
Abstrak
- Tumbuhan menyusun mikrobioma rizosfer yang bermanfaat melalui mekanisme ‘teriakan minta tolong’ saat patogen atau serangga herbivora menyerang. Jamur mikoriza arbuskular (AMF) dapat memengaruhi komposisi komunitas mikroba di rizosfer tumbuhan. Namun, dampaknya pada mikrobioma rizosfer sebagai respons terhadap serangan patogen dan serangga herbivora masih belum diketahui.
- Dalam penelitian ini, mesocosm alfalfa, Rhizophagus intraradices , Phoma medicaginis dan Acyrthosiphon pisum digunakan untuk menjelaskan bagaimana AMF mengubah mikrobioma rizosfer tanaman dan responsnya terhadap berbagai tekanan biotik.
- AMF secara signifikan meningkatkan aktivitas fosfatase asam tanah, urease, dan β-glukosidase, yang menyebabkan peningkatan penyerapan fosfor tanah oleh tanaman. Enzim tanah juga berkorelasi positif dan signifikan dengan sifat tunas tanaman, termasuk polifenol oksidase dan peroksidase, asam salisilat (SA), dan penghambat tripsin (TI). Meskipun semua AMF, P. medicaginis, dan kutu daun mengubah mikrobioma rizosfer alfalfa, AMF merekrut mikroorganisme bermanfaat di rizosfer alfalfa sebagai respons terhadap serangan patogen dan kutu daun. Interaksi ganda juga meningkatkan kompleksitas dan stabilitas jaringan mikroba rizosfer. Selain itu, ada korelasi signifikan antara aktivitas enzim tanah, nitrogen nitrat, TI, SA, biomassa, tingkat keparahan penyakit, dan mikrobioma rizosfer.
- Oleh karena itu, AMF meningkatkan pertahanan tanaman dan profil biokimia tanah serta membantu alfalfa dalam menyusun mikrobioma rizosfer yang bermanfaat sebagai respons terhadap serangan P. medicaginis dan kutu daun. Temuan ini penting untuk memahami interaksi antara spesies lintas batas dalam sistem di atas dan di bawah tanah yang diubah oleh AMF dalam ekosistem alami.
Baca Ringkasan Bahasa Sederhana gratis untuk artikel ini di blog Jurnal.
1. PENDAHULUAN
Dalam ekosistem alami, tanaman menghadapi beberapa tekanan secara bersamaan, termasuk fitopatogen dan serangga herbivora (Fernandez-Conradi et al., 2018 ), yang menimbulkan ancaman terhadap stabilitas dan produktivitas ekosistemnya, efektivitas penyerang, dan struktur jaring makanan (Hauser et al., 2013 ; Tack et al., 2012 ). Patogen tanaman dan serangga herbivora sering kali berbagi tanaman inang yang sama (Fernandez-Conradi et al., 2018 ), yang berpotensi memengaruhi dampak yang lain pada tanaman inang sementara pada saat yang sama merusak inangnya secara parah (Tack et al., 2012 ). Mikroorganisme rizosfer sangat memengaruhi pertumbuhan, kesehatan, dan adaptasi tanaman (Petipas et al., 2021 ; Zhang et al., 2017 ).
Mikrobioma rizosfer sangat penting untuk kinerja dan ketahanan tanaman terhadap serangga hama dan patogen di atas tanah (Berendsen et al., 2018 ; Hubbard et al., 2019 ; Friman, Pineda, et al., 2021 ). Patogen dan hama serangga juga dapat menginduksi rizosfer untuk merekrut mikroorganisme tanah yang bermanfaat untuk membantu tanaman melawan penyakit (Kwak et al., 2018 ; Liu et al., 2024 ) dan hama serangga (Friman, Karssemeijer, et al., 2021 ; Friman, Pineda, et al., 2021 ; Wang, Zhang, Liu, et al., 2024 ), seperti yang telah dilaporkan dalam akar gandum ( Triticum aestivum ) (Liu et al., 2021 ; Yin et al., 2021 ), bit gula ( Beta vulgaris ) (Carrión et al., 2019 ) dan Arabidopsis (Berendsen et al., 2018 ). Selain itu, Panax notoginseng merekrut mikroorganisme rizosfer yang bermanfaat dan meningkatkan kesehatan tanahnya setelah daunnya terinfeksi Alternaria panax (Luo et al., 2022 ), sementara kacang tunggak ( Vigna unguiculata ) yang terinfeksi penambang daun ( Liriomyza trifolii ) merekrut mikroba rizosfer yang terkait dengan metabolisme nitrogen (N) untuk melawan herbivora serangga di atas tanah (Gao et al., 2024 ). Di sisi lain, herbivori oleh serangga pemakan pucuk seperti ulat ngengat ( Plutella xylostella ) dan kutu daun ( Brevicoryne brassicae ) menyebabkan komunitas mikroba rizosfer yang berbeda dalam kubis brussel ( Brassica oleracea ) (Friman, Karssemeijer, et al., 2021 ). Namun, penelitian tentang pengaruh interaksi tanaman-patogen-serangga di atas tanah pada mikrobioma rizosfer terbatas (van Dijk et al., 2022 ). Pemahaman ini penting untuk memahami efek interaksi tanaman dengan berbagai patogen dan serangga hama di lingkungan alami dan dapat menginformasikan strategi yang memanfaatkan mikrobioma tanah untuk pengelolaan hama dan penyakit serangga pertanian berkelanjutan.
Di antara mikrobioma rizosfer, jamur mikoriza arbuskular (AMF) merupakan bagian penting dari mikrobiota tanah, membentuk asosiasi endomikoriza dengan lebih dari 80% tanaman terestrial (Smith & Read, 2008 ). Jamur ini meningkatkan ketahanan tanaman terhadap serangan patogen atau serangga hama dengan menghubungkan ekosistem di atas dan di bawah tanah (Meier & Hunter, 2018 ). AMF, patogen, dan hama hidup berdampingan dalam ekosistem, membentuk interaksi kompleks dengan tanaman inangnya yang memengaruhi produktivitas dan stabilitas sistem ini (Li et al., 2023 ). Ketergantungan AMF simbiosis, patogen jamur, dan herbivori serangga pada tanaman inang untuk sumber daya nutrisinya juga mendorong hubungan kompleks antara pertahanan tanaman di atas tanah, mikroorganisme rizosfer bawah tanah, dan nutrisi tanah, yang menghasilkan interaksi yang rumit. Melalui interaksi rumitnya dengan tanaman inang, AMF meningkatkan penyerapan nutrisi dan membantu tanaman dalam mengurangi dampak dari serangga yang memakan (Balog et al., 2017 ) dan infeksi penyakit daun (Jiang et al., 2017 ). Mereka juga meningkatkan aktivitas enzim pertahanan tanaman dan kadar hormon, sehingga meningkatkan ketahanan tanaman, seperti alfalfa ( Medicago sativa ) dan rumput gandum hitam ( Lolium perenne ) terhadap patogen dan serangga (Guo et al., 2019 ; Li et al., 2018 , 2023 ; Li, Duan et al., 2021 ). AMF juga diketahui memengaruhi kelimpahan dan komposisi komunitas mikroba rizosfer (Emmett et al., 2021 ; Wang, Fang, et al., 2021 ; Zhou et al., 2022 ). Misalnya, pada lingkungan rendah fosfor (P), AMF, Funneliformis mosseae , mempengaruhi jumlah mikroba rizosfer dan perakitan komunitas pada jagung ( Zea mays ) (Zhou et al., 2022 ).
