | 176,5 (∆1,7) |
| Hutan lindung
Jarak 10 kilometer |
1 | 14.9 | 21 | 23.2 | <0,001 | 174,8 (∆0) | |
| Hutan lindung
Jarak tempuh 20 kilometer |
1 | 7.5 | 21 | 23.4 | 0,006 | 179,6 (∆4,8) | |
| Hutan yang ditebang habis
Jarak 5 kilometer |
1 | 3.9 | 21 | 23.4 | 0,05 | 182.3 | |
| Kekayaan saproksilik | Hutan lindung
Jarak 5 kilometer |
1 | 6.04 | 21 | 21.9 | 0,01 | 141,4 (∆1,1) |
| Hutan lindung
Jarak 10 kilometer |
1 | 7.1 | 21 | 20.8 | 0,008 | 140,3 (∆0) | |
| Hutan yang ditebang habis
Jarak 5 kilometer |
1 | 4.6 | 21 | 22.1 | 0,03 | 142,6 (∆0) | |
| Hutan yang ditebang habis
Jarak 10 kilometer |
1 | 3.9 | 21 | 22.8 | 0,05 | 143,3 (∆0,7) | |
| Kelimpahan yang disukai api | Hutan yang ditebang habis
5 km* dengan ekstrim |
1 | 11.3 | 21 | 20.9 | <0,001 | 138.9 |
| Hutan yang ditebang habis
5 km* tanpa ekstrim |
1 | 4.7 | 20 | 27.0 | 0,03 | 119.6 |
Catatan : Hanya hasil p ≤ 0,05 yang disertakan. Variabel lanskap dengan pengaruh marjinal dapat ditemukan di Tabel S9 . AIC model dan ∆AIC disertakan untuk memudahkan interpretasi skala efek. Plot efek dapat ditemukan di Gambar 2 .
3.2 Komposisi spesies saproksilik
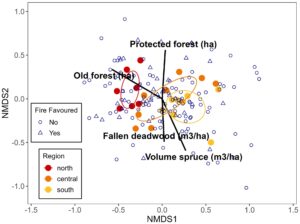
Komposisi spesies kumbang saproksilik berbeda di antara wilayah (Gambar 3 ; PERMANOVA R 2 = 0,18. F 2,22 = 2,24 dan p = 0,001). Pada skala lanskap, luas hutan yang berusia lebih dari 120 tahun penting untuk menjelaskan perbedaan lokasi (Gambar 3 ; Tabel 2 ). Variabel lanskap, luas hutan lindung dan volume pohon cemara juga berpengaruh. Pada skala lokal, jumlah kayu mati yang tumbang penting.
| Skala | Variabel lingkungan | NMDS1 | NMDS2 | R 2 | nilai p |
|---|---|---|---|---|---|
| Lokal | Kayu mati yang tumbang (m3 / ha) | 0,05 | -0,99 | 0,25 | 0,05 |
| 2 kilometer | Hutan lindung (ha) | 0,06 | 0,99 | 0.32 | 0,02 |
| Jarak 5 kilometer | Hutan berumur lebih dari 120 tahun (ha) | -0,80 | 0,60 | 0.43 | 0,003 |
| Jarak 10 kilometer | Hutan berumur lebih dari 120 tahun (ha) | -0,86 | 0,50 | 0,50 | 0,001 |
| Jarak tempuh 20 kilometer | Hutan berumur lebih dari 120 tahun (ha) | -0,85 | 0.51 | 0.45 | 0,004 tahun |
| Volume pohon cemara (m3 / ha) | 0.40 | -0,92 | 0.43 | 0,005 |
Catatan : Nilai dalam kolom NMDS1 dan NMDS2 menunjukkan hubungan variabel lingkungan terhadap sumbu NMDS pertama dan kedua. Nilai p merupakan hasil uji permutasi. Jika variabel lingkungan signifikan pada beberapa skala ( p ≤ 0,05), nilai rata-ratanya dimasukkan ke dalam plot NMDS (Gambar 3 ).
