Abstrak
- Skala makro yang kita gunakan untuk mengukur, memodelkan, dan memprediksi perubahan iklim tidak selaras dengan skala mikro yang digunakan hewan ektotermik kecil untuk merasakan iklim. Untuk memahami pengaruh iklim terhadap keanekaragaman hayati dan potensi dampak ekologis dari perubahan iklim, diperlukan lebih banyak penelitian untuk memahami bagaimana fisiologi hewan ektotermik berhubungan dengan suhu iklim mikro.
- Kanopi pohon merupakan contoh habitat yang menghasilkan iklim mikro ekstrem, dan artropoda di kanopi hutan tropis terancam oleh panas dan pemanasan ekstrem. Namun, situasi di kanopi beriklim sedang kurang jelas. Kearifan konvensional menunjukkan bahwa dinginnya musim dingin membatasi keanekaragaman artropoda arboreal di hutan beriklim sedang, tetapi karena kanopi kurang terlindungi dari suhu ekstrem, panas musim panas juga dapat berperan. Komunitas yang dibatasi oleh panas dan dingin akan merespons perubahan iklim secara berbeda, jadi perbedaan ini sangat penting.
- Dengan menggunakan kerangka hipotesis adaptasi termal dan asimetri relung termal, kami menanyakan apakah semut arboreal beradaptasi secara fisiologis dengan lingkungan ekstrem mereka dan apakah panas musim panas atau dingin musim dingin lebih membuat stres. Kami melacak iklim mikro internal sarang semut di kanopi dan di tanah selama siklus musiman di hutan beriklim sedang di North Carolina, AS. Kemudian, kami mengukur panas (CT maks ) dan toleransi dingin (CT min ) semut pekerja di musim panas dan semi dan membandingkannya dengan iklim mikro yang dialami semut.
- Sarang di tajuk beriklim sedang mengalami suhu ekstrem yang lebih panas dan lebih dingin serta suhu udara yang lebih terpantau dibandingkan sarang semut di tanah. Semut arboreal sebagian mematuhi hipotesis adaptasi termal. Mereka lebih toleran terhadap panas dibandingkan spesies yang bersarang di tanah, tetapi meskipun mengalami suhu yang lebih rendah, mereka kurang toleran terhadap dingin. Semut menyesuaikan toleransi dingin mereka sesuai dengan perubahan musim, tetapi toleransi panas lebih dibatasi secara filogenetik. Panas musim panas tidak mendekati toleransi panas semut di kedua strata, tetapi suhu terendah musim dingin dan musim semi di tajuk melebihi toleransi dingin semut yang bersarang di sana.
- Dengan membandingkan suhu mikroklimat dan fisiologi termal, kami menunjukkan bahwa dinginnya musim dingin–dan bukan panasnya musim panas–kemungkinan membatasi keanekaragaman artropoda di kanopi beriklim sedang.
1. PENDAHULUAN
Suhu lingkungan membentuk pola keanekaragaman hayati dalam skala besar dan kecil (Barreto et al., 2021 ; Coelho et al., 2023 ; González-Tokman et al., 2020 ; Sanders et al., 2007 ). Ketika kita berusaha memahami dampak perubahan iklim dan ancamannya terhadap keanekaragaman hayati, konsekuensi ekologis tetap sulit diprediksi, sebagian karena kurangnya pemahaman mekanistik tentang respons organisme individu terhadap pemanasan (Dawson et al., 2011 ; Urban et al., 2016 ). Misalnya, sebagian besar spesies di planet ini adalah serangga yang panjangnya kurang dari 1 cm (Potter et al., 2013 ; Stork, 2018 ), dan karena ektotermik kecil ini bergantung pada suhu lingkungan untuk mengatur proses fisiologis, mereka mungkin sangat rentan terhadap dampak perubahan iklim (Angilletta, 2009 ; Buckley et al., 2013 ). Sebagian besar serangga mengalami iklim pada skala sentimeter, di mana perubahan tidak dapat diprediksi dan sangat heterogen (Cloudsley-Thompson, 1962 ; Potter et al., 2013 ; Stark et al., 2017 ; Woods et al., 2015 ), tetapi prediksi klimatologi sering kali didasarkan pada suhu udara dan dilaporkan pada skala regional atau global antroposentris (Intergovernmental Panel on Climate Change, 2021 ). Akibatnya, suhu tubuh serangga diprediksi dengan buruk oleh suhu udara skala makro yang disertakan dalam sebagian besar model prediktif (Pincebourde & Woods, 2020 ). Memahami bagaimana fisiologi ektoterm berhubungan dengan suhu iklim mikro yang dialami, dan bagaimana hal itu dapat memengaruhi proses ekologi, dapat memberikan wawasan utama tentang bagaimana iklim memengaruhi keanekaragaman hayati dan potensi dampak ekologis dari perubahan iklim.
Hewan ektotermik di iklim mikro ekstrem sudah mengalami kondisi stres fisiologis, yang membuat mereka rentan terhadap efek negatif pemanasan (Chappell, 1984 ; Kaspari et al., 2015 ; Pincebourde & Casas, 2019 ). Kanopi hutan mewakili salah satu lingkungan ekstrem tersebut, karena iklim mikro arboreal biasanya lebih panas, lebih kering, kurang terisolasi, dan lebih bervariasi daripada yang ada di tanah (De Frenne et al., 2021 ; Hood & Tschinkel, 1990 ; Nadkarni, 1994 ; Scheffers et al., 2013 ). Meskipun demikian, kanopi hutan menampung keanekaragaman ektotermik yang luar biasa, termasuk reptil, amfibi, serangga, dan artropoda lainnya (Erwin, 2013 ; Nadkarni & Longino, 1990 ; Oliveira & Scheffers, 2019 ). Di hutan tropis, artropoda arboreal lebih melimpah dan beragam daripada yang ada di tanah, dan mereka memainkan peran penting dalam fungsi ekosistem (Floren et al., 2002 ; Metcalfe et al., 2014 ; Stork & Blackburn, 1993 ). Organisme yang penting secara ekologis ini sudah beroperasi mendekati batas fisiologis atasnya, dan pemanasan lebih lanjut akibat perubahan iklim diperkirakan akan mengurangi keragaman tajuk tropis (Diamond et al., 2012 ; Kaspari et al., 2015 ; Leahy et al., 2021 ; Nadkarni & Solano, 2002 ; Spicer et al., 2017 ; Woods et al., 2015 ).