Meskipun penting, penelitian tentang pengaruh AMF pada interaksi tanaman-patogen-serangga di atas tanah terbatas. Juga masih belum diketahui apakah perakitan mikrobioma rizosfer oleh AMF dikaitkan dengan resistensi tanaman terhadap patogen di atas tanah dan serangan serangga hama. Sebagian besar penelitian tentang respons mikroorganisme rizosfer tanaman dan nutrisi tanah terhadap lingkungan biotik juga berfokus pada satu spesies penyerang (Liu et al., 2024 ; Wang, Zhang, Liu, et al., 2024 ). Namun, dalam ekosistem alami, beberapa penyerang patogen dan hama serangga sering kali hidup berdampingan (Bardgett & van der Putten, 2014 ). Selain itu, sedikit penelitian yang mengeksplorasi dampak serangan gabungan pada mikroorganisme rizosfer tanaman, khususnya AMF, sehingga sulit untuk memahami bagaimana jamur AMF mengatur respons mikroorganisme rizosfer bawah tanah terhadap hama dan penyakit, terutama ketika serangga dan penyakit muncul secara bersamaan pada tanaman inang yang sama. Oleh karena itu, memahami efek interaksi tanaman dengan berbagai patogen dan hama serangga di lingkungan alami dapat menginformasikan strategi yang memanfaatkan mikrobioma tanah untuk pengelolaan hama serangga dan penyakit pertanian berkelanjutan (Rosa et al., 2018 ).
Studi ini mengeksplorasi hubungan rumit antara penyakit daun di atas tanah, hama serangga, dan mikrobioma rizosfer bawah tanah yang diubah oleh AMF. Dengan memeriksa perubahan komunitas mikroba rizosfer sebagai respons terhadap inokulasi AMF dan tantangan yang ditimbulkan oleh penyakit daun dan hama serangga, kami mengungkap mekanisme yang digunakan jamur untuk menghubungkan proses di atas dan di bawah tanah, mengatur kerusakan dari ancaman ini sambil meningkatkan kesehatan tanaman. Kami berhipotesis bahwa AMF ( Rhizophagus intraradices ) dapat berinteraksi dengan Phoma medicaginis dan kutu daun untuk mengubah keanekaragaman dan struktur komunitas mikroorganisme rizosfer tanaman dan merekrut mikroorganisme rizosfer yang bermanfaat yang meningkatkan pertumbuhan tanaman dan meningkatkan ketahanan terhadap penyerang. Efek ini berkorelasi dengan perubahan sifat tanah bawah tanah dan sifat pucuk tanaman di atas tanah. Temuan kami akan berkontribusi pada pemahaman yang lebih luas tentang interaksi antara organisme di atas tanah dan di bawah tanah dan membuka jalan bagi pendekatan inovatif dalam pengendalian stres biologis dan pertanian berkelanjutan.
2 BAHAN DAN METODE
2.1 Tanaman dan jamur AM
Benih alfalfa cv. Longdong diperoleh dari Pusat Pengujian Benih Makanan Ternak dan Herba Kementerian Pertanian dan Urusan Pedesaan di Tiongkok, sementara AMF, R. intraradices diperoleh dari Bank Glomeromycota di Beijing, Tiongkok. Inokulum untuk AMF disiapkan dengan membudidayakannya pada Trifolium repens dalam pot di rumah kaca dan terdiri dari tanah kering yang mengandung spora dan segmen akar yang terkolonisasi.
2.2 Phoma medicaginis dan Acyrthosiphon pisum
Patogen yang digunakan dalam penelitian ini adalah Phoma medicaginis , yang menyebabkan penyakit bercak daun dan batang hitam musim semi pada alfalfa (Li et al., 2023 ). Patogen diisolasi dari tanaman alfalfa yang terinfeksi di ladang di Chifeng, Mongolia Dalam, Tiongkok dan dikultur pada agar dekstrosa kentang (PDA). Konidia P. medicaginis kemudian dipanen pada PDA dan digunakan untuk mengonfirmasi identitas jamur secara morfologis dan menggunakan teknik molekuler. Di sisi lain, klon kutu daun, Acyrthosiphon pisum yang dikumpulkan dari ladang alfalfa Kampus Yuzhong Universitas Lanzhou, Lanzhou, Tiongkok, dipelihara dalam inkubator, dijaga pada suhu 23°C dengan 16 jam dalam cahaya dan 21°C dalam gelap selama 8 jam pada bibit alfalfa yang diganti setiap minggu. Penelitian ini tidak memerlukan persetujuan etis.
2.3 Desain Eksperimen
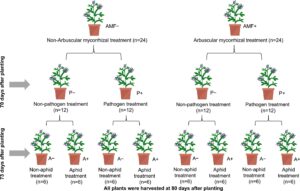
Percobaan rumah kaca dilakukan dengan menanam tanaman alfalfa dari biji dengan AMF (AMF+) atau tanpa inokulum mikoriza (AMF−), dengan (P+) atau tanpa patogen (P-), dan dengan (A+) atau tanpa kutu daun (A−) menggunakan desain faktorial delapan perlakuan. Delapan perlakuan tersebut meliputi: (1) AMF+ diinokulasi tanpa infeksi patogen dan infestasi kutu daun (AMF+P−A−), (2) AMF− tanpa infeksi patogen dan infestasi kutu daun (AMF−P−A−) sebagai kontrol, (3) AMF+ dengan infestasi kutu daun tanpa infeksi patogen (AMF+P−A+), (4) AMF− dengan infestasi kutu daun tanpa infeksi patogen (AMF−P−A+), (5) AMF+ dengan infeksi patogen tanpa infestasi kutu daun (AMF+P+A−), (6) AMF− dengan infeksi patogen tanpa infestasi kutu daun (AMF−P+A−), (7) AMF+ dengan infeksi patogen dan infestasi kutu daun (AMF+P+A+) dan (8) AMF− dengan infeksi patogen dan infestasi kutu daun (AMF−P+A+). Setiap perlakuan diulang enam kali (pot).
Oxydol 10% (Sinopharm Chemical Reagent Co., Ltd., Shanghai, Tiongkok) digunakan untuk mensterilkan benih alfalfa selama 5 menit, diikuti dengan pencucian menyeluruh tiga kali dengan air suling steril dan perkecambahan pada kertas saring basah dalam kegelapan pada suhu 25°C selama 48 jam. Setiap pot yang berisi AMF diinokulasi dengan 50 g inokulum R. intraradices , sedangkan yang tidak berisi (AMF−) menerima 20 mL filtrat mikroba (30 μm), seperti yang telah dijelaskan (Gao et al., 2018 ). Media pertumbuhan adalah campuran 25% tanah (dikumpulkan dari horizon 0–20 cm ladang alfalfa di Universitas Lanzhou, Tiongkok) dan 75% pasir, diayak dengan saringan 2 mm dan diautoklaf pada suhu 121°C selama 1 jam dua kali selama 3 hari, diikuti dengan pengeringan pada suhu 110°C selama 36 jam. Setelah benih berkecambah, pilih bibit dengan ukuran seragam untuk dipindahkan. Bibit kemudian dipindahkan ke pot individu, empat tanaman per pot, masing-masing dengan media pertumbuhan 3500 g dengan atau tanpa inokulum AMF. Tanah dicampur dengan air dalam rasio 1:2 dan kemudian disaring menggunakan saringan 30 μm untuk mendapatkan bubur dan 60 mL digunakan untuk melengkapi media pertumbuhan di setiap pot seperti yang dijelaskan sebelumnya (Johnson et al., 2010 ). Tanaman alfalfa yang terinfeksi patogen disemprot dengan 20 mL 6 × 106 konidia /mL P. medicaginis , sementara tanaman yang tidak diobati patogen disemprot dengan 20 mL air steril. Semua pot ditutup dengan kantong plastik hitam selama 48 jam pasca-inokulasi. Insidensi dan indeks penyakit (DI, Metode Informasi Pendukung S1 ) dinilai 10 hari kemudian. Tanaman alfalfa yang diberi makan kutu daun diinokulasi dengan 12 serangga yang dikurung dalam kandang plastik (tinggi: 35 cm, diameter atas: 10 cm, diameter bawah: 16 cm) dengan ventilasi kain organdy selama 7 hari sebelum memanen alfalfa (Gambar 1 ; Tabel S1 ).