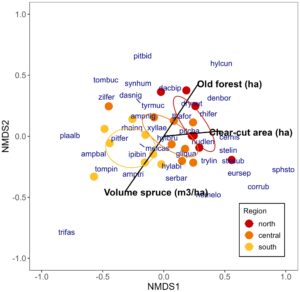
3.3 Komposisi spesies yang menyukai api
Komposisi spesies kumbang yang menyukai api berbeda di antara wilayah (Gambar 4 ; PERMANOVA R 2 = 0,18. F 2,22 = 2,21, p = 0,009). Hutan yang berusia lebih dari 120 tahun merupakan prediktor penting komposisi spesies, terutama pada skala 5 dan 10 km (Tabel 3 ). Volume pohon cemara yang berdiri dan luas hutan yang ditebang habis juga menjelaskan perbedaan lokasi.
| Skala | Variabel lingkungan | NMDS1 | NMDS2 | R 2 | nilai p |
|---|---|---|---|---|---|
| Jarak 5 kilometer | Hutan berumur lebih dari 120 tahun (ha) | 0.51 | 0,85 | 0.40 | 0,005 |
| Jarak 10 kilometer | Hutan berumur lebih dari 120 tahun (ha) | 0,55 | 0.83 | 0.41 | 0,004 tahun |
| Volume pohon cemara (m3 / ha) | -0,70 | -0,71 | 0.26 | 0,04 | |
| Jarak tempuh 20 kilometer | Hutan berumur lebih dari 120 tahun (ha) | 0.54 | 0.83 | 0,35 | 0,02 |
| Volume pohon cemara (m3 / ha) | -0,57 | -0,82 | 0.38 | 0,008 | |
| Hutan tebang habis (ha) | 0,99 | 0,09 | 0.27 | 0,04 |
Catatan : Nilai dalam kolom NMDS1 dan NMDS2 menunjukkan hubungan variabel lingkungan terhadap sumbu NMDS pertama dan kedua. Nilai p adalah hasil uji permutasi. Jika variabel lingkungan memiliki efek signifikan secara statistik pada beberapa skala, nilai rata-ratanya dimasukkan ke dalam plot NMDS (Gambar 4 ).
4 DISKUSI
Hasil kami menunjukkan bahwa jumlah habitat lanskap tampaknya memiliki efek yang lebih kuat pada pembentukan komunitas kumbang saproksilik setelah pembakaran yang ditentukan daripada jumlah habitat lokal. Ini sesuai dengan hipotesis jumlah habitat (Fahrig, 2013 ), yang telah didukung dalam studi kumbang saproksilik (Seibold et al., 2017 ). Ada hubungan positif antara luas hutan lindung di lanskap sekitarnya dan kekayaan kumbang saproksilik di lokasi pembakaran. Hutan lindung yang tidak dikelola menampung kekayaan dan kelimpahan spesies saproksilik yang lebih besar dibandingkan dengan hutan produksi dan tebang habis (Paillet et al., 2010 ; Stenbacka et al., 2010 ), mungkin karena volume dan keanekaragaman kayu mati yang lebih besar dibandingkan dengan matriks hutan produksi di sekitarnya (Kyaschenko et al., 2022 ; Siitonen, 2001 ). Hal ini menyoroti pentingnya menjaga atau meningkatkan jumlah habitat hutan lindung di lanskap untuk konservasi kumbang saproksilik, karena area ini dapat berfungsi sebagai habitat sumber untuk kolonisasi pasca-kebakaran.
