
ABSTRAK
Centrostephanus longispinus (Echinodermata: Echinoidea) merupakan spesies bulu babi endemik dan dilindungi di Laut Mediterania. Bulu babi berduri panjang lainnya, Diadema setosum dari Laut Merah, memasuki Laut Mediterania pada tahun 2006 dan menyebar dengan cepat ke seluruh cekungan timur. Sebagai hewan pemakan rumput kompetitif yang penting, D. setosum berpotensi menyebabkan perubahan signifikan pada kumpulan komunitas bentik asli. Tujuan utama dari penelitian ini adalah (i) untuk memberikan sintesis ciri morfologi dan ekologi utama Diadematidae, (ii) untuk menentukan pola distribusi spasiotemporal spesies bulu babi endemik dan invasif, (iii) untuk mengidentifikasi area kritis tempat upaya konservasi untuk C. longispinus dan strategi pengelolaan untuk D. setosum harus diprioritaskan dan (iv) untuk menerapkan protokol sederhana untuk melatih peneliti dan ilmuwan warga dalam identifikasi spesies. Kumpulan data terbesar tentang distribusi spasial dan temporal dari dua spesies pada skala Mediterania diperoleh dari berbagai sumber, sementara Model Aditif Umum diterapkan untuk memperkirakan probabilitas keberadaan spesies di seluruh cekungan. Mediterania barat laut muncul sebagai hotspot untuk C. longispinus , menyorotinya sebagai area utama untuk melestarikan spesies endemik ini dan mempelajari dinamika populasinya. Sebaliknya, pengumpulan data perlu diintensifkan di wilayah Adriatik timur, Levantina, dan Mediterania timur-tengah, di mana informasi saat ini terbatas atau kurang. Laut Aegea dan Ionia muncul sebagai wilayah prioritas untuk menilai risiko invasi yang ditimbulkan oleh D. setosum , di mana penerapan protokol identifikasi yang diusulkan akan sangat penting untuk mencegah kesalahan identifikasi dengan spesies asli.
1 Pendahuluan
Famili Diadematidae (Echinodermata: Echinoidea) saat ini mencakup tujuh genera dan 31 spesies yang diketahui, tersebar di laut beriklim sedang dan tropis. Di Laut Mediterania, dua spesies dari famili ini telah dilaporkan: Centrostephanus longispinus asli (Philippi, 1845) dan Diadema setosum non-asli (Leske, 1778). Meskipun tingkat perlindungannya (Tabel 1 ) dan berbagai tekanan yang dihadapinya ( S1 , Tabel 1), studi biogeografis pada C. longispinus masih sedikit, dan penilaian keseluruhan Status Konservasinya di Negara-negara UE Mediterania terutama didasarkan pada penilaian ahli daripada survei kuantitatif (Pelaporan berdasarkan Pasal 17 dari Arahan Habitat). Tidak adanya garis dasar pada distribusi spasiotemporal invertebrata bentik mempersulit penerapan strategi konservasi yang tepat berdasarkan biologi dan ekologi spesies (Stranga et al. 2024 ). Selain itu, C. longispinus dapat dianggap sebagai spesies bendera dari kumpulan koraligen, yang berkontribusi pada penataan habitat dengan mengatur pertumbuhan alga dan menyediakan tempat berlindung bagi organisme yang lebih kecil (Andrew dan Byrne 2007 ; Guallart dan Templado 2012 ). Kehadirannya dianggap sebagai indikator integritas habitat koraligen (La Mesa et al. 2021 ). Karena daya tarik estetikanya, bulu babi ini juga berfungsi sebagai simbol yang berharga untuk meningkatkan kesadaran publik tentang pentingnya ekosistem mesofotik beriklim sedang (Cerrano et al. 2019 ).