Situasi di tajuk beriklim sedang, bagaimanapun, kurang jelas. Berbagai taksa artropoda yang mencakup beberapa tingkat trofik dan kelompok fungsional bergantung pada tajuk hutan beriklim sedang untuk sumber daya utama, setidaknya secara musiman (Sallé et al., 2021 ; Seifert, 2008 ; Ulyshen, 2011 ), tetapi secara keseluruhan, tajuk beriklim sedang menampung lebih sedikit keanekaragaman lokal daripada yang ada di daerah tropis, dan beberapa taksa beriklim sedang menyelesaikan seluruh siklus hidupnya di sana (Floren et al., 2014 ; Floren & Schmidl, 2008 ; Jaffe et al., 2007 ). Kebijaksanaan konvensional menunjukkan bahwa iklim musiman, terutama musim dingin yang dingin, memaksakan filter seleksi, membatasi sebagian besar artropoda beriklim sedang ke tanah yang lebih terisolasi dan membatasi keanekaragaman di tajuk pohon beriklim sedang (Dobzhansky, 1950 ; Wallace, 1878 ). Sementara distribusi serangga di iklim yang lebih dingin diprediksi dengan baik oleh kemampuan mereka untuk mentolerir suhu rendah (Bujan, Roeder, de Beurs, et al., 2020 ; Leahy et al., 2021 ; Sunday et al., 2019 ), hipotesis pembatas musim dingin belum diuji pada gradien vertikal di iklim sedang (tetapi lihat Oliveira & Scheffers, 2019 ). Lebih jauh lagi, studi terbaru menunjukkan bahwa stres panas juga merupakan ancaman bagi komunitas serangga beriklim sedang, terutama di dunia yang memanas (Pincebourde & Casas, 2019 ; Roeder, Bujan, et al., 2021 ; Youngsteadt et al., 2017 , 2023 ). Komunitas yang dibatasi oleh panas dan dibatasi oleh dingin akan merespons perubahan iklim secara berbeda, jadi mengklasifikasikan kendala termal di seluruh strata hutan sangat penting untuk memprediksi dampak pemanasan di masa mendatang. Misalnya, jika musim dingin membatasi keragaman tajuk, maka kita mungkin melihat lebih banyak spesies yang masuk ke tajuk saat suhu musim dingin meningkat, tetapi jika ektoterm tajuk beriklim sedang dibatasi oleh panas, maka, di dunia yang memanas, kita mungkin kehilangan sedikit keragaman tajuk beriklim sedang yang ada. Setiap hasil akan memiliki konsekuensi terhadap fungsi ekosistem hutan beriklim sedang dan struktur komunitas, jadi penting untuk menentukan skenario mana yang lebih mungkin terjadi.
Untuk menentukan apakah keragaman tajuk beriklim sedang dibatasi oleh panas atau dingin musiman, kami memadukan data lingkungan dan fisiologi lintas strata vertikal dan berbagai musim menggunakan semut sebagai sistem studi. Semut (Hymenoptera: Formicidae) adalah sistem yang ideal untuk menilai pembatasan iklim mikro pada spesies tajuk beriklim sedang, karena mereka adalah model mapan untuk fisiologi termal (misalnya Bujan, Roeder, de Beurs, et al., 2020 ; Leahy et al., 2021 ; Roeder, Bujan, et al., 2021 ; Youngsteadt et al., 2023 ) dan membentuk komunitas arboreal dan terestrial yang berbeda, bahkan di hutan beriklim sedang (Floren et al., 2014 ; Seifert, 2008 ). Meskipun fisiologi termal semut darat telah diteliti di wilayah beriklim sedang (misalnya Bujan, Roeder, de Beurs, et al., 2020 ; Roeder, Bujan, et al., 2021 ; Verble-Pearson et al., 2015 ), semut kanopi beriklim sedang, terutama di Amerika Utara, sebagian besar masih belum diteliti. Untuk menentukan bagaimana iklim mikro telah membentuk komunitas semut beriklim sedang, kami mengajukan pertanyaan berikut:
1.1 Apakah semut kanopi sedang beradaptasi secara fisiologis terhadap iklim mikro ekstrem?
Menurut hipotesis adaptasi termal (sensu Kaspari et al., 2015 ), toleransi termal organisme harus berkorelasi dengan suhu minimum dan maksimum lingkungan yang mereka alami, karena secara fisiologis mahal untuk mempertahankan batas toleransi termal yang terlalu luas (Angilletta, 2009 ; Janzen, 1967 ). Dengan kata lain, organisme dalam lingkungan yang bervariasi secara termal akan memiliki rentang toleransi termal yang lebih luas daripada yang ada di lingkungan yang konsisten secara termal. Ektoterm terestrial, termasuk semut, mengikuti hipotesis adaptasi termal dalam skala global (Diamond et al., 2012 ; Diamond & Chick, 2018 ; Kingsolver et al., 2013 ) dan di kanopi tropis (Kaspari et al., 2015 ; Leahy et al., 2021 ). Jika iklim tajuk sedang yang ekstrem membentuk keanekaragaman artropoda, kami perkirakan semut arboreal akan menoleransi suhu rendah yang lebih dingin dan suhu tinggi yang lebih panas daripada spesies yang hidup di tanah, sesuai dengan hipotesis adaptasi termal.
1.2 Kondisi ekstrem manakah—panas atau dingin—yang paling membatasi keanekaragaman tajuk di daerah beriklim sedang?
Suhu terendah musim dingin yang sangat dingin di tajuk pohon yang tidak terisolasi dianggap mengecualikan sebagian besar spesies dari gaya hidup arboreal sedang (Seifert, 2008 ; Ulyshen, 2011 ). Namun, dalam pola yang dikenal sebagai asimetri relung termal (Herrando-Pérez et al., 2020 ), toleransi dingin pada sebagian besar ektotermik, khususnya semut, bersifat labil, sementara toleransi panas lebih dibatasi secara evolusioner (Bujan, Roeder, de Beurs, et al., 2020 ; Hoffmann et al., 2013 ; Leahy et al., 2021 ). Hal ini menunjukkan semut mungkin dapat beraklimatisasi secara musiman terhadap suhu terendah musim dingin yang lebih dingin di tajuk, dan oleh karena itu, mungkin lebih dibatasi oleh suhu tertinggi musim panas yang lebih panas dan lebih rentan terhadap perubahan iklim (Kingsolver et al., 2013 ). Ada bukti bahwa semut dan serangga lain dapat mengubah toleransi termal atas dan bawah mereka untuk melacak perubahan musiman (Bujan, Roeder, Yanoviak, et al., 2020 ; Coulin et al., 2019 ; Maysov & Kipyatkov, 2009 ; Noh et al., 2017 ; Teets & Denlinger, 2013 ), tetapi tidak diketahui apakah aklimatisasi ini cukup untuk melindungi mereka dari ekstrem termal di kanopi. Jika komunitas kanopi beriklim sedang dibatasi oleh panas, suhu tinggi musim panas di kanopi akan melebihi batas fisiologis sebagian besar spesies semut dan mendekati atau melampaui batas bahkan spesies yang beradaptasi dengan kanopi. Jika komunitas kanopi dibatasi oleh dingin, suhu rendah musim dingin akan melebihi toleransi sebagian besar spesies dan paling mendekati batas spesies kanopi.