Berbeda dengan efek positif kawasan hutan lindung pada kekayaan spesies, kami menemukan hubungan negatif antara tebang habis dan kekayaan spesies. Yang sebaliknya berlaku untuk kelimpahan; kelimpahan kumbang saproksilik menurun dengan luas hutan lindung tetapi meningkat dengan meningkatnya luas tebang habis di lanskap. Juga, untuk spesies yang disukai api, kelimpahan berasosiasi positif dengan area tebang habis. Tebang habis adalah lingkungan terbuka, terang matahari dengan volume kayu mati berdiameter lebih kecil yang relatif tinggi. Studi sebelumnya telah menemukan bahwa tebang habis memiliki kekayaan spesies yang lebih rendah daripada hutan yang berdiri dan memiliki komunitas yang berbeda yang didominasi oleh spesialis yang beradaptasi dengan gangguan (termasuk yang disukai api) dan habitat terbuka (Johansson et al., 2007 ; Stenbacka et al., 2010 ). Proporsi tebang habis di lanskap kami pada skala 5 dan 10 km berkisar antara 2% hingga 10%. Proporsi tinggi penebangan habis-habisan baru-baru ini mungkin menunjukkan lanskap produksi intensif, yang berpotensi menyebabkan berkurangnya jumlah spesies, yang dapat menjelaskan berkurangnya kekayaan di area dengan lebih banyak penebangan habis-habisan. Pola kelimpahan kemungkinan besar disebabkan oleh kesamaan habitat antara lokasi pembakaran dan penebangan habis-habisan, karena keduanya mungkin mendukung spesies yang beradaptasi dengan gangguan tertentu. Misalnya, Hylastes brunneus adalah spesies yang beradaptasi dengan gangguan yang melimpah dalam penelitian kami. Spesies ini memakan akar pohon pinus yang baru mati, yang melimpah di penebangan habis-habisan dan di beberapa lokasi pembakaran kami.
Meskipun kami mempelajari efek komposisi lanskap hingga 20 km di sekitar lokasi kebakaran, efek terkuat terlihat dalam jarak 5 dan 10 km, yang menunjukkan bahwa ini adalah rentang skala spasial di mana struktur lanskap paling baik memprediksi respons kumpulan kumbang saproksilik. Namun, karena skala respons spasial sangat bervariasi di antara spesies saproksilik (Ranius et al., 2011 , 2014 ), skala efek ini harus dilihat dalam konteks komunitas secara keseluruhan dan bukan untuk spesies tertentu. Kami menyarankan bahwa lanskap dengan radius 5–10 km harus dipertimbangkan saat merencanakan pembakaran yang direncanakan dalam konteks kumbang saproksilik dan kumbang yang menyukai api.
Bertentangan dengan harapan kami, kami tidak menemukan hubungan antara kekayaan atau kelimpahan kumbang saproksilik atau kumbang yang menyukai api dan area hutan yang terbakar atau tua. Dalam jarak 10 km dari lokasi kami, proporsi rata-rata hutan yang terbakar adalah 0,4% dan untuk hutan tua 1%. Mungkin, kurangnya hubungan tersebut mungkin karena jenis hutan ini terlalu langka di lanskap untuk secara signifikan memengaruhi kumbang di lokasi kebakaran kami. Studi lain menunjukkan bahwa kekayaan spesies saproksilik secara umum berhubungan positif dengan proporsi hutan tua (Hämäläinen et al., 2023 ; Olsson et al., 2012 ) dalam lanskap yang mirip dengan milik kami. Namun, studi-studi ini berfokus pada spesies atau interaksi lain dan bukan pada kumbang saproksilik atau kumbang yang menyukai api.
Kami menemukan perbedaan yang jelas dalam komposisi spesies di antara wilayah, seperti yang diharapkan, tetapi tidak ada perbedaan dalam kekayaan spesies antar wilayah, yang tidak terduga. Secara umum, kekayaan spesies diharapkan lebih tinggi di lintang yang lebih rendah, mengikuti gradien keanekaragaman lintang (Hillebrand, 2004 ). Hasil kami mungkin menunjukkan bahwa perbedaan dalam penggunaan lahan saat ini dan historis serta rentang iklim kumbang (Gossmann et al., 2024 ) lebih penting dalam membentuk komunitas kumbang saproksilik, yang menghasilkan komposisi spesies yang berbeda tetapi kekayaan yang sama di seluruh wilayah.