Genus Diadema Gray, 1825, merupakan salah satu kelompok bulu babi tropis yang paling melimpah, tersebar luas, dan penting secara ekologis yang menghuni lingkungan laut dangkal (Bronstein dan Kroh 2018 ). Genus ini terdiri dari delapan spesies, termasuk D. setosum (Leske, 1778), yang diperkenalkan di Laut Mediterania pada tahun 2006 dan telah dianggap invasif sejak tahun 2018 (Zirler, Leck, et al. 2023 ; Zirler, Schmidt, et al. 2023 ). Beberapa jalur pengenalan telah diusulkan (Yokes dan Galil 2006 ), termasuk transportasi larva melalui Terusan Suez, perdagangan akuarium, dan air pemberat (Zirler, Leck, et al. 2023 ; Zirler, Schmidt, et al. 2023 ). Yang awalnya dimulai sebagai penampakan sporadis individu-individu yang terisolasi atau kelompok-kelompok kecil telah meningkat menjadi pengamatan puluhan (Katsanevakis et al. 2020 ; Artüz dan Artüz 2019 ) dan dalam beberapa kasus, ratusan individu (Vafidis et al. 2021 ), yang menggambarkan penyebaran cepat D. setosum di seluruh Mediterania timur (Zirler, Leck, et al. 2023 ; Zirler, Schmidt, et al. 2023 ). Sebagai penggembala unggul kompetitif yang penting, D. setosum dapat menyebabkan perubahan signifikan dalam kumpulan komunitas bentik autochthone. Proliferasinya dapat menyebabkan berkurangnya populasi bulu babi asli seperti Arbacia lixula (Linnaeus, 1758) dan Paracentrotus lividus (Lamarck, 1816) di daerah yang padat koloni (Vafidis et al. 2021 ; Zirler, Leck, et al. 2023 ; Zirler, Schmidt, et al. 2023 ). Sementara perubahan iklim dan tekanan antropik juga dapat berkontribusi terhadap penurunan bulu babi asli (Martino et al. 2021 ), perluasan pesaing kuat seperti D. setosum dapat lebih menghambat pemulihannya (Zirler, Leck, et al. 2023 ; Zirler, Schmidt, et al. 2023 ). Selain itu, penggembalaannya yang intensif tidak hanya mengurangi tutupan alga tetapi juga dapat mengubah substrat, sehingga menghambat pemukiman spesies merobenthos (Sammarco 1980 ). Masalah ekologi utama yang terkait dengan penyebaran D. setosum meliputi degradasi habitat, penipisan sumber daya, dan potensi hibridisasi (Zirler, Leck, et al. 2023 ; Zirler, Schmidt, et al. 2023 ). Namun, masih belum ada protokol pemantauan untuk spesies tersebut, yang sangat penting untuk mengelola dan mengurangi perluasannya (Huseyinoglu et al. 2024 ).
Sampai saat ini, baik Diadema africanum (Rodriguez et al. 2013 )—yang hidup berdampingan dengan C. longispinus di Atlantik timur laut, dari Kepulauan Madeira hingga Teluk Guinea (Rodriguez et al. 2013 )—maupun Diadema savignyi (Audouin, 1809), yang bersimpati dengan D. setosum , belum tercatat di Laut Mediterania. Namun, kepadatan tinggi D. africanum di Kepulauan Canary telah menyebabkan penurunan yang nyata dalam tutupan makroalga dan keanekaragaman hayati terkait (Rodriguez et al. 2013 ). Mengingat potensi D. africanum untuk hadir setidaknya di dekat Selat Gibraltar, sensus distribusinya diperlukan. Demikian pula, perhatian yang lebih besar harus diberikan kepada D. savignyi dari Laut Merah (Zeina et al. 2016 ), yang berpotensi memasuki cekungan dari Terusan Suez. Pengumpulan data terstandarisasi tentang Diadematidae sangat penting untuk menilai status konservasi spesies asli dan untuk mengevaluasi perluasan bulu babi asing.