2 BAHAN DAN METODE
2.1 Lokasi penelitian
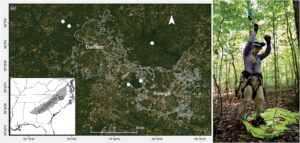
Kami mengambil sampel dari lima lokasi hutan kayu keras sekunder campuran di taman negara bagian atau lahan permainan di ekoregion Piedmont di Carolina Utara bagian tengah (NC), AS (Gambar 1 ). Suhu rata-rata di wilayah ini berkisar dari −2,8°C pada bulan Januari hingga 31,7°C pada bulan Juli (Griffith et al., 2002 ). Di setiap lokasi, kami memilih tiga pohon ek putih ( Quercus alba L.) yang sehat dan dewasa (diameter rata-rata pada tinggi dada: 62,7 cm), spesies yang umum di wilayah tersebut. Semua lokasi dipisahkan oleh setidaknya 3 km, dan pohon yang diambil sampelnya di setiap lokasi berjarak setidaknya 20 m. Selama penelitian, kami mengunjungi setiap pohon empat kali: Juni 2022 untuk memasang pencatat suhu, Juli–Agustus 2022 untuk mengumpulkan pencatat dan semut untuk fisiologi musim panas, Oktober–November 2022 untuk memasang kembali pencatat suhu, dan Maret–April 2023 untuk mengumpulkan pencatat dan semut untuk fisiologi musim semi. Semua kerja lapangan dilakukan dengan izin dari NC State Parks (No. 2021_0490) atau NC Wildlife Resources Commission (No. 22-SC01368). Studi ini tidak memerlukan persetujuan etis untuk perawatan hewan, tetapi kami mengganggu sarang seminimal mungkin, hanya mengumpulkan sejumlah hewan yang diperlukan untuk pengujian fisiologis, dan membekukan semua individu segera setelah pengujian termal. Pernyataan replikasi untuk studi ini dapat ditemukan di Tabel S1 .
2.2 Pengukuran iklim mikro
Pada satu pohon per lokasi, kami mencatat suhu udara sekitar setiap jam di dua lapisan vertikal—kanopi dan tanah—menggunakan pencatat data suhu HOBO® Pendant® (UA-001-08/UA-001-64) berpelindung (Tarara & Hoheisel, 2007 ). Kami menggantung pencatat arboreal di kanopi (~20 m) dengan tali parasut dan memasang pencatat terestrial di tanah tepat di bawahnya (~0,5 m) pada penyangga tanaman logam.
Bahasa Indonesia: Di setiap pohon, kami mencatat suhu sarang internal per jam di setidaknya satu sarang semut per strata (kanopi dan tanah) menggunakan termokopel baja tahan karat fleksibel Tipe K (RS PRO 7877765/7877769; diameter: 1 mm, panjang: 250 mm atau 500 mm) yang terpasang pada pencatat data (Lascar EL-USB-TC). Pencatat data tahan cuaca pada sambungan termokopel dengan Parafilm® dan pita listrik. Kami memasang pencatat ini di sarang semut aktif di kanopi dan di tanah setidaknya 2 minggu sebelum pengambilan sampel semut dengan memasukkan kabel termokopel ke dalam lubang masuk sarang dan memasukkannya sedalam mungkin ke dalam sarang tanpa merusak strukturnya. Meskipun termokopel mungkin tidak berada di lokasi kepadatan semut tertinggi di dalam setiap sarang, termokopel adalah indikator terbaik yang tersedia untuk suhu interior sarang yang utuh. Semut masih aktif di 90% sarang yang ditebang setelah termokopel dilepas. Suhu udara dan sarang dicatat pada tanggal 6 Juni 2022–3 Agustus 2022 dan 30 Oktober 2022–13 April 2023.
2.3 Pengambilan sampel semut
Pada bulan Juni 2022, kami mensurvei tajuk dan tanah di sekitar setiap pohon untuk mencari sarang semut yang aktif. Kami menggunakan teknik panjat tali tunggal (SRT) untuk mengakses tajuk (ketinggian awal rata-rata ± SD: 16,5 ± 2,6 m, kisaran: 13–19 m) (Gambar 1 ). Dalam konteks ini, kami menganggap tajuk pohon mencakup semuanya mulai dari cabang utama pertama hingga daun terluar (Erwin, 2013 ). Dengan menggunakan tali panjat sebagai transek vertikal, kami mencari sarang semut arboreal dengan mengupas kulit kayu, mematahkan cabang yang mati, dan mengikuti pengumpul makanan selama 2 jam per pohon. Secara bersamaan, kami mensurvei sarang terestrial dalam radius 5 m dari pangkal pohon dengan membalik serasah daun, membalik batu, memecahkan batang kayu mati, dan mengikuti pengumpul makanan selama 2 jam. Semua sarang aktif di kedua strata ditandai dengan pita penanda atau bendera pin untuk kunjungan berikutnya.
Kami mengambil sampel semut dari setiap pohon pada satu hari kering di musim panas (11 Juli–3 Agustus 2022) dan musim semi (28 Maret–13 April 2023) antara pukul 8:00 dan 14:00. Kami tidak mengambil sampel semut di musim dingin karena spesies yang dipelajari di sini menjalani diapause selama kondisi dingin (Kipyatkov, 2006 ; Lopatina, 2018 ). Semut di dalam sarang akan sulit ditemukan, dan efek fisiologis dari mengganggu diapause untuk menguji semut mungkin telah mengganggu pengukuran batas termal mereka yang akurat. Kami bekerja di kedua strata secara bersamaan untuk mengunjungi setiap sarang yang ditandai sebelumnya dan sarang yang baru ditemui. Dari setiap sarang aktif, kami mengumpulkan setidaknya 10 pekerja untuk pengujian toleransi termal menggunakan forsep ringan, aspirator, atau InsectaVac (BioQuip 2809B). Semut diangkut dalam kelompok sarang dalam botol plastik berisi bola kapas basah dan disimpan dalam wadah gelap dan terisolasi. Setelah pengukuran toleransi termal, semut dibekukan dan disimpan dalam etanol 70%. Kami mengidentifikasi perwakilan dari setiap sarang menggunakan kunci taksonomi spesifik genus terkini (misalnya DeMarco, 2015 ; Ward & Blaimer, 2022 ) (Tabel S2 ). Spesimen voucher disimpan di Museum Serangga Universitas Negeri NC.