2.4 Panen dan pengukuran tanaman
Tanaman alfalfa dipanen pada usia 12 minggu. Tanaman yang tumbuh di setiap pot digabungkan menjadi satu sampel. Tunas dan akar dipisahkan, dan berat segar tunas dan berat kering akar dicatat, sementara tunas dengan gejala bercak daun digunakan untuk mengisolasi patogen. Sekitar 0,4 g setiap tunas segar digunakan untuk menentukan aktivitas asam jasmonat (JA) dan asam salisilat (SA), penghambat tripsin (TI) dan total fenol (TP) atau aktivitas enzim pertahanan, termasuk katalase (CAT), fenilalanina amonialiase (PAL), polifenol oksidase (PPO), dan peroksidase (POD) seperti yang dijelaskan sebelumnya (Li, Duan et al., 2021 ; Li et al., 2023 ). Bahan tunas yang tersisa digunakan untuk menghitung persentase bahan kering tunas dan berat kering. Kolonisasi AMF pada akar dinilai menggunakan metode intersect garis grid (Giovannetti & Mosse, 1980 ). Singkatnya, jaringan akar dipotong sepanjang 1 cm, dibersihkan dalam KOH 10% dan diwarnai dengan trypan biru 1%.
Tanah rizosfer dikumpulkan seperti yang dijelaskan sebelumnya oleh McPherson et al. ( 2018 ) (lihat Informasi Pendukung: Metode S2 ). pH tanah diukur dalam suspensi tanah-air 1:2,5 menggunakan pH meter (PHS-3C, Shanghai Yidian China). Total fosfor (TP) tanah (sTP), pucuk (shTP) dan akar (rTP) ditentukan dengan metode kolorimetri molibdat-biru menggunakan peralatan smartchem (SMARTCHEM 450, Beijing Lijia China) setelah pencernaan dengan H2SO4 pekat . Kalium yang tersedia di tanah (AK) diekstraksi dengan 1,0 mol/L NH4OAc pada pH 7,0 dan diukur dengan fotometri nyala (Model 410 Flame Photometer, Sherwood Scientific, United Kingdom). Fosfor yang tersedia (AP) diekstraksi dengan 0,5 mol/L NaHCO3 dan ditentukan menggunakan metode kolorimetri molibdat-biru. Nitrogen nitrat (NO 3 − -N), nitrogen amonium (NH 4 + -N) dan enzim tanah, termasuk fosfatase asam (S-ACP), β-glukosidase (S-β-GC), urease (S-UE) dan katalase (S-CAT), diuji menggunakan kit komersial (Suzhou Grace Biotechnology Co., Ltd., Suzhou, Cina), mengikuti petunjuk produsen.
2.5 Sequencing berthroughput tinggi
DNA genom diekstraksi dari 48 sampel tanah rizosfer (enam kali ulangan per perlakuan) menggunakan Kit DNA Tanah Magnetik TGuide S96 (Tiangen Biotech Co., Ltd., Beijing, Tiongkok) mengikuti petunjuk pabrik pembuatnya. Kualitas DNA dinilai melalui elektroforesis pada gel agarosa 1,8%, sedangkan konsentrasi dan kemurniannya diukur menggunakan spektrofotometer NanoDrop 2000 (Thermo Scientific, Wilmington, AS). Wilayah V3-V4 yang sangat bervariasi dari gen 16S rRNA bakteri diamplifikasi menggunakan pasangan primer 338F (5′-ACTCCTACGGGAGGCAGCA-3′) dan 806R (5′-GGACTACHVGGGTWTCTAAT-3′), sementara wilayah internal transcribed spacer 1 (ITS1) dari jamur diamplifikasi dengan pasangan primer F (5′-CTTGGTCATTTAGAGGAAGTAA-3′) dan R (5′-GCTGCGTTCTTCATCGATGC-3′). Produk PCR diperiksa pada gel agarosa, dan dimurnikan menggunakan kit pemurnian DNA Omega (Omega Inc., Norcross, GA, AS) mengikuti petunjuk pabrik. DNA genomik kemudian digabungkan dan diurutkan pada platform Illumina Novaseq 6000 (2 × 250 bp). Kondisi reaksi PCR disediakan dalam Metode S3 .
2.6 Analisis bioinformatika
Pembacaan bersih diklasifikasikan ke dalam varian urutan amplikon (ASV) menggunakan jalur dada2 (Callahan et al., 2016 ). ASV dengan hitungan <2 di semua sampel dihilangkan. Anotasi taksonomi ASV dilakukan menggunakan pengklasifikasi Naive Bayes QIIME2 (Bolyen et al., 2019 ) menggunakan basis data SILVA v138.1 (Quast et al., 2013 ) dengan ambang batas kepercayaan 70%. Indeks keragaman alfa juga dihitung dengan QIIME2 dan divisualisasikan dalam R (Bolyen et al., 2019 ; Wickham & Sievert, 2009 ), sementara keragaman beta dinilai menggunakan QIIME2 tetapi dengan penskalaan multidimensi nonmetrik (NMDS) untuk analisis. Perbedaan taksonomi yang signifikan antar kelompok diidentifikasi menggunakan Linear Discriminant Analysis (LDA) Effect Size (LEfSe) (Segata et al., 2011 ), dengan ambang batas skor LDA logaritmik sebesar 3,5. 2.7 Analisis data Data, termasuk biomassa pucuk, biomassa akar, TI, TP, DI, aktivitas enzim tanaman, hormon tanaman, aktivitas enzim tanah, nutrisi tanah, kolonisasi AMF dan keragaman alfa mikrobioma rizosfer, dianalisis dengan model campuran linier umum faktorial 2 3 standar menggunakan paket LMERTEST dalam R dengan replikasi pot setiap perlakuan sebagai unit eksperimen dan istilah untuk AMF+/AMF−, P+/P− dan A+/A− (dengan interaksi dua dan tiga arahnya). Uji HSD Tukey digunakan untuk menilai signifikansi pemisahan rata-rata dalam SPSS 20.0 (SPSS Inc., Chicago, IL, AS). Perbandingan antara rata-rata didasarkan pada tingkat probabilitas 0,05. Jika rata-rata perlakuan mencakup rentang yang besar, data ditransformasikan secara logaritmik untuk menghindari nilai numerik yang besar yang tidak semestinya memengaruhi perbandingan statistik antara perlakuan. Uji Mantel digunakan untuk menganalisis dampak keragaman tingkat genus rizosfer terhadap aktivitas enzim, hormon tanaman, dan indikator tanah. Jaringan ko-kemunculan mikroba dibuat secara independen untuk setiap perlakuan menggunakan ASV berdasarkan frekuensi ASV dalam sampel ≥4/6 (Li et al., 2022 ). Korelasi kuat berdasarkan koefisien korelasi Spearman ρ > 0,8 atau <0,8 dan tingkat penemuan palsu yang dikoreksi p < 0,05 digunakan untuk membangun jaringan. Untuk menilai kompleksitas jaringan, kami menghitung berbagai metrik karakterisasi, termasuk jumlah total node dan link, koefisien pengelompokan rata-rata, dan derajat rata-rata. Nilai-nilai karakterisasi ini dihitung menggunakan paket ‘igraph’ (Qiao et al., 2024 ), sementara jaringan divisualisasikan menggunakan perangkat lunak Gephi interaktif (v. 0.9.7). Konstruksi model pemodelan persamaan struktural (SEM) dilakukan dengan menggunakan paket ‘piecewiseSEM’ di R.