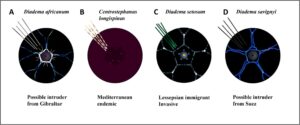
Saat ini, tidak ada materi edukasi visual yang membantu membedakan C. longispinus , D. setosum , D. savignyi, dan D. africanum . Kesenjangan pengetahuan ini dapat menyebabkan kesalahan identifikasi, yang mengakibatkan penilaian yang tidak akurat terhadap ukuran dan distribusi populasi, dan kemungkinan intervensi pengelolaan yang tidak tepat (misalnya, pemusnahan, Austen et al. 2016 ), khususnya selama survei Citizen Science (CS) (Encarnação et al. 2021 ).
Tujuan utama dari pekerjaan ini adalah (i) untuk memberikan sintesis ciri morfologi dan ekologi utama bulu babi Diadematidae untuk mencegah kesalahan identifikasi selama pengumpulan data, (ii) untuk menetapkan distribusi spasiotemporal bulu babi asli dan invasif saat ini, memperoleh tonggak sejarah untuk rencana pemantauan dan pengelolaan standar di masa mendatang, (iii) untuk mengidentifikasi area prioritas untuk konservasi C. longispinus dan pemantauan D. setosum melalui pemodelan distribusi, dan (iv) untuk menerapkan protokol sederhana untuk melatih orang mengidentifikasi spesies, mendapatkan catatan baru melalui CS dan memperoleh lebih banyak—dan lebih akurat—data spasial skala besar tentang bulu babi diadematoid.
2 Bahan dan Metode
2.1 Ciri Morfologi dan Ekologi Spesies Diadematoid yang Dipertimbangkan
Untuk membantu identifikasi yang benar, karakter diagnostik utama spesies Diadematidae Centrostephanus longispinus dan Diadema setosum dirangkum, yang memudahkan penerapan protokol pelatihan CS. Meskipun saat ini tidak ditemukan di Laut Mediterania, karakteristik morfologi utama D. africanum dan D. savignyi juga disajikan untuk mencegah potensi kesalahan identifikasi dengan spesies asli.
2.2 Distribusi Spesies Diadematidae
2.2.1 Sumber Data
Bahasa Indonesia : Tinjauan komprehensif catatan kejadian untuk Centrostephanus longispinus dan Diadema setosum di Laut Mediterania dilakukan hingga 17 Januari 2025. Penelitian ini menggunakan berbagai sumber data, termasuk literatur (diperoleh dari platform Web of Science [WoS]), Web Ecological Knowledge (WEK, Di Camillo et al. 2018a ), Marine Biodiversity Platforms (MBP, misalnya, Global Biodiversity Information Facility [GBIF, www.gbif.org ], Ocean Biodiversity Information System [OBIS, www.obis.org ], Données d’Observations pour la Reconnaissance et l’Identification de la faune et la flore Subaquatiques [DORIS, www.doris.ffessm.fr ], European Nucleotide Archive [ENA, www.ebi.ac.uk ]) dan platform CS laut (iNaturalist, misalnya, inaturalist.org ; Reef Check Med, www.reefcheckmed.org ; Dive4Diadema, https://dive4diadema.org/ ). Kata kunci yang digunakan untuk mencari sumber yang disebutkan di atas adalah Centrostephanus dan ‘Mediterania’ untuk spesies endemik, dan Diadema dan ‘Mediterania’ untuk echinoid asing. Catatan dari iNaturalist dan Doris disertakan dalam basis data GBIF, dan sumber asli hanya dikonsultasikan untuk memverifikasi catatan melalui foto. WEK merujuk pada data ilmiah yang tidak sengaja dibagikan di World Wide Web oleh jutaan internaut yang tidak terlatih (Di Camillo et al. 2018a ), yang mewakili sumber informasi gratis, sering kali disertai dengan foto atau video yang bermanfaat. Data ini diverifikasi dan divalidasi. Catatan yang diperoleh dari MBP ditinjau dengan cermat untuk menghindari redundansi di seluruh platform dan difilter untuk mengecualikan yang terkait hanya dengan spesimen yang diawetkan. Catatan lebih lanjut dikumpulkan oleh penulis melalui survei bawah air terarah yang dilakukan di Kepulauan Tremiti dari 1 Juli hingga 30 September 2023 (lihat paragraf khusus di depan), serta dari penampakan sesekali di bagian lain Laut Mediterania. Untuk setiap penampakan, koordinat geografis, beserta semua informasi yang tersedia (misalnya, kedalaman pengambilan sampel, suhu, jumlah individu, warna, paparan, dan jenis substrat) dikompilasi ke dalam lembar kerja standar mengikuti Di Camillo et al. (2018b). This dataset, along with its associate metadata, is provided as supplementary materials (https://doi.org/10.5281/zenodo.15209518). When available, photographs of sea urchins were analysed to validate the records and to gather additional ecological and morphological information (i.e., colour morph).