2.4 Uji toleransi termal
Batas termal kritis adalah ukuran umum toleransi termal yang mewakili suhu atas (CT maks ) dan bawah (CT min ) di mana ektotermik kehilangan fungsi lokomotor (Terblanche et al., 2011 ). Dalam waktu 12 jam setelah pengumpulan, kami bertujuan untuk mengukur toleransi panas (CT maks ) untuk lima pekerja per sarang (pekerja rata-rata per sarang ± SD: 4,4 ± 1,0 karena kehilangan setelah pengumpulan) dan toleransi dingin (CT min ) untuk lima pekerja lainnya per sarang (4,5 ± 1,0). Jika memungkinkan, kami menguji beberapa sarang per spesies (musim panas: 3,2 ± 2,5 sarang/spesies; musim semi: 3,2 ± 2,9) (Tabel S2 ). Kami menempatkan semut individu dalam tabung mikrocentrifuge 1,5 mL yang ditutup dengan sepotong spons basah (irisan kosmetik up&up™) untuk menghilangkan perlindungan termal dan mencegah pengeringan. Bahasa Indonesia: Untuk mengukur CT maks , kami menempatkan tabung dalam blok kering pemanasan/pendinginan digital (Grant Instruments PCH-3) yang diatur ke 36°C dan membiarkan semut beraklimatisasi selama 10 menit. Kami menggunakan protokol peningkatan dinamis dengan laju peningkatan rata-rata 0,2°C/menit, yang umum dalam penelitian sebelumnya (misalnya Bujan, Roeder, de Beurs, dkk., 2020 ); setiap 5 menit, kami memeriksa semut untuk kehilangan kontrol otot dan meningkatkan suhu 1°C hingga semua semut mencapai CT maks . Selama pemeriksaan, setiap tabung dikocok perlahan untuk menilai apakah semut telah kehilangan respons perbaikannya; jika ya, suhu tersebut dicatat sebagai CT maks . Kami mengukur CT min dengan cara yang sama dengan menurunkan suhu pada laju yang sama pada blok kering digital pemanasan/pendinginan (VWR 260014-2V) yang diatur ke 20°C. Untuk memantau suhu botol secara langsung di setiap blok, kami menggunakan termokopel Tipe K (RS PRO 7877765/7877769) yang dipasang pada termometer kontak (Fluke 566) dalam tabung yang identik dengan tabung yang berisi semut. Kami menggunakan suhu terukur ini, bukan titik setel blok, dalam analisis. Setiap pengujian (14–19 semut) mencakup beberapa spesies, dan posisi semut dalam satu blok diacak. Selama setiap pengujian, kami menyimpan setidaknya satu individu per spesies dalam tabung yang identik pada suhu ruangan di rak tabung mikrocentrifuge buram sebagai kontrol. Tidak ada kontrol yang kehilangan respons perbaikan atau mati selama pengujian.
2.5 Analisis data
Analisis data dilakukan di R 4.3.2 dan RStudio 2023.12.1.402 (Posit Team, 2024 ; R Core Team, 2023 ). Kecuali jika dinyatakan lain, kami mengevaluasi kecocokan setiap model campuran linier dengan plot diagnostik dan pengujian di DHARMa 0.4.6 (Hartig & Lohse, 2022 ), menguji signifikansi prediktor menggunakan uji Wald χ 2 Tipe II di mobil 3.1-2 (Fox & Weisberg, 2019 ), dan menghitung estimasi rata-rata marginal untuk setiap strata dengan emmean 1.8.9 (Lenth et al., 2023 ).
2.5.1 Suhu iklim mikro
Untuk membandingkan kondisi iklim mikro antar strata, kami membatasi analisis kami pada tanggal saat semua sensor suhu dipasang dan direkam; di musim panas, ini sesuai dengan 16 hari sebelum uji toleransi termal pertama (25 Juni 2022–10 Juli 2022). Untuk memastikan bahwa hasil kami sebanding di seluruh musim, kami juga membatasi analisis iklim mikro musim dingin dan musim semi pada periode 16 hari. Untuk mewakili musim dingin, kami menganalisis 16 hari terdingin berturut-turut (14 Desember 2022–29 Desember 2022), dan di musim semi, kami menganalisis 16 hari sebelum uji toleransi termal (12 Maret 2023–27 Maret 2023). Untuk setiap periode waktu tersebut, kami menghitung suhu udara minimum, rata-rata, dan maksimum harian di tajuk dan di tanah di setiap lokasi. Kami membangun model campuran linear terpisah dengan glmmTMB 1.1.8 (Brooks et al., 2017 ) untuk setiap metrik suhu (minimum, rata-rata, maksimum) dan musim (musim panas, musim dingin, musim semi). Kami memodelkan metrik suhu sebagai fungsi strata (arboreal/terestrial) dengan efek acak lokasi, sensor, dan tanggal. Untuk memperhitungkan autokorelasi temporal, kami menyertakan struktur varians autoregresif orde pertama (AR1) yang dikelompokkan berdasarkan sensor (Zuur et al., 2009 ). Kami membandingkan suhu sarang antara strata dengan cara yang sama seperti suhu udara, setelah menghitung suhu harian minimum, rata-rata, dan maksimum setiap sarang dan menyertakan efek acak tambahan pohon di setiap model.
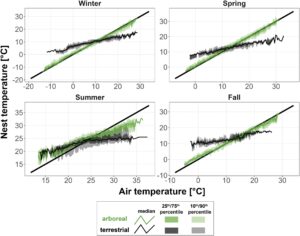
Untuk memvisualisasikan seberapa baik suhu sarang melacak suhu udara, kami membuat plot kuantil bersyarat untuk seluruh durasi pengukuran iklim mikro sarang (7 Juni 2022–2 Agustus 2022 dan 31 Oktober 2022–11 April 2023). Sementara plot kuantil-kuantil tradisional (plot QQ) mempertimbangkan distribusi pengamatan dan prediksi secara terpisah, kuantil bersyarat dihitung menggunakan pengamatan yang secara langsung sesuai dengan interval prediksi tertentu (Carslaw & Ropkins, 2012 ). Kami menggunakan aturan Freedman–Diaconis untuk menentukan jumlah interval yang tepat untuk kumpulan data kami (Freedman & Diaconis, 1981 ). Kemudian, kami menghitung median, persentil ke-10, ke-25, ke-75 dan ke-90 dari suhu sarang yang dikondisikan pada suhu udara yang tercatat selama interval yang sama untuk setiap strata (arboreal/terestrial) dan musim (musim dingin/semi/panas/gugur) menggunakan fungsi ‘conditionalQuantile’ dari openair 2.18-0 (Carslaw & Ropkins, 2012 ). Karena pencatat termokopel sarang kami tidak aktif sepanjang tahun, setiap musim sesuai dengan tanggal-tanggal berikut: musim panas: 7 Juni 2022–2 Agustus 2022; musim gugur: 31 Oktober 2022–30 November 2022; musim dingin: 1 Desember 2022–28 Februari 2023; musim semi: 1 Maret 2023–11 April 2023. Kami memplot kuantil bersyarat dengan ggplot2 3.4.4 (Wickham, 2016 ).