Kami menemukan bahwa area hutan tua di lanskap, yang tertinggi di wilayah utara, merupakan variabel penting yang menjelaskan komposisi kumbang saproksilik dan kumbang yang menyukai api. Ini mendukung temuan sebelumnya bahwa proporsi hutan tua di lanskap penting untuk membentuk komposisi spesies kumbang saproksilik (Olsson et al., 2012 ). Komposisi spesies di lokasi selatan dikaitkan dengan volume kayu mati lokal yang tinggi dan volume pohon cemara yang tinggi di lanskap. Ini kemungkinan mencerminkan adaptasi kumpulan spesies terhadap proporsi pohon cemara yang lebih tinggi di selatan dibandingkan dengan utara.
Riwayat penggunaan lahan juga dapat berdampak pada pembentukan komunitas kumbang kita, dengan wilayah utara memiliki riwayat pengelolaan yang lebih pendek dan dengan demikian hutan alam yang lebih utuh (Svensson et al., 2019 ). Kouki et al. ( 2012 ) menemukan bahwa penurunan kekayaan spesies dari Finlandia timur ke barat diimbangi dengan peningkatan intensitas pengelolaan. Meskipun kami tidak menemukan perbedaan kekayaan antarwilayah, mayoritas spesies pirofilik ditemukan di wilayah utara, sementara tidak ada yang tercatat di wilayah selatan. Gradien selatan-utara kami mencakup perbedaan iklim serta penggunaan lahan saat ini dan historis, yang semuanya dapat memengaruhi komposisi komunitas. Karena faktor-faktor ini bervariasi bersama dengan gradien lintang, efek individualnya dan kontribusinya masing-masing terhadap pola yang diamati sulit untuk dipisahkan. Namun demikian, jelas bahwa wilayah menampung kumpulan spesies yang berbeda, yang menunjukkan bahwa efek seragam dari pembakaran yang direncanakan pada komposisi komunitas tidak dapat diharapkan di antara wilayah, yang penting untuk dipertimbangkan saat merencanakan di mana melakukan pembakaran yang direncanakan.
5 KESIMPULAN
Studi kami menunjukkan bahwa lanskap di sekitarnya memainkan peran penting dalam membentuk komunitas kumbang saproksilik setelah pembakaran terencana. Kekayaan dan kelimpahan spesies, serta komposisi komunitas, bervariasi tergantung pada luas hutan lindung, hutan tua, dan penebangan habis di lanskap sekitarnya. Hal ini menunjukkan bahwa pengelolaan hutan dan tindakan konservasi di lanskap sekitarnya memengaruhi keanekaragaman hayati hutan yang terbakar dan harus dipertimbangkan saat merencanakan pembakaran yang bertujuan untuk meningkatkan populasi kumbang saproksilik. Karena kami menemukan bahwa lanskap dengan radius 5–10 km merupakan skala yang paling relevan untuk memprediksi respons kumpulan kumbang saproksilik, kami menyarankan agar setidaknya skala ini diperhitungkan saat merencanakan pembakaran terencana. Pembakaran terencana dalam studi ini dilakukan di hutan lindung, dan hutan lindung di sekitarnya memengaruhi kekayaan dan komposisi spesies yang diamati. Oleh karena itu, pembakaran terencana dapat dilihat sebagai salah satu alat di antara alat lainnya dalam konservasi skala lanskap, dengan, misalnya, melestarikan hutan di cagar alam juga menjadi penting. Terakhir, karena komposisi spesies bervariasi antarwilayah tetapi bukan kekayaannya, kami tidak menemukan bukti jelas yang menunjukkan bahwa satu wilayah harus diprioritaskan daripada wilayah lainnya. Variasi dalam kumpulan spesies di berbagai wilayah sebaliknya menunjukkan bahwa pembakaran yang direncanakan di berbagai wilayah saling melengkapi.