2.2.2 Model Distribusi Spesies
Seluruh dataset diubah menjadi file teks dan digunakan untuk membangun peta distribusi spesies melalui perangkat lunak QGIS ( https://www.qgis.org/it/site/ ). Sebelum menerapkan model prediktif untuk menilai pola distribusi spasiotemporal dari dua bulu babi, analisis eksploratori dilakukan untuk mengevaluasi frekuensi setiap variabel yang dikumpulkan (misalnya, suhu, kedalaman, tahun, substrat; deskripsi lapangan, https://doi.org/10.5281/zenodo.15209518 ) di antara spesies yang berbeda. Langkah awal ini membantu mengidentifikasi pendekatan statistik yang paling sesuai. Untuk memperkirakan distribusi spasial Centrostephanus dan Diadema , estimasi kepadatan kernel dilakukan dengan menggunakan paket ‘terra’ di R (versi 4.3.3, Hijmans 2024 ). Data distandarisasi menjadi matriks ada/tidaknya, di mana keberadaan pusat penyelaman, tetapi tidak ada penampakan spesies target yang tercatat diperlakukan sebagai ‘0’. Dataset diproyeksikan ke grid 0,5 × 0,5° yang mencakup Laut Mediterania, menggabungkan penampakan spesies dan jumlah pusat penyelaman. Pendekatan ini memungkinkan penyertaan area tempat spesies tersebut mungkin ada tetapi tidak terdeteksi karena aktivitas penyelaman terbatas atau tidak ada. Untuk memodelkan keberadaan spesies, Generalized Additive Models (GAMs) dengan distribusi binomial dan fungsi tautan logit diterapkan (Hijmans 2023 ). Untuk Centrostephanus , model awal mencakup variabilitas spasial (lintang dan bujur, x,y), jenis substrat, sumber data (misalnya, CS, WEK dan MBP) dan jumlah pusat penyelaman (NUM_DIVING) di setiap sel grid sebagai offset. Membandingkan model dengan dan tanpa jenis substrat dan sumber data menunjukkan bahwa kovariat ini meningkatkan deviasi yang dijelaskan dari 47% menjadi 58,3%. Mengingat peningkatan dalam daya penjelasan model ini, variabel-variabel ini dipertahankan dalam model akhir untuk Centrostephanus . Modelnya ditetapkan sebagai
![]()
di mana PA = Kehadiran Ketidakhadiran. Sebaliknya, model yang lebih sederhana diterapkan untuk Diadema , hanya mencakup variabel spasial (lintang dan bujur) dan tahun sebagai istilah yang halus, karena kurangnya substrat dan data sumber untuk sebagian besar penampakan. Model untuk Diadema adalah
![]()
Akhirnya, prediksi spasial distribusi spesies dibuat untuk kedua genera di seluruh cekungan Mediterania.