2.5.2 Toleransi termal di seluruh strata
Kami menghitung rata-rata maksimum termal kritis (CT maks ) dan minimum (CT min ) untuk setiap sarang semut di musim panas dan semi. Untuk sarang tempat kami mengumpulkan beberapa spesies ( n = 5), toleransi termal rata-rata setiap spesies dihitung secara terpisah. Semua spesies yang disertakan dalam analisis eksklusif untuk satu strata. Untuk membandingkan toleransi termal di kanopi dan di tanah, kami membuat model campuran linier terpisah untuk CT maks dan CT min di setiap musim (musim panas, semi) menggunakan lme4 1.1-35.1 (Bates et al., 2015 ). Kami memodelkan setiap sifat termal sebagai fungsi strata dengan efek acak dari lokasi, pohon, dan spesies. Karena kami mengajukan pertanyaan kami di tingkat komunitas dan tidak tertarik pada perbedaan interspesifik, beberapa sarang dari spesies yang sama tidak memberikan informasi independen pada analisis kami tentang perbedaan antara strata; dengan demikian, kami memperlakukan spesies sebagai efek acak.
Bahasa Indonesia: Untuk menguji sinyal filogenetik dalam toleransi termal, kami menggunakan filogeni semut global yang dikalibrasi waktu yang mencakup beberapa spesies representatif dari setiap genus (Moreau & Bell, 2013 ). Kami memangkas pohon tersebut menjadi 10 genera yang disertakan dalam studi ini menggunakan ‘keep.tip’ ape 5.7.1 (Paradis & Schliep, 2019 ) dan menambahkan Nylanderia sebagai saudara bagi Paratrechina (Kallal & LaPolla, 2012 ) dengan ‘AddTip’ dari TreeTools 1.10.0 (Smith, 2019 ). Dalam kasus di mana spesies representatif tidak cocok dengan spesies yang kami pelajari, kami menambahkan spesies baru ke pohon tersebut sebagai saudara bagi spesies representatif untuk genus tersebut menggunakan ‘AddTip’ dari TreeTools . Kami membuat politomi lunak pada setiap simpul genus dengan memberikan ujung baru tersebut panjang cabang yang sama dengan spesies saudara. Kami kemudian menyingkirkan semua spesies yang tidak termasuk dalam studi kami dengan ‘drop.tip’ kera (Gambar S1 ). Kami menggunakan model campuran linear dalam lme4 untuk memperkirakan CT maks dan CT min setiap spesies di musim panas dan semi dengan memodelkan setiap sifat termal di setiap musim dengan efek tetap spesies dan efek acak lokasi dan pohon. Menggunakan ‘phylosig’ dalam phytools 2.1.1 (Revell, 2024 ), kami menghitung λ Pagel (Pagel, 1999 ) dan K Blomberg (Blomberg et al., 2003 ) untuk menguji sinyal filogenetik di setiap sifat termal di setiap musim.
2.5.3 Plastisitas musiman
Untuk memahami aklimatisasi musiman dalam toleransi termal, kami membatasi kumpulan data kami ke 14 spesies (4 arboreal, 10 terestrial) yang sifat termalnya kami ukur baik di musim panas maupun musim semi (Tabel S1 dan S2 ). Kemudian, kami memodelkan CT maks dan CT min sebagai fungsi strata, musim, dan interaksinya, dengan efek acak dari lokasi, pohon, dan spesies, menggunakan model campuran linier dalam lme4 . Kami menghitung estimasi mean marginal dan merencanakan kontras efek utama sederhana untuk interaksi antara strata dan musim dengan emmean , dengan memperhitungkan beberapa perbandingan dengan probabilitas t multivariat (Lenth et al., 2023 ).
3 HASIL
3.1 Iklim Mikro
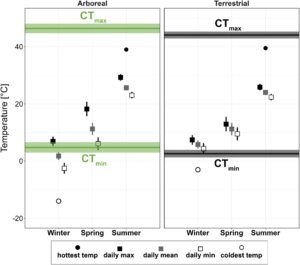
Suhu udara menunjukkan beberapa bukti lingkungan arboreal yang lebih ekstrem. Pada musim panas, suhu udara sekitar rata-rata 0,5°C lebih hangat di tajuk daripada di tanah, dan suhu tertinggi harian 1°C lebih hangat, sementara suhu terendah harian tidak berbeda antara strata (Tabel 1 ). Pada musim dingin dan semi, baik suhu udara rata-rata, suhu tertinggi harian, maupun suhu terendah harian tidak berbeda antara tajuk dan tanah (Tabel 1 ; Gambar S2 ).
| Musim | Tanggapan | Udara | Sarang | ||||||
|---|---|---|---|---|---|---|---|---|---|
| Perkiraan (°C), arboreal–terestrial | χ 2 | df | p (> χ2 ) adalah | Perkiraan (°C), arboreal–terestrial | χ 2 | df | p (>χ2 ) adalah | ||
| Musim dingin | Maksimal | 0,01 | 0,0004 | 1 | 0,985 | -0,47 | 0,518 | 1 | 0.472 |
| Berarti | -0,54 | 0.897 | 1 | 0.344 | -4,06 | 94.46 | 1 | <0,001 | |
| menit | -1,81 | 2.048 | 1 | 0.153 | -6,87 | 195.80 | 1 | <0,001 | |
| Musim semi | Maksimal | -0,21 | 0,035 | 1 | 0.851 | 5.27 | 66.57 | 1 | <0,001 |
| Berarti | 0,01 | 0,0005 | 1 | 0,981 | 0,06 | 0,011 | 1 | 0,915 | |
| menit | -0,60 | 0.846 | 1 | 0.358 | -3,44 | 55.56 | 1 | <0,001 | |
| Musim panas | Maksimal | 1.01 | Tanggal 26.21 | 1 | <0,001 | 3.39 | Tanggal 38.09 | 1 | <0,001 |
| Berarti | 0.51 | Tanggal 20.19 | 1 | <0,001 | 1.63 | 16.29 | 1 | <0,001 | |
| menit | -0,01 | 0,003 | 1 | 0,951 | 0.69 | 1.145 | 1 | 0,285 | |
Catatan : Nilai estimasi adalah kontras yang diestimasikan model antara habitat arboreal dan terestrial. Setiap kombinasi musim (musim dingin, musim semi, musim panas), respons suhu (suhu maksimum harian, rata-rata, minimum), dan iklim mikro (udara, sarang) diperlakukan dalam model terpisah. Nilai yang dicetak tebal menunjukkan signifikansi statistik pada level 0,05.