3 HASIL
3.1 AM kolonisasi mikoriza, pertumbuhan tanaman dan tingkat keparahan penyakit
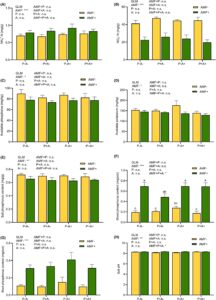
Akar alfalfa yang tidak diinokulasi dengan AMF memiliki sangat sedikit struktur mikoriza yang dapat diamati, tidak seperti yang diinokulasi dengan jamur, yang menunjukkan 46,36%–57,83% dari kolonisasi segmen akar 1 cm (Gambar 2A ). AMF secara signifikan meningkatkan biomassa pucuk dan akar alfalfa ( p < 0,001; Gambar 2B,C ), sementara infeksi P. medicaginis secara signifikan mengurangi biomassa pucuk ( p < 0,001) sebesar 15,16% (Gambar 1a ). AMF juga secara signifikan menurunkan kejadian penyakit dan indeks penyakit, sedangkan infestasi kutu daun meningkatkan kejadian penyakit ( p < 0,05; Gambar 2D,E ).

3.2 Aktivitas enzim pertahanan tanaman
Inokulasi AMF dan pemberian makan kutu daun secara signifikan meningkatkan aktivitas PAL, PPO dan POD ( p < 0,05; Gambar 3A–C ), sementara infeksi P. medicaginis hanya meningkatkan aktivitas PAL dan POD secara signifikan ( p < 0,05; Gambar 3A–C ). Infeksi P. medicaginis (AMF+P+A−), infestasi kutu daun (AMF+P−A+) atau kedua perlakuan (AMF+P+A+) meningkatkan aktivitas PPO pada tanaman alfalfa yang diinokulasi AMF masing-masing sebesar 13,30%, 15,20% dan 14,58%, dibandingkan dengan tanaman yang hanya diobati dengan AMF (AMF+P−A−) (Gambar 3B ). Namun, aktivitas PPO serupa pada semua perlakuan yang tidak diinokulasi AMF. Infeksi P. medicaginis , infestasi kutu daun, atau kedua perlakuan tersebut meningkatkan aktivitas POD pada tanaman alfalfa yang diinokulasi AMF masing-masing sebesar 23,85%, 15,96%, dan 23,80%, dibandingkan pada tanaman yang hanya diinokulasi AMF (Gambar 3C ), tetapi aktivitas POD tidak berbeda secara signifikan pada semua tanaman alfalfa yang tidak diinokulasi AMF yang terinfeksi P. medicaginis dan yang diinfestasi kutu daun. Baik tanaman alfalfa yang tidak diinokulasi AMF atau yang diinfeksi P. medicaginis saja atau yang hanya diinfestasi kutu daun atau yang terpapar kutu daun dan patogen menunjukkan aktivitas PAL yang lebih tinggi dibandingkan pada perlakuan tanpa patogen dan kutu daun. Secara komparatif, tanaman yang diinfestasi kutu daun tanpa infeksi P. medicaginis pada alfalfa yang tidak diinokulasi AMF dan yang diinokulasi memiliki aktivitas PAL yang lebih tinggi dibandingkan perlakuan dengan atau tanpa patogen dan kutu daun dan dengan patogen saja. Namun demikian, AMF, kutu daun dan P. medicaginis tidak secara signifikan mempengaruhi aktivitas CAT (Gambar 3D ).

3.3 Aktivitas fitohormon dan TI serta perubahan kandungan TP
Perlakuan AMF dan patogen secara signifikan meningkatkan konsentrasi SA ( p < 0,001) tetapi menurunkan konsentrasi JA pada tanaman alfalfa ( p < 0,001), sedangkan pemberian makan kutu daun tidak secara signifikan mengubah kedua konsentrasi hormon pada alfalfa yang diinokulasi non- dan AMF (Gambar 3E,F ). Tanaman alfalfa yang diinokulasi AMF yang diinfestasi kutu daun memiliki 34% lebih banyak SA daripada tanaman yang hanya terpapar kutu daun, sedangkan tanaman yang terpapar AMF, kutu daun, dan P. medicaginis memiliki 16,6% lebih banyak SA daripada tanaman yang diobati dengan kutu daun dan hanya P. medicaginis (Gambar 3E ). SA juga lebih tinggi pada tanaman yang diinokulasi AMF yang hanya terinfeksi P. medicaginis atau yang hanya diinfestasi kutu daun, atau yang diobati dengan serangga dan patogen daripada pada tanaman yang hanya diinokulasi AMF. Namun, konsentrasi SA serupa di semua perlakuan yang tidak diinokulasi AMF, kecuali pada tanaman yang terpapar patogen, yang memiliki SA sedikit lebih tinggi. JA pada tanaman yang diinokulasi AMF yang terpapar patogen adalah 16,63% lebih rendah daripada tanaman yang tidak diinokulasi AMF dengan P. medicaginis tetapi 13,19% lebih rendah pada tanaman yang diinokulasi AMF dengan patogen dan kutu daun daripada pada tanaman alfalfa yang diobati dengan patogen dan kutu daun saja (Gambar 3F ). Aktivitas TI meningkat secara signifikan sebesar 35,73% pada tanaman alfalfa yang diinokulasi AMF, sebesar 14,76% pada tanaman yang terinfeksi P. medicaginis dibandingkan dengan perlakuan patogen yang tidak diinokulasi AMF dan tidak terinfeksi, masing-masing ( p < 0,001) (Gambar 3G ). Infestasi kutu daun secara signifikan meningkatkan kandungan TP alfalfa sebesar 11,54% ( p < 0,001; Gambar 3H ).
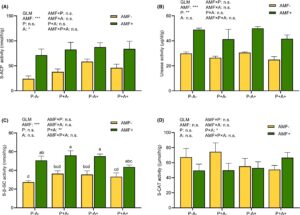
3.4 Aktivitas enzim dan kandungan nutrisi tanah rizosfer
Inokulasi dengan AMF secara signifikan meningkatkan aktivitas S-ACP, S-UE dan S-β-GC pada tanaman alfalfa masing-masing sebesar 95,66%, 62,81% dan 54,75%, dibandingkan pada alfalfa yang tidak terinfeksi AMF ( p < 0,001; Gambar 4A–C ). Infestasi kutu daun secara signifikan meningkatkan ( p = 0,042) aktivitas S-ACP sebesar 27,69%, sedangkan infeksi P. medicaginis secara signifikan menurunkan ( p = 0,008) aktivitas urease sebesar 15,76% (Gambar 4B ). AMF juga secara signifikan meningkatkan kandungan NH 4 + -N ( p < 0,001) tetapi menurunkan kandungan NO 3 − -N ( p < 0,001), AP ( p = 0,024) dan AK ( p = 0,021) (Gambar 5A–D ). Kandungan NH4 + -N dalam tanah 17,53% lebih tinggi pada tanaman yang diinokulasi AMF dibandingkan dengan perlakuan non-AMF. Namun, kandungan NO3 – -N, AP, dan AK dalam tanah pada alfalfa yang diinokulasi AMF menurun masing-masing sebesar 48,10%, 10,45%, dan 10,47% dibandingkan dengan kandungan pada tanaman alfalfa yang tidak diinfeksi AMF (Gambar 5B–D ). Inokulasi dengan AMF juga secara signifikan ( p < 0,001) meningkatkan kandungan fosfor pada pucuk dan akar, serta pH tanah (Gambar 5E–H ), sehingga menghasilkan fosfor yang berkurang secara signifikan dalam tanah tanaman alfalfa yang diolah dengan AMF.