2.2.3 Implementasi dan Pengujian Protokol Pengumpulan Data Standar oleh CS
Mengingat potensi CS dalam mengumpulkan sejumlah besar data keanekaragaman (Lucrezi et al. 2018 ), sebuah catatan penyelaman sederhana dan protokol pelatihan untuk pemantauan bawah air bulu babi Diadematidae dikembangkan ( S2 , Protokol pemantauan). Protokol CS ini dapat mengintegrasikan metodologi pemantauan yang ada untuk C. longispinus (La Mesa et al. 2021 ), yang didasarkan pada sensus visual bawah air yang dilakukan dalam batas tanpa dekompresi penyelaman SCUBA rekreasi (hingga kedalaman 40 m). Protokol yang diusulkan mencakup sedikit penyesuaian untuk merekam individu yang terisolasi (setidaknya berjarak 20 cm) versus agregasi (individu berjarak <20 cm), mengingat kesulitan yang lebih besar untuk memperoleh data yang akurat dalam agregasi yang padat. Penyelam disarankan untuk tidak menyentuh atau memindahkan bulu babi selama survei, baik untuk menghindari kerusakan pada hewan dan mencegah cedera yang menyakitkan pada diri mereka sendiri. Seperti yang dijelaskan secara rinci di S2 , untuk setiap pengamatan bulu babi yang terisolasi, penyelam harus mencatat spesies, warna, konteks lingkungan (didefinisikan menurut protokol Reef Check Med Underwater Coastal Environment, Turicchia et al. 2021a , 2021b ), suhu, kedalaman, dan apakah individu tersebut tersembunyi atau terekspos. Daftar penyelaman mencakup penggaris untuk memudahkan pengukuran diameter uji (dalam cm). Untuk agregasi, penyelam harus memperkirakan kelimpahan dengan memilih kategori kelimpahan (Turicchia et al. 2021a , 2021b ) (detail di S2 ). Untuk menguji protokol, dan mengumpulkan catatan tambahan dari spesies endemik, populasi C. longispinus dipelajari melalui penyelaman SCUBA di 11 lokasi dalam Kawasan Konservasi Laut (KKL) Pulau Tremiti, Foggia, Apulia, Italia ( S1 , Gambar 1). Dengan dukungan pusat penyelaman lokal, 100 penyelam rekreasi dilatih untuk mengidentifikasi Diadematidae Mediterania dan mengumpulkan data. Setiap lokasi disurvei setidaknya tiga kali. Selain data CS, fotoquadrat berukuran 50 × 50 cm digunakan untuk mengkarakterisasi substrat di dua lokasi dengan jumlah penampakan tertinggi— Cala dei Turchi dan Secca della Vedova —menggunakan kamera Olympus TG-6. Lima belas fotoquadrat per lokasi diambil secara acak pada kedalaman 25–30 m, dengan panjang sekitar 10 m, yang mencakup total area pengambilan sampel seluas 50 m 2Komposisi substrat (rata-rata persentase tutupan ± SD) dinilai menggunakan pemrosesan citra digital, dengan mengkategorikan enam kelompok bentik utama: Rumput laut, Hexacorallia, Porifera, Polychaeta, Tunicata, dan Bryozoa, beserta dua kategori tambahan: Tidak teridentifikasi dan Lainnya. Kategori ‘Lainnya’ mencakup komponen minor atau campuran, seperti rumput alga, skleraktinia soliter, dan ascidia kecil soliter, yang tidak dapat dibedakan dengan baik dalam citra.
3 Hasil
3.1 Ciri Morfologi dan Ekologi Spesies Diadematoid yang Dipertimbangkan
Deskripsi terperinci tentang Centrostephanus longispinus, Diadema setosum , D. africanum, dan D. savignyi disediakan di S1 . Fitur diagnostik utama dirangkum dalam Gambar 1A–D dan S1 , Tabel 2–4.