Perbedaan mikroklimat antara strata vertikal lebih jelas dalam kumpulan data sarang, di mana lingkungan tajuk jauh lebih ekstrem (Tabel 1 ; Gambar 2 ). Setelah membuang sensor yang terganggu dan tidak berfungsi, kami memperoleh suhu di 10 sarang arboreal dan terestrial di musim panas dan 10 sarang arboreal dan sembilan sarang terestrial di musim dingin dan semi (Tabel S1 dan S3 ). Termokopel arboreal mencatat suhu di sarang 8–19 m di atas tanah (rata-rata ± SD: 14,3 ± 3,2 m). Seperti yang diharapkan, tajuk lebih panas di musim panas: suhu rata-rata sarang arboreal 1,6°C lebih hangat dan suhu tertinggi harian 3,4°C lebih hangat daripada di tanah, tetapi suhu terendah harian tidak berbeda secara signifikan antara strata. Kami mengamati pola yang berlawanan di musim dingin: suhu rata-rata sarang arboreal 4°C lebih dingin, dan suhu sarang rendah harian 6,7°C lebih dingin daripada di tanah, sementara suhu tinggi tidak berbeda antara strata. Di musim semi, suhu tinggi dan rendah kanopi sama-sama ekstrem: Sarang semut arboreal mengalami suhu tinggi harian 5,3°C lebih hangat dan suhu rendah harian 3,4°C lebih dingin. Variabilitas suhu tinggi di kanopi ini menyebabkan rata-rata harian yang sama di kanopi dan di tanah, meskipun ada perbedaan yang jelas dalam ekstrem. Di semua musim, suhu sarang di kanopi melacak suhu udara dengan cermat, sementara sarang di tanah lebih terisolasi dari fluktuasi suhu (Gambar 3 ).
3.2 Toleransi termal di seluruh strata
Kami mengukur toleransi termal atas dan bawah dari 18 spesies di musim panas (6 arboreal, 12 terestrial) dan 17 spesies di musim semi (4 arboreal, 13 terestrial) (Tabel S1 dan S2 ; Gambar S1 ). Satu spesies tambahan ditemukan bersarang di kedua strata: semut jarum invasif, Brachyponera chinensis (Emery). Karena ini adalah satu-satunya spesies non-asli yang kami kumpulkan dan satu-satunya spesies yang tidak dapat secara definitif ditugaskan ke suatu strata, kami mengecualikannya dari semua analisis.
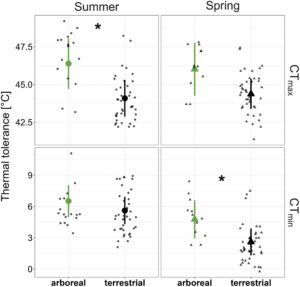
Sejalan dengan hipotesis adaptasi termal, semut di kanopi memiliki toleransi panas yang lebih tinggi daripada semut di tanah. Rata-rata, CT maksimum musim panas 2,3°C lebih tinggi pada spesies arboreal ( χ2 = 5,86 , df = 1, p = 0,015), sedangkan CT minimum musim panas tidak berbeda antara strata ( χ2 = 1,27, df = 1, p = 0,260) (Gambar 4 ). Namun, bertentangan dengan harapan kami, pada musim semi, semut arboreal tidak menoleransi suhu ekstrem yang lebih dingin daripada spesies yang bersarang di tanah. Sebaliknya, hasil kami menunjukkan pola yang berlawanan, dan rata-rata CT minimum musim semi adalah 2,2°C lebih rendah di tanah ( χ 2 = 5,74, df = 1, p = 0,017), sedangkan CT maksimum musim semi tidak berbeda secara signifikan antar strata ( χ 2 = 3,03, df = 1, p = 0,082) (Gambar 4 ). Kami mendeteksi sinyal filogenetik yang kuat dalam CT max di kedua musim (musim panas: Pagel’s λ = 1, p = 0,004, Blomberg’s K = 1,12, p = 0,002; musim semi: Pagel’s λ = 1, p = 0,005, Blomberg’s K = 0,947, p = 0,002), tetapi tidak ada bukti sinyal filogenetik untuk CT min (musim panas: Pagel’s λ = 0,553, p = 0,334, Blomberg’s K = 0,962, p = 0,099; musim semi: Pagel’s λ = 0,498, p = 0,225, Blomberg’s K = 0,722, p = 0,079).
3.3 Plastisitas musiman
Semut tidak menggeser toleransi panas mereka antara musim panas dan musim semi di kedua strata, mendukung komponen filogenetik yang kuat pada CT maks (Tabel 2 ). Namun, untuk mendukung asimetri relung termal, semut di tanah beraklimatisasi dengan kondisi yang lebih dingin, menurunkan CT min mereka sebesar 2,9 °C dari musim panas ke musim semi (Tabel 2 ; Gambar S3 ). Salah satu spesies semut arboreal, Crematogaster vermiculata Emery, tampaknya sedikit meningkatkan CT min -nya di musim dingin, berlawanan dengan semua spesies lain yang kami ukur (Tabel S2 , Gambar S1 ). Ketika spesies ini dihilangkan dari analisis, semut di kanopi beraklimatisasi mirip dengan rekan-rekan terestrial mereka, menurunkan CT min mereka sebesar 2,1 °C dari musim panas ke musim semi.
| Tanggapan | Lapisan | Musim | Kontras | Perkiraan (°C) | Bahasa Inggris | df | rasio t | nilai p |
|---|---|---|---|---|---|---|---|---|
| CT maks | Pohon | — | Musim panas/musim semi | 0.409 | 0.31 | 83.6 | 1.32 | 0.344 |
| Terestrial | — | Musim panas/musim semi | -0,365 | 0.19 | 86 | -1,93 | 0.11 | |
| — | Musim panas | Pohon/terestrial | 2.54 | 0,99 | 12.5 | 2.54 | 0,034 tahun | |
| — | Musim semi | Pohon/terestrial | 1.77 | 1 | 12.7 | 1.76 | 0.132 | |
| CT menit | Pohon | — | Musim panas/musim semi | 0.83 | 0.618 | 82.2 | 1.34 | 0.332 |
| Terestrial | — | Musim panas/musim semi | 2.86 | 0,371 tahun | 85.6 | 7.73 | <0,001 | |
| — | Musim panas | Pohon/terestrial | 0.323 | 1.11 | 13.6 | 0.291 | 0,918 tahun | |
| — | Musim semi | Pohon/terestrial | 2.36 | 1.12 | 14.3 | 2.1 | 0,083 tahun | |
| CT menit
tidak termasuk Crematogaster vermiculata |
Pohon | — | Musim panas/musim semi | 2.11 | 0,735 | 74.8 | 2.88 | 0,010 |
| Terestrial | — | Musim panas/musim semi | 2.88 | 0,364 tahun | 77.1 | 7.9 | <0,001 | |
| — | Musim panas | Pohon/terestrial | 1.13 | 1.3 | 12.7 | 0,87 | 0,538 | |
| — | Musim semi | Pohon/terestrial | 1.89 | 1.3 | 12.7 | 1.45 | 0.246 |
Catatan : Nilai yang dicetak tebal menunjukkan signifikansi statistik pada tingkat 0,05.