3.5 Analisis korelasi
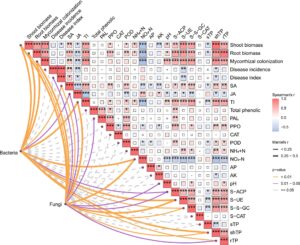
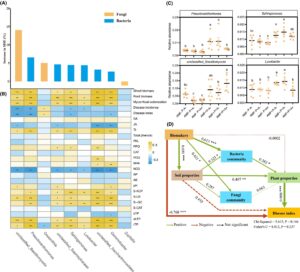
Analisis uji mantel secara signifikan mengkorelasikan biomassa pucuk dan akar, kolonisasi mikoriza, indeks penyakit, SA, JA, TI, PAL, NO 3 − N, pH, S-ACP, S-UE, S-β-GC, TP pucuk dan kandungan TP akar dengan komunitas jamur, sementara biomassa pucuk dan akar, kolonisasi mikoriza, SA, JA, TI, NO 3 − N, S-ACP, S-UE, S-β-GC, TP pucuk dan kandungan TP akar secara signifikan berkorelasi dengan komunitas bakteri (Gambar 6 ). Analisis korelasi mengungkapkan korelasi positif signifikan antara S-ACP dan S-UE, S-β-GC, PAL, PPO dan POD; S-UE dan S-β-GC, NH 4 + N, S-β-GC dan NH 4 + N; dan PPO dan PAL ( p < 0,05). Korelasi negatif yang signifikan juga diamati antara S-ACP dan NO 3 − N; S-UE dan NO 3 − N, S-TP, kejadian penyakit, indeks penyakit; S-β-GC dan NO 3 − N; NO 3 − N dan POD dan PPO ( p < 0,05; Gambar 6 ).
3.6 Keanekaragaman dan struktur komunitas mikrobioma rizosfer
Inokulasi alfalfa dengan AMF secara signifikan menurunkan indeks Shannon jamur ( p = 0,007), Simpson ( p = 0,001) dan Pielou ( p < 0,001) (Gambar 7A ). Tanaman alfalfa yang tidak diinokulasi AMF dan tidak terinfeksi patogen dan kutu daun (kontrol) memiliki indeks Simpson dan Pielou tertinggi, sedangkan tanaman yang terinfeksi AMF dan P. medicaginis memiliki indeks Simpson dan Pielou terendah. Selain itu, indeks Simpson lebih rendah pada alfalfa AMF yang terinfeksi P. medicaginis dibandingkan pada yang hanya diserang oleh satu kutu daun atau oleh kutu daun dan patogen (Gambar 7A ). Paparan alfalfa terhadap AMF, kutu daun dan P. medicaginis tidak secara signifikan ( p > 0,05) mempengaruhi kekayaan ASV jamur, indeks ACE dan Chao1 (Gambar 7A ). Inokulasi AMF meningkatkan secara signifikan ( p < 0,05) ACE bakteri dan kekayaan (Gambar 7B ), sedangkan infestasi kutu daun meningkatkan secara signifikan ( p < 0,05) ACE bakteri, Chao1 dan kekayaan (Gambar 7B ). Indeks Chao1 bakteri meningkat secara signifikan pada tanaman alfalfa yang tidak diinokulasi AMF yang diobati dengan kutu daun dan P. medicaginis dibandingkan dengan kontrolnya yang tidak diobati. Perlu dicatat bahwa alfalfa yang tidak diinokulasi dengan AMF dan bebas dari kutu daun dan patogen memiliki kekayaan, ACE, Chao1 dan indeks Shannon terendah. Selain itu, AMF, kutu daun dan P. medicaginis tidak secara signifikan mempengaruhi indeks Pielou dan Simpson bakteri (Gambar 7B ).

Perbedaan yang signifikan dalam struktur komunitas jamur dan bakteri tanah diamati di rizosfer antara tanaman alfalfa yang diinokulasi non- dan yang diinokulasi AMF ( p < 0,001; Gambar 8a–d ). Struktur komunitas jamur berbeda secara signifikan ( p = 0,045) antara kontrol yang diinokulasi non-AMF dan tanaman yang hanya dihinggapi kutu daun (Tabel S2 ) dan tidak signifikan antara tanaman yang hanya diinfeksi AMF dan tanaman alfalfa yang diinokulasi AMF dan dihinggapi kutu daun ( p = 0,752; Gambar 8c ). Selain itu, infeksi P. medicaginis ( p = 0,036) dan infestasi kutu daun ( p = 0,015) secara signifikan mengubah struktur komunitas bakteri tanah rizosfer pada tanaman alfalfa yang diinokulasi non-AMF, dibandingkan dengan tanaman kontrolnya yang tidak diobati (Gambar 8e,f ; Tabel S3 ). Namun, infeksi P. medicaginis ( p = 0,315) dan pemberian makan serangga ( p = 0,361) tidak mengubah struktur komunitas bakteri tanah rizosfer pada tanaman alfalfa yang diinokulasi AMF secara signifikan (Gambar 8e,f ; Tabel S3 ).
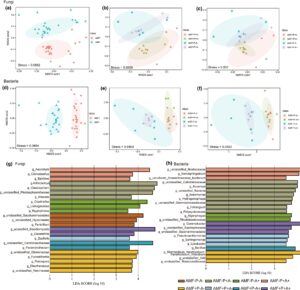
3.7 Komposisi jamur dan bakteri rizosfer
Jamur rizosfer dalam kelompok alfalfa yang berbeda didominasi oleh Ascomycota dan Basidiomycota, dengan Basidiomycota menyumbang lebih banyak jamur pada tanaman yang terinfeksi P. medicaginis dan yang terinfestasi kutu daun pada perlakuan AMF (20,20%), dan pada tanaman alfalfa yang tidak diinokulasi AMF (7,10%) (Gambar S2 ). Glomeromycota menyumbang proporsi jamur yang lebih tinggi pada tanaman alfalfa yang diobati dengan AMF, sedangkan perlakuan P. medicaginis dan kutu daun menurunkan proporsinya. Filum Glomeromycota adalah yang paling melimpah (4,80%) pada tanaman alfalfa yang hanya diinokulasi dengan AMF. Genus Cladosporium menyumbang proporsi jamur yang lebih tinggi pada alfalfa yang tidak diinokulasi AMF dibandingkan dengan alfalfa yang diobati dengan AMF (Gambar S2 ). Bakteri rizosfer dalam berbagai kelompok yang diberi perlakuan alfalfa didominasi oleh Proteobacteria dan Bacteroidota (Gambar S2 ). Hasil penelitian menunjukkan bahwa Actinobacteriota menyumbang proporsi bakteri yang lebih tinggi dalam perlakuan alfalfa AMF, dibandingkan dengan tanaman yang tidak diinokulasi AMF. Selain itu, Gemmobacter menyumbang proporsi bakteri yang lebih tinggi dalam perlakuan alfalfa AMF, dengan 4,09% pada tanaman yang terinfeksi P. medicaginis dan diberi makan kutu daun dalam perlakuan AMF dan 0,22% pada tanaman alfalfa yang hanya diberi makan kutu daun (Gambar S2 ).
Kami juga menggunakan analisis LESfe untuk mengidentifikasi genus karakteristik dalam perlakuan yang berbeda. Hasilnya menunjukkan bahwa genus Zopfiella diperkaya secara signifikan pada tanaman alfalfa yang diinokulasi dengan AMF dan terinfeksi dengan P. medicaginis , sementara Geosmithia dan unclassified_Basidiomycota diperkaya secara signifikan pada alfalfa yang diinokulasi AMF yang terpapar P. medicaginis dan kutu daun. Paramyrothecium dan unclassified_Ceratobasidiaceae secara signifikan lebih banyak pada tanaman yang diinokulasi AMF yang hanya diinfestasi dengan kutu daun (Gambar 8g ). Selain itu, genus Pseudoxanthomonas , Sphingomonas dan Lysobacter diperkaya secara signifikan pada tanaman yang terinfeksi dengan AMF dan P. medicaginis . Gen Gemmobacter secara signifikan lebih banyak pada tanaman alfalfa yang terinfeksi AMF dan P. medicaginis dan diinfestasi kutu daun (AMP+A+), sementara Bacillus diperkaya secara signifikan pada perlakuan AMP-A+ (Gambar 8h ). Khususnya, genus Bacillus diperkaya secara signifikan pada tanaman yang diinokulasi AMF yang hanya diberi makan kutu daun.
3.8 Sifat Topologi Jaringan Mikroba
Kami membangun jaringan mikroba untuk mengeksplorasi potensi interaksi mikroba dalam berbagai perlakuan. Dalam jaringan bakteri, jumlah total simpul dan tautan terendah pada 491 dan 11.962 pada kontrol alfalfa yang tidak diobati dibandingkan dengan perlakuan patogen, kutu daun dan AMF (Gambar 9 ). Infeksi P. medicaginis dan infestasi kutu daun secara signifikan meningkatkan kompleksitas jaringan bakteri pada tanaman alfalfa yang tidak diobati dengan AMF, tidak seperti pada tanaman yang diobati dengan AMF. Namun, dibandingkan dengan alfalfa yang tidak diobati, kompleksitas jaringan bakteri tanah rizosfer tanaman yang diobati dengan AMF lebih kuat. Hasilnya juga menunjukkan bahwa inokulasi AMF secara signifikan meningkatkan stabilitas jaringan bakteri (Gambar S3 ). Meskipun AMF meningkatkan simpul dan tautan jaringan jamur, infeksi patogen dan infestasi kutu daun meningkatkan simpul jaringan jamur pada tanaman alfalfa yang tidak diobati dengan AMF dan AMF. Namun, perlakuan P. medicaginis dan kutu daun menurunkan hubungan pada alfalfa yang tidak diobati dengan AMF tetapi meningkatkan hubungan pada tanaman alfalfa yang diobati dengan AMF (Gambar 9 ), sementara pada saat yang sama secara signifikan meningkatkan stabilitas jaringan jamur (Gambar S3 ). Namun, alfalfa yang terinfeksi dengan AMF dan P. medicaginis memiliki stabilitas jaringan jamur tertinggi, sedangkan mikrobioma rizosfer alfalfa yang diobati dengan AMF memiliki stabilitas jamur yang lebih tinggi daripada alfalfa yang tidak diobati (Gambar S3 ).

3.9 Analisis hutan acak dan model persamaan struktural
Kami menggunakan analisis hutan acak untuk menentukan peringkat genus mikroba yang secara signifikan diperkaya dalam tanaman alfalfa yang diinokulasi AMF yang terinfeksi dengan P. medicaginis atau terpapar pada P. medicaginis dan kutu daun dalam analisis LEfSe, yang menunjukkan bahwa Basidiomycota , Pseudoxanthomonas , Sphingomonas , dan Lysobacter yang tidak terklasifikasi secara signifikan memengaruhi tingkat keparahan penyakit tanaman (Gambar 10A ). Analisis korelasi juga menunjukkan bahwa insiden penyakit tanaman berkorelasi negatif secara signifikan dengan Basidiomycota , Pseudoxanthomonas , Sphingomonas, dan Lysobacter yang tidak terklasifikasi , sementara indeks penyakit berkorelasi negatif secara signifikan dengan Basidiomycota , Pseudoxanthomonas , Sphingomonas , dan Gemmobacter yang tidak terklasifikasi (Gambar 10B ). Hasilnya juga menunjukkan kelimpahan relatif Pseudoxanthomonas , Sphingomonas , dan Lysobacter yang secara signifikan lebih tinggi pada tanaman alfalfa yang terinfeksi P. medicaginis dan diinokulasi AMF dibandingkan pada tanaman yang tidak diinokulasi AMF (Gambar 10C ). Dengan demikian, mikroorganisme ini adalah biomarker mikroba dengan dampak yang lebih besar pada indeks penyakit dibandingkan dengan sifat tanah dan komunitas mikroba (Gambar 10D ) seperti yang diungkapkan oleh pemodelan persamaan struktural (SEM). Secara khusus, biomarker mikroba menunjukkan korelasi negatif langsung dengan indeks penyakit dan juga secara tidak langsung memengaruhinya dengan memengaruhi komunitas mikroba, tanah, dan sifat tanaman (Gambar 10D ).
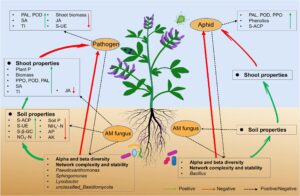
4 DISKUSI
Studi kami adalah salah satu yang pertama mendokumentasikan dampak AMF pada dua penyerang di atas tanah (patogen dan herbivora) dengan memengaruhi rizosfer di bawah tanah. Hubungan kompleks AMF, patogen, dan mesokosmos serangga kurang dipahami, terutama respons mikroba rizosfer bawah tanah terhadap patogen dan kutu daun di atas tanah. Dalam studi ini, kombinasi alfalfa dan tiga organisme lain digunakan untuk menyelidiki apakah respons tanaman AMF terhadap berbagai tekanan biotik dimediasi oleh mikrobioma rizosfer. Hasil kami menunjukkan bahwa AMF secara signifikan memengaruhi kondisi tanah bawah tanah, termasuk AP, AK, dan aktivitas enzim tanah (Tang et al., 2024 ; Wang et al., 2023 ) sementara mikrobioma hifosfernya meningkatkan aktivitas fosfatase tanah, yang mendorong mineralisasi P dan ketersediaan nutrisi tanah (Wang et al., 2023 ). Selain itu, AMF meningkatkan aktivitas enzim pada tunas di atas tanah (PAL, PPO dan POD), serta SA dan TI, sambil menurunkan kadar JA. Perubahan aktivitas enzim tanaman dan hormon di atas tanah secara signifikan terkait dengan sifat tanah. Yang penting, AMF secara signifikan merekrut mikroorganisme yang bermanfaat ke rizosfer alfalfa, meningkatkan ketahanan alfalfa terhadap stres biotik di atas tanah. Lebih jauh lagi, AMF, patogen, dan kutu daun semuanya jelas meningkatkan kompleksitas dan stabilitas jaringan mikroba rizosfer, yang dapat meningkatkan ketahanan tanaman secara keseluruhan. Secara keseluruhan, penemuan-penemuan ini menekankan peran penting jamur AM dalam membentuk interaksi tanaman-patogen-serangga di atas tanah, yang terkait erat dengan mikrobioma rizosfer (Gambar 11 ).
4.1 AMF meningkatkan aktivitas enzim tanah, mendorong penyerapan nutrisi tanaman dan meningkatkan pertahanan terhadap patogen dan kutu daun.
AMF memperluas hifa mereka ke dalam tanah di sekitarnya, mengakses P yang tidak tersedia untuk akar tanaman, menyebabkan penyerapan unsur-unsur mineral tanah yang signifikan, terutama P oleh akar tanaman (Wipf et al., 2019 ), yang dapat mengurangi AP, AK, dan NO 3 − -N tanah. Kandungan P pucuk dan akar yang lebih tinggi, bersama dengan peningkatan biomassa tanaman yang diinokulasi AMF, juga mendukung penurunan AP, AK, dan NO 3 − -N tanah dalam perlakuan tanah AMF. Akuisisi nutrisi yang ditingkatkan ini tidak hanya penting untuk pertumbuhan dan perkembangan tanaman tetapi juga terkait erat dengan aktivitas enzim tanah. Inokulasi AMF meningkatkan aktivitas S-ACP, S-UE, dan S-β-GC, yang terkait erat dengan ketersediaan nutrisi (Qin et al., 2020 ). Meningkatnya kadar P dalam tanah juga berkorelasi positif dengan aktivitas berbagai enzim tanah, yang memainkan peran penting dalam dekomposisi bahan organik dan siklus nutrisi (Zhou et al., 2022 ). Yang terpenting, penelitian telah menunjukkan bahwa aktivasi penyerapan dan asimilasi NH4 + N membantu tanaman padi untuk melawan penyakit hawar pelepah (Chi et al., 2019 ), sementara kadar fosfor berhubungan erat dengan kejadian penyakit blas pada tanaman padi (Campos-Soriano et al., 2020 ). Interaksi antara dinamika nutrisi bawah tanah dan kesehatan tanaman di atas tanah ini menggarisbawahi pentingnya praktik pengelolaan tanah terpadu yang mempertimbangkan interaksi mikroba dan optimalisasi nutrisi.
Dalam studi ini, AMF, patogen, dan kutu daun semuanya mengubah aktivitas enzim di atas tanah dan kadar hormon alfalfa, serta aktivitas enzim tanah bawah tanah, hingga tingkat yang bervariasi (Gambar 3 dan 4 ). Enzim pucuk dan hormon tanaman ini penting untuk berbagai jalur biokimia yang terlibat dalam pertumbuhan, perkembangan, dan mekanisme pertahanan, yang menunjukkan bahwa aktivitas enzim pucuk yang ditingkatkan AMF dan kandungan SA membekali alfalfa dengan mekanisme pertahanan yang lebih baik terhadap P. medicaginis dan kutu daun, yang berkontribusi pada kesehatan ekosistem tanah secara keseluruhan. Analisis korelasi lebih lanjut mengungkapkan hubungan yang signifikan antara aktivitas enzim tanah bawah tanah dan enzim serta hormon pertahanan tanaman di atas tanah (Gambar 6 ). Hasil-hasil ini menunjukkan bahwa perubahan dalam lingkungan tanah dapat memiliki implikasi penting bagi respons fisiologis tanaman di atas tanah (Oldroyd & Leyser, 2020 ), yang berpotensi memainkan peran positif dalam respons tanaman terhadap stres patogen dan serangga hama di atas tanah.
4.2 AMF mengubah mikrobioma rizosfer sebagai respons terhadap tekanan patogen dan kutu daun
AMF mengubah keragaman dan struktur komunitas mikrobioma rizosfer alfalfa yang terpapar infestasi patogen dan kutu daun. AMF secara signifikan merekrut mikroorganisme yang bermanfaat ke rizosfer alfalfa, meningkatkan ketahanan tanaman secara keseluruhan. Penelitian sebelumnya telah menunjukkan bahwa AMF (Xu et al., 2023 ), patogen daun (Berendsen et al., 2018 ; Luo et al., 2022 ) dan serangga pemakan pucuk (Friman, Karssemeijer, et al., 2021 ; Friman, Pineda, et al., 2021 ; Malacrinò et al., 2021 ) dapat mengubah keragaman mikroba rizosfer dan komunitas tanaman, yang mengonfirmasi hasil kami. Penelitian kami selanjutnya menunjukkan bahwa serangan ganda patogen dan serangga hama mengubah keragaman dan struktur komunitas mikrobioma rizosfer. Tumbuhan dapat memodifikasi mikrobioma rizosfernya untuk beradaptasi dengan berbagai pola serangan di atas tanah saat terpapar pada berbagai tekanan (Berendsen et al., 2018 ; Friman, Karssemeijer, et al., 2021 ; Friman, Pineda, et al., 2021 ). Perubahan dalam mikrobioma rizosfer mengindikasikan bahwa serangan tunggal oleh patogen atau kutu daun berbeda dari serangan bersamaan oleh patogen dan kutu daun. Dalam penelitian ini, infeksi gabungan dari patogen dan kutu daun secara signifikan meningkatkan indeks Chao1 bakteri rizosfer pada alfalfa yang tidak diberi perlakuan AMF. Namun, baik patogen tunggal maupun infeksi gabungan dari patogen dan kutu daun pada alfalfa yang diinokulasi AMF tidak memengaruhi keragaman bakteri rizosfer. Terdapat indeks kekayaan, Chao1, ACE, dan Shannon terendah dari bakteri rizosfer pada alfalfa tanpa inokulasi AMF. Hasil-hasil ini menunjukkan bahwa kolonisasi AMF yang lebih awal memainkan peran ekologis yang krusial dalam menjaga keberagaman mikroorganisme di rizosfer, dengan demikian membantu tanaman inang dalam mencegah infeksi patogen di atas tanah dan infestasi kutu daun. Baik AMF maupun kutu daun secara signifikan meningkatkan kekayaan dan ACE bakteri rizosfer, dengan interaksi mereka selanjutnya memperkuat peningkatan ini, menunjukkan efek sinergis. Ini juga dapat disebabkan oleh hubungan simbiosis dalam interaksi tanaman-mikoriza (Zhang et al., 2024 ). AMF merekrut beberapa bakteri, yang dipasoknya dengan karbon yang berasal dari tanaman. Di sisi lain, bakteri menjajah hifa dan mengeluarkan enzim ekstraseluler yang membantu AMF untuk memanfaatkan nutrisi secara efisien (Duan et al., 2024 ). Berbeda dengan bakteri rizosfer, penelitian kami juga mengungkapkan bahwa serangan kutu daun dan patogen, baik sendiri atau dalam kombinasi, dengan atau tanpa AMF, memiliki dampak yang berbeda pada keanekaragaman dan struktur komunitas jamur rizosfer (Wang, Feng, et al., 2021 ; Wang & Kuzyakov, 2024) .). Struktur komunitas jamur dan bakteri berbeda secara signifikan antara kontrol yang tidak diinokulasi AMF dan tanaman yang hanya dihinggapi kutu daun. Namun, pada alfalfa yang tidak diinokulasi AMF, hanya bakteri rizosfer yang menunjukkan respons signifikan terhadap infeksi patogen. Penelitian kami selanjutnya mengungkap bahwa dalam ekosistem sintesis tanaman, mikroorganisme simbiotik AMF, patogen, dan hama serangga, respons jamur dan bakteri rizosfer tanaman terhadap proses di atas dan di bawah tanah berbeda, dengan bakteri tampak lebih rentan terhadap berbagai interaksi (Wang & Kuzyakov, 2024 ).
AMF membentuk hubungan simbiosis dengan akar tanaman dan merekrut mikroorganisme yang bermanfaat, yang mendorong lingkungan mikroba yang lebih mendukung di sekitar akar (Xu et al., 2023 ). Dalam penelitian kami, AMF menginduksi pengayaan jamur Zopfiella , dan bakteri Pseudoxanthomonas , Sphingomonas , dan Lysobacter di tanah rizosfer di bawah infeksi P. medicaginis , yang menunjukkan bahwa AMF membentuk lingkungan mikroba bawah tanah yang bermanfaat, terutama untuk bakteri, setelah stres di atas tanah. Hasil-hasil ini menunjukkan bahwa perakitan dan fungsi mikrobioma tanaman di kompartemen bawah dan di atas tanah mungkin saling berhubungan secara sistematis di bawah infeksi patogen. Beberapa penelitian juga menunjukkan bahwa infeksi patogen di bagian tanaman di atas tanah memicu pembentukan konsorsium bakteri yang bermanfaat di akar dan tanah (Berendsen et al., 2018 ; Luo et al., 2022 ). Misalnya, daun Arabidopsis yang terinfeksi Hyaloperonospora arabidopsidis memperkaya tiga taksa bakteri di rizosfer, yang menginduksi resistensi sistemik terhadap patogen dan meningkatkan pertumbuhan tanaman (Berendsen et al., 2018 ). Selain itu, Pseudoxanthomonas (Deng et al., 2022 ; Xun et al., 2023 ), Sphingomonas (Li et al., 2022 ; Wang, Zhang, Zhang, et al., 2024 ) dan Lysobacter (Li, Xu, et al., 2021 ; Zhao et al., 2021 ) yang memainkan peran penting dalam resistensi patogen, berkorelasi secara signifikan dan negatif dengan indeks dan insidensi penyakit. Yang penting, bakteri ini merupakan biomarker respons positif yang diinduksi AMF terhadap penyakit di atas tanah dalam penelitian ini. Selain ketiga taksa bakteri ini, Bacillus , yang mendorong pertumbuhan, melawan patogen, dan mengurangi kerusakan penyakit pada tanaman inang (Xu et al., 2024 ; Xun et al., 2023 ), paling banyak terdapat di rizosfer alfalfa AM yang dihinggapi kutu daun. Bacillus juga terbukti meningkatkan kemampuan Ageratina adenophora untuk melawan kutu daun ( Aphis gossypiis ) dengan meningkatkan NH4 + -N tanah dan mengakumulasi bakteri bermanfaat lainnya (Yu et al., 2024 ). Meskipun belum ditemukan penelitian tentang dampak Pseudoxanthomonas terhadap kerusakan serangga, Sphingomonas dan Lysobacterjuga dapat menghambat kerusakan serangga pada tanaman inang (Wang, Zhang, Liu, et al., 2024 ; Xun et al., 2023 ; Zhao et al., 2021 ). Oleh karena itu, hasil kami menunjukkan bahwa perakitan mikrobioma rizosfer oleh AMF dikaitkan dengan ketahanan tanaman terhadap patogen di atas tanah dan serangan serangga hama. Komposisi taksa mikroba yang menjajah rizosfer tanaman alfalfa yang terinfeksi P. medicaginis atau yang dihinggapi kutu daun diubah oleh AMF sebagai respons terhadap penyakit dan herbivori serangga. Khususnya, AMF dapat meningkatkan konstruksi dan komposisi komunitas rhizobium (Qiu et al., 2024 ). Hal ini konsisten dengan perubahan pola rhizobia dalam penelitian kami. Kelimpahan relatif rhizobia meningkat secara signifikan di tanah rizosfer tempat AMF berada (Gambar S4 ). Yang penting, AMF dapat memengaruhi komposisi mikrobiota akar melalui modulasi Rhizobiales (Wang, Fang, et al., 2021 ).
Menariknya, tanah rizosfer dapat menjadi sumber utama bakteri di filosfer (Massoni et al., 2021 ; Tkacz et al., 2020 ; Zarraonaindia et al., 2015 ). Mikroorganisme filosfer juga dapat berinteraksi langsung dengan hama dan penyakit daun, dan memengaruhi stres di atas tanah (Li et al., 2022 ; Liu et al., 2023 ; Wang, Zhang, Liu, et al., 2024 ), dan AMF dapat mengubah mikrobioma filosfer (Debray et al., 2022 ). Ini mungkin menjadi alasan penting mengapa AMF mengatur mikroorganisme rizosfer, sehingga secara signifikan memengaruhi stres di atas tanah. Namun, penyelidikan lebih lanjut diperlukan untuk menentukan apakah mikroorganisme yang bermanfaat bermigrasi dari rizosfer alfalfa ke filosfernya. Peningkatan ketahanan terhadap patogen dan kutu daun melalui perekrutan mikroorganisme rizosfer yang bermanfaat oleh AMF berkorelasi dengan perubahan sifat tanah bawah tanah dan sifat pucuk tanaman di atas tanah. Lebih jauh, interaksi AMF dengan patogen dan kutu daun jelas meningkatkan kompleksitas dan stabilitas jaringan mikroba rizosfer, yang selanjutnya meningkatkan ketahanan alfalfa terhadap stres biotik di atas tanah (Qiao et al., 2024 ; Yuan et al., 2021 ). Oleh karena itu, temuan kami menunjukkan bahwa AMF memengaruhi kinerja tanaman, organisme di atas tanah, mikrobioma bawah tanah, dan juga memodulasi interaksi di antara organisme di atas tanah. Penemuan ini menekankan peran penting jamur AM dalam membentuk interaksi tanaman-patogen-serangga di atas tanah, yang terkait erat dengan mikrobioma rizosfer. Hal ini meningkatkan pemahaman mendasar kita tentang hubungan antara tanaman, jamur simbiotik, patogen jamur, dan jaring makanan serangga herbivora, dan dapat memberikan wawasan tentang strategi pengelolaan yang ditujukan untuk mengendalikan hama dan penyakit pertanian secara berkelanjutan melalui manipulasi jamur AM.
4.3 Perubahan mikrobioma rizosfer berhubungan dengan sifat tanah dan profil pertahanan tanaman
Analisis Mantel menunjukkan korelasi signifikan antara aktivitas enzim tanah, nitrogen nitrat, TI, hormon tanaman, biomassa, indeks penyakit, dan mikrobioma rizosfer. Temuan ini menunjukkan bahwa AMF mengubah mikrobiota tanah rizosfer dan terkait erat dengan indikator tanah dan respons pertahanan di atas tanah. AMF mendorong hubungan simbiosis mutualistik yang vital antara tanaman dan mikroorganisme rizosfer dengan meningkatkan penyerapan nutrisi tanaman dan memfasilitasi perolehan karbon fotosintesis mikroba (Duan et al., 2024 ). Pembentukan hubungan simbiosis ini menguntungkan bagi mikroorganisme rizosfer untuk terlibat dalam pertahanan tanaman terhadap berbagai tekanan biotik di atas tanah. Yang penting, SEM menunjukkan bahwa AMF akan merekrut mikroorganisme yang bermanfaat ketika terinfeksi oleh kutu daun dan patogen; mikroorganisme yang bermanfaat memiliki peran positif langsung terhadap infeksi patogen dan juga secara tidak langsung memengaruhinya dengan memengaruhi komunitas mikroba, tanah, dan sifat tanaman. Hasil ini selanjutnya menggarisbawahi peran penting biomarker mikroba rizosfer dalam menentukan indeks penyakit melalui interaksinya dengan komunitas mikroba rizosfer, tanah, dan sifat tanaman. Namun, penelitian di masa mendatang diperlukan untuk menyelidiki efek interaktif biota tanah dan organisme di atas tanah yang hidup berdampingan pada kinerja berbagai anggota serikat untuk menentukan keumuman dampak ini. Eksperimen harus mengeksplorasi efek timbal balik antara berbagai organisme di atas tanah dan komposisi mikroba tanah melalui umpan balik tanaman-tanah (Pineda et al., 2020 ). Wawasan ini akan menjelaskan peran jamur AM dalam membentuk ekologi dan evolusi interaksi tanaman-patogen-serangga.
5 KESIMPULAN
Studi kami menunjukkan bahwa AMF menghubungkan proses dan fungsi tanaman di atas dan di bawah tanah dengan meningkatkan aktivitas enzim tanah bawah tanah yang meningkatkan penyerapan nutrisi dan mendorong perekrutan mikroorganisme rizosfer yang bermanfaat sebagai respons terhadap dua tekanan biotik di atas tanah. AMF juga mengubah respons pertahanan tanaman terhadap infeksi serangga dan patogen di atas tanah, yang menunjukkan bahwa mikrobioma tanah yang diubah oleh jamur AM merupakan pendorong penting dinamika jamur simbiotik berbasis tanaman, patogen jamur, dan jaring makanan serangga herbivora. Selain itu, AMF meningkatkan kompleksitas dan stabilitas mikrobioma rizosfer, yang pada gilirannya berkorelasi dengan peningkatan ketahanan tanaman. Temuan ini menggarisbawahi pentingnya AMF dalam memfasilitasi interaksi tanaman-mikroba yang bermanfaat dan menunjukkan potensi aplikasinya dalam mengelola hama dan penyakit pertanian melalui manipulasi mikroorganisme terkait AMF, dengan implikasi signifikan untuk memahami interaksi spesies dalam sistem alami.