3.2 Distribusi Spesies Diadematidae
3.2.1 Sumber Data
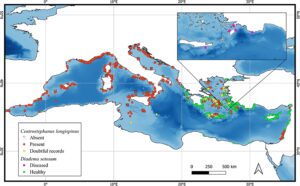
Sebanyak 3314 catatan ( https://doi.org/10.5281/zenodo.15209518 ) diambil dari berbagai sumber: WEK (3% dari jumlah total catatan), CS (36%), MBP (25%), literatur (33%) dan karya saat ini (3%). Di antara ini, 2220 catatan adalah C. longispinus , termasuk catatan kehadiran ( n = 1653, 74%) dan ketidakhadiran ( n = 567, 26%), sementara 1094 adalah catatan kehadiran D. setosum (Tabel S5 ). Data tersebut mencakup dari tahun 1955 (ketika C. longispinus pertama kali dilaporkan dalam literatur oleh Cherbonnier 1956 ) hingga Januari 2025. Sebagian besar rekaman untuk C. longispinus diperoleh melalui CS (953, termasuk 816 dari Reef Check Med) dan MBP (801, 723 di antaranya dari GBIF), dibandingkan dengan 306 dari literatur. Sebaliknya, sebagian besar rekaman untuk D. setosum berasal dari literatur (775) ( S1 , Tabel 5). Mayoritas rekaman C. longispinus berasal dari Italia (1091, 49%) dan Prancis (808, 36%), dengan sekitar 300 rekaman didistribusikan di negara-negara Mediterania lainnya. Menganalisis distribusi sumber ( S1 , Tabel 6 dan Gambar 2), data Prancis terutama diperoleh dari MBP (716), sedangkan rekaman dari pantai Italia sebagian besar berasal dari CS (759). Catatan D. setosum sebagian besar terpusat di empat negara: Yunani (420 catatan, 38%), Israel (219, 20%), Siprus (212, 19%) dan Turki (225, 21%). Di wilayah-wilayah ini, D. setosum hidup berdampingan dengan C. longispinus , meskipun dengan jumlah penampakan yang jauh lebih banyak (1076 catatan untuk D. setosum vs. 115 untuk C. longispinus ). Peta distribusi catatan ada-tidaknya (Gambar 2 ) menunjukkan bahwa penampakan D. setosum yang terkonfirmasi paling utara hingga saat ini adalah Dardanelles, Teluk Saros (Lat: 40.59278°N, Lon: 26.51278°E, Artüz dan Artüz 2019 ) dan Pulau Paxos di Yunani (Laut Ionia, Lat: 39.20275°N, Lon: 20.18698°E). Catatan yang diberi label ‘berpenyakit’ merujuk pada spesimen D. setosum yang terkena dampak peristiwa kematian massal 2022–2023 di beberapa lokasi, termasuk Pelabuhan Kastellorizo (Yunani), Teluk Anthony Quinn (Rhodes), Pulau Alimia, Gökova, Fethiye, dan Muğla (Turki) (Zirler, Leck, et al. 2023 ; Zirler, Schmidt, et al. 2023 ; Dinçtürk et al. 2024 ). Masih banyak daerah—termasuk Siprus, Lebanon, Suriah, Türkiye, Libya, dan Mesir—di mana keberadaan maupun ketiadaan C. longispinus belum dilaporkan. Meskipun C. longispinustelah tercatat sejak 1950-an, sebagian besar catatan (1198) dilaporkan mulai 2013 dan seterusnya. Demikian pula, catatan D. setosum mulai meningkat dari 2010, dengan puncak yang nyata antara 2020 dan Agustus 2024 ( S1 , Gambar 3, 4). Frekuensi tahunan catatan untuk kedua spesies disajikan dalam S1 , Gambar 5. Data paparan untuk bulu babi tersedia dalam jumlah kasus yang terbatas—sekitar 12% dari catatan C. longispinus dan 25% dari catatan D. setosum . Ketika informasi ini dilaporkan, C. longispinus tidak menunjukkan perbedaan substansial antara individu yang terpapar dan tersembunyi (terpapar = 52%; tersembunyi = 48%, dari total kejadian kehadiran). Sebaliknya, D. setosum sebagian besar diamati pada posisi terpapar (terpapar: 78%; tersembunyi: 22%). Kedua spesies ini terutama berasosiasi dengan substrat keras alami ( C. longispinus : 90%; D. setosum : 92%). Proporsi penampakan yang lebih kecil terjadi pada substrat lunak ( C. longispinus : 2%; D. setosum : 1%), substrat campuran ( C. longispinus : 5%; D. setosum : 2%) atau struktur buatan ( C. longispinus : 2%; D. setosum : 5%), seperti bangkai kapal dan anjungan gas. Kisaran batimetri untuk kedua spesies dirangkum dalam Tabel 2. Meskipun data kedalaman tidak tersedia di semua catatan, sebagian besar penampakan terjadi di perairan yang relatif dangkal (hingga 40 m). Beberapa penelitian melaporkan kejadian pada kedalaman yang lebih dalam, baik oleh Kendaraan yang Dioperasikan dari Jarak Jauh (ROV) (Leonard et al. 2020 ; Bonhomme et al. 2014 ; Toma et al. 2024 ) atau melalui pengambilan sampel pukat (Templado dan Moreno 1996 ; Cherbonnier 1956 ). Agregasi besar dari 344 individu C. longispinus didokumentasikan oleh ROV pada kedalaman antara 45 dan 96 m di sepanjang jaringan pipa antara Port-Miou Cove dan kepala Cassidaigne Canyon (Provence Barat dan Timur) (Bonhomme et al. 2014 ).
| Pohon Centrostephanus longispinus | Diadema setosum | |
|---|---|---|
| Kedalaman minimum | 8 m (EM) a , 3 m (WM) b | 0,20 m (EM ) |
| Kedalaman maksimum | 80 m (EM) c , 550 m (WM) d | 55 m (EM) untuk |
Simboura dkk., 1995. b Templado dan Moreno, 1996. c dari GBIF, ID IL-EchCL-2018-0001787, S1. d Moya-Urban et al. 2025 (yang memperbarui rekor sebelumnya pada 379 m oleh Leonard et al. 2020 ). dan Zirler, Leck, dkk. 2023 . f Katsanevakis dkk. 2020 .
Perbandingan distribusi batimetri dan temporal catatan kemunculan C. longispinus dan D. setosum —khususnya di area tempat kedua spesies tersebut hidup berdampingan (misalnya, pantai Yunani dan Turki serta cekungan Levant)—mengungkapkan tumpang tindih kedalaman parsial antara 10 dan 40 m. Namun, catatan C. longispinus telah menurun setelah tahun 2016, sementara penampakan D. setosum mencapai puncaknya pada tahun 2021 dan 2022, dengan jumlah maksimum 287 catatan ( S1 , Gambar 6 A,B).
3.2.2 Model Distribusi Spesies
Catatan Diadema setosum hanya terdiri dari data kehadiran, sedangkan catatan untuk Centrostephanus longispinus mencakup pengamatan kehadiran dan ketidakhadiran, dengan ketidakhadiran mencakup 25,5% dari kumpulan data. Data tambahan tentang fitur abiotik (misalnya, Suhu atau Kedalaman) terbatas untuk kedua spesies. Hanya 92 dari 2220 untuk C. longispinus dan 34 dari 1094 catatan untukD. setosum included Temperature information. Depth data were slightly more frequent, with 127 records for C. longispinus and 481 for D. setosum reporting this variable.
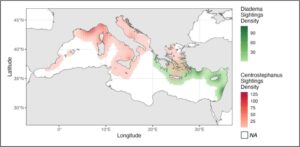
Peta kepadatan menunjukkan pola distribusi yang berbeda antara kedua spesies tersebut. C. longispinus lebih umum ditemukan di Mediterania barat laut, sedangkan D. setosum lebih sering ditemukan di bagian timur cekungan (Gambar 3 ).
Untuk memprediksi distribusi spasiotemporal C. longispinus dan Diadema setosum di Laut Mediterania, kami menerapkan GAM dengan distribusi binomial dan fungsi tautan logit (Gambar 4A,B ). Model tersebut menggabungkan kovariat spasial dan temporal. Untuk C. longispinus , model tersebut juga menyertakan faktor lingkungan kualitatif (tipe substrat: Keras, Campuran, Tidak Diketahui, Posidonia , dan Lunak) dan sumber data (WEK, Literatur, MBP, dan Karya Sekarang) untuk mengevaluasi pengaruhnya terhadap keberadaan spesies. Untuk D. setosum, variasi spasial menunjukkan efek positif signifikan pada kemunculan spesies ( p < 0,001), sedangkan efek temporal tidak signifikan. Model tersebut menjelaskan 98,1% deviasi, dengan nilai R -kuadrat yang disesuaikan sebesar 0,985.
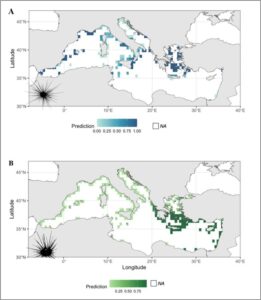
Untuk C. longispinus, istilah spasial sangat signifikan dan positif ( p < 0,001), seperti halnya istilah temporal ( p < 0,001). Di antara prediktor kategoris, jenis substrat tertentu (misalnya, Keras dan Campuran) memiliki efek signifikan pada keberadaan spesies, sementara yang lain (misalnya, Posidonia , Lunak) tidak. Sumber data juga memainkan peran penting, dengan kontribusi dari beberapa kumpulan data (misalnya, MBP, Present Work, dan WEK) yang memengaruhi prediksi.
3.3 Implementasi dan Pengujian Protokol Pengumpulan Data oleh CS
Selama periode 3 bulan, 97 catatan ada/tidaknya Centrostephanus longispinus dikumpulkan dari Daerah Konservasi Laut Kepulauan Tremiti menggunakan protokol CS yang diusulkan. Hingga lima bulu babi per survei diamati di sembilan dari 11 lokasi penyelaman yang dikunjungi ( S1 , Tabel 7), yang semuanya dieksplorasi selama siang hari. Sebagian besar penampakan terjadi pada kedalaman antara 20 dan 30 m, dengan individu ditemukan baik yang terekspos (37%) maupun tersembunyi (58%). Lokasi dengan jumlah individu tertinggi yang tercatat adalah Cala dei Turchi (5 per survei), yang kedalamannya berkisar antara 16,9 hingga 32,7 m. Karakteristik utama setiap lokasi dirinci dalam S1 , Tabel 8. Hampir semua lokasi utara dimulai dari tebing vertikal yang berakhir dengan substrat horizontal yang terdiri dari campuran batu dan pasir. Di Secca di Punta Secca dan Cala Caffè, dinding berbatu memanjang hingga kedalaman 40 m atau lebih. Sebaliknya, lokasi selatan pulau tersebut dikarakterisasi oleh lereng landai dan formasi batuan pesisir. Karakterisasi substrat menggunakan fotokuadrat di dua area terpilih ( S1 , Gambar 7), mengungkapkan persentase tutupan berikut untuk kategori Lainnya: 81% ± 11% SD di Cala dei Turchi dan 60% ± 21% SD di Secca della Vedova . Kategori ini mencakup rumput alga, skleraktinia soliter, dan ascidia soliter kecil. Tutupan rumput laut adalah 34,5% ± 25,6% SD di Secca della Vedova dan 7,2% ± 10% SD di Cala dei turchi , sementara tutupan Porifera adalah 8,25% ± 8% SD di Cala dei Turchi, dan 3,5% ± 9% SD di Secca della Vedova . Organisme paling melimpah yang ditemukan di kuadrat termasuk Peyssonnelia spp. dan spons seperti Haliclona sp., Ircinia sp., Chondrosia reniformis , dan Phorbas sp., yang merupakan jenis spons khas lingkungan dengan cahaya redup seperti gua dan habitat koraligi.
4 Diskusi