3.4 Membandingkan iklim mikro sarang dengan toleransi termal
Suhu musim panas tidak mendekati toleransi termal atas semut di kedua strata. Bahkan suhu terpanas yang kami ukur di sarang kanopi (39,0 °C) masih lebih dari 5 °C di bawah perkiraan rata-rata marginal untuk komunitas terestrial dan lebih dari 7 °C di bawah spesies arboreal (Gambar 2 ). Di sisi lain, suhu sarang musim dingin di kedua strata, terutama suhu terendah musim dingin, mendekati atau melebihi toleransi dingin semut. Sementara hanya suhu musim dingin terdingin yang mengancam semut di tanah, spesies arboreal mengalami suhu 3 °C di bawah CT min mereka pada hari musim dingin rata-rata (Gambar 2 ). Suhu sarang terdingin yang kami catat di kanopi (−14,0 °C) lebih dari 12 °C di bawah toleransi dingin terendah dari semua semut yang kami ukur. Di musim semi, suhu tertinggi dan rata-rata harian berada dalam batas termal semut di kedua strata; tetapi selama jam-jam terdingin di hari musim semi, sarang di tajuk mendekati toleransi dingin semut arboreal, sementara semut terestrial mengalami suhu 7°C di atas batas termal bawah mereka (Gambar 2 ). Pada lebih dari setengah hari antara 31 Oktober 2022 dan 12 April 2023, kami mencatat suhu sarang tajuk di bawah perkiraan rata-rata marginal dari CT min komunitas semut arboreal .
4 DISKUSI
Kami membandingkan suhu mikroklimat skala halus dan toleransi termal semut selama siklus musiman untuk memahami kendala iklim pada keanekaragaman serangga arboreal di hutan beriklim sedang. Kami menyajikan bukti untuk mendukung lebih dari 150 tahun kearifan konvensional yang sebelumnya tidak teruji dan hipotesis bahwa dinginnya musim dingin—bukan panasnya musim panas—kemungkinan membatasi keanekaragaman tajuk di ekosistem hutan beriklim sedang. Selain itu, kami menemukan dukungan parsial untuk dua pola biologi termal di komunitas hutan beriklim sedang kami. Toleransi panas semut mengikuti hipotesis adaptasi termal, tetapi toleransi dingin semut tidak. Konsisten dengan asimetri relung termal, semut menyesuaikan toleransi dingin mereka, tetapi tidak toleransi panas, agar sesuai dengan perubahan musim.
Bahasa Indonesia: Sesuai dengan penelitian sebelumnya dalam sistem tropis dan sedang, tajuk di area penelitian kami memiliki lingkungan termal yang lebih ekstrem daripada lantai hutan (De Frenne et al., 2021 ; Friedland et al., 2003 ; Scheffers et al., 2013 ). Namun, hasil kami menunjukkan bahwa hanya mengandalkan suhu udara, bahkan pada skala meso (sensu Pincebourde & Woods, 2020 ), meremehkan intensitas divergensi iklim mikro antara strata vertikal, terutama pada cuaca yang lebih dingin (Tabel 1 ; Gambar 3 dan S2 ). Di musim dingin dan semi, suhu udara harian konsisten antara strata vertikal; sementara itu, sarang semut di tajuk mengalami variasi suhu harian 6–9°C lebih banyak daripada yang ada di tanah (Gambar 2 ). Mikrohabitat sering kali dapat menyangga organisme dari suhu lingkungan yang penuh tekanan (Scheffers, Edwards, et al., 2014 ; Scheffers, Evans, et al., 2014 )—yaitu, suhu lingkungan yang menyebabkan kerusakan fisiologis dengan mencapai atau mendekati batas termal kritis. Sarang semut terestrial—terisolasi oleh naungan tajuk, serasah daun, dan tanah—adalah contoh dari fenomena ini. Sebaliknya, sarang semut arboreal di kayu mati dan celah-celah kulit kayu tidak memiliki penyangga yang baik dan melacak dengan cermat variasi termal yang kami deteksi dalam suhu udara (Gambar 3 ). Lingkungan termal yang sangat bervariasi dapat memengaruhi perkembangan, rentang hidup, kesuburan, dan morfologi serangga dengan efek berjenjang pada dinamika populasi, fenologi, dan interaksi spesies (Colinet et al., 2015 ), dan variabilitas iklim diperkirakan akan meningkat seiring dengan perubahan iklim di masa mendatang (Panel Antarpemerintah tentang Perubahan Iklim, 2021 ). Hasil kami menekankan pentingnya menggunakan pengukuran lingkungan yang relevan secara ekologis saat menguji hipotesis termal, yang dapat dicapai dengan mencocokkan skala dan substrat pengukuran dengan organisme yang diinginkan.
Kami menemukan bukti parsial bahwa semut di kanopi mematuhi hipotesis adaptasi termal dan beradaptasi secara fisiologis dengan lingkungan ekstrem mereka. Di musim panas, toleransi panas semut sedang lebih tinggi di kanopi panas daripada di tanah yang lebih dingin (Gambar 4 ), menggemakan temuan dari kanopi tropis (Kaspari et al., 2015 ; Leahy et al., 2021 ). Di kanopi tropis, suhu udara, kulit kayu, dan permukaan daun secara teratur melebihi batas termal atas semut (Kaspari et al., 2015 ; Leahy et al., 2021 ; Spicer et al., 2017 ). Namun, dalam sistem hutan sedang kami, kami menemukan bahwa bahkan suhu udara, permukaan, atau sarang terpanas pun tidak mendekati toleransi panas semut (Gambar 2 ; Gambar S1 dan S2 ) (suhu permukaan batang: data tidak dipublikasikan). Hal ini menunjukkan bahwa panas musim panas tidak membatasi keanekaragaman pohon di tajuk beriklim sedang, dan semut pohon beriklim sedang memiliki risiko lebih rendah terhadap stres panas sebagai dampak langsung dari pemanasan global di masa mendatang dibandingkan semut di daerah tropis. Meskipun demikian, peningkatan kekeringan, frekuensi kejadian panas ekstrem, dan matinya tajuk akibat perubahan iklim masih dapat mengubah iklim mikro tajuk melebihi kemampuan toleransi beberapa spesies (De Frenne et al., 2021 ; Intergovernmental Panel on Climate Change, 2021 ; Sallé et al., 2021 ).
Semut arboreal tidak beradaptasi secara fisiologis dengan musim dingin yang ekstrem atau lingkungan kanopi musim semi yang mudah berubah. Sebaliknya, semut terestrial menoleransi suhu yang lebih rendah daripada semut arboreal, meskipun sarang terestrial tidak sedingin yang ada di kanopi (Gambar 2 dan 4 ). Ada kemungkinan bahwa karena kami mengukur fisiologi termal semut di musim semi, suhu musim dingin terdingin tidak secara langsung relevan dengan metrik toleransi dingin kami. Namun, para pekerja yang kami uji di musim semi akan terpapar pada kondisi musim dingin karena mereka melewati musim dingin sebagai orang dewasa di sarang (Kipyatkov, 2006 ; Lopatina, 2018 ). Lebih jauh lagi, bahkan di musim semi, semut kanopi di dalam sarang mengalami minimum harian yang sama dengan CT min mereka , sementara semut di tanah mengalami minimum sekitar 7°C lebih hangat daripada CT min mereka . Jadi, pertanyaannya tetap: Mengapa toleransi dingin semut menyimpang dari apa yang diprediksi di bawah hipotesis adaptasi termal?
Satu penjelasan untuk pola yang diamati dalam toleransi dingin bisa jadi karena plastisitas musiman. Toleransi dingin dan plastisitas dalam toleransi dingin melacak suhu lingkungan lebih dekat daripada toleransi panas dan plastisitasnya, yang lebih dibatasi secara evolusioner (Araújo et al., 2013 ; Bujan, Roeder, de Beurs, et al., 2020 ; Gunderson & Stillman, 2015 ; Hoffmann et al., 2013 ; Sunday et al., 2019 ). Pola ini, yang dikenal sebagai asimetri relung termal, tampak dalam hasil plastisitas musiman kami, di mana semut menurunkan toleransi dingin mereka sebagai respons terhadap suhu musim semi yang dingin, tetapi tidak meningkatkan toleransi panas mereka dalam suhu panas musim panas (Tabel 2 ; Gambar S3 ). Mengingat sifat toleransi dingin yang fleksibel dan bahwa kami hanya mengukur fisiologi musim semi dan musim panas, ada kemungkinan kami melewatkan periode waktu di musim dingin di mana semut di kanopi paling toleran terhadap dingin, yang cocok dengan lingkungan dingin yang ekstrem. Akan tetapi, suhu terendah di kanopi pohon pada musim semi masih lebih ekstrem dibandingkan di tanah, dan meskipun begitu, semut arboreal tetap kurang tahan terhadap dingin dibandingkan semut terestrial (Gambar 2 ).
Terlepas dari apakah toleransi dingin semut beriklim sedang tidak mengikuti hipotesis adaptasi termal, atau kami gagal mendeteksinya dengan desain pengambilan sampel kami, tampaknya dinginnya musim dingin mungkin merupakan faktor pembatas bagi keragaman tajuk beriklim sedang. Titik terendah di tajuk lebih ekstrem daripada titik terendah di tanah (Gambar 2 ). Bahkan suhu terdingin dari sarang terestrial (−1,5°C) sama dengan menit CT terendah yang kami ukur, dalam kisaran batas termal semut yang dipublikasikan, dan secara umum dapat bertahan hidup bagi sebagian besar spesies semut beriklim sedang (Roeder, Roeder, et al., 2021 ; Willot et al., 2023 ). Namun, di kanopi, suhu sarang terdingin (−14°C) mendekati titik superdingin yang mematikan bagi semut beriklim sedang dan boreal (−40 hingga −8°C) (Berman et al., 2017 ; Cannon & Fell, 1992 ; Martin-Roy et al., 2021 ; Roeder, Roeder, et al., 2021 ), yang menunjukkan bahwa bahkan dengan CT min musim dingin yang lebih rendah , semut arboreal akan mengalami stres termal yang parah. Kami mengakui bahwa CT min mungkin bukan pengukuran toleransi dingin yang paling informatif bagi spesies yang mampu melakukan diapause musim dingin yang sebenarnya, seperti yang dipelajari di sini. CT min mengukur timbulnya koma dingin karena hilangnya fungsi otot, dan sementara mortalitas individu lebih tinggi pada suhu pada atau di bawah CT min , sebagian besar individu dapat pulih dari periode koma dingin (Andersen et al., 2015 ; Kipyatkov, 2001 ; Overgaard & MacMillan, 2017 ; Roeder, Roeder, et al., 2021 ). Hilangnya koordinasi otot selama koma dingin mungkin tidak penting bagi semut dalam diapause, karena mereka menghentikan perkembangan, reproduksi, dan pada beberapa spesies, aktivitas sama sekali (Kipyatkov, 2001 ). Dengan demikian, metrik toleransi dingin lainnya yang lebih akurat mewakili mortalitas, seperti suhu mematikan yang lebih rendah, waktu mematikan pada suhu rendah atau titik superdingin, mungkin lebih menunjukkan toleransi dingin tingkat koloni. Meskipun demikian, paparan kronis terhadap suhu dingin yang tidak mematikan dapat mengakumulasi cedera dingin seluler yang akhirnya mengakibatkan kematian (Overgaard & MacMillan, 2017 ), dan penelitian di masa depan harus berupaya untuk mengukur secara pasti tekanan termal semut di kanopi.
Kami tidak menemukan plastisitas musiman dalam toleransi panas di tingkat komunitas. Mengingat komponen filogenetik yang kuat pada CT max dan kurangnya suhu panas yang berbahaya, tidak mengherankan bahwa toleransi panas tetap konsisten sepanjang tahun. Hasil kami kontras dengan hasil Bujan, Roeder, Yanoviak, et al. ( 2020 ), di mana CT max lebih rendah di musim yang lebih dingin. Ada kemungkinan perbedaan ini disebabkan oleh habitat pengambilan sampel yang berbeda. Sementara komunitas semut kami berasal dari hutan beriklim sedang, komunitas mereka berasal dari habitat padang rumput beriklim sedang yang lebih terbuka, di mana suhu biasanya lebih hangat daripada di hutan dan radiasi matahari musim panas dapat memanaskan permukaan secara berlebihan (Villegas et al., 2010 ). Hasil yang kontras ini menunjukkan bahwa beberapa spesies lebih plastis secara musiman daripada yang lain, dan penelitian di masa depan diperlukan untuk memahami bagaimana spesies tertentu dapat menyesuaikan toleransi panas mereka, sementara yang lain tidak.
5 KESIMPULAN
Kondisi musim dingin memainkan peran penting dalam struktur, produktivitas, dan fenologi ekosistem beriklim sedang (Kreyling, 2020 ; Ladwig et al., 2016 ; Williams et al., 2015 ), dan hasil kami menunjukkan bahwa kondisi tersebut juga dapat membatasi keanekaragaman tajuk beriklim sedang. Musim dingin rata-rata yang lebih hangat akibat perubahan iklim dapat mengurangi sebagian tekanan dingin yang kami amati di tajuk. Selain itu, berkurangnya lapisan salju dan meningkatnya embun beku di tanah akibat variabilitas suhu dapat meningkatkan tekanan dingin bagi spesies yang hidup di tanah. Oleh karena itu, di dunia yang semakin hangat, kita mungkin melihat lebih banyak spesies yang menjelajah ke dalam panas tajuk beriklim sedang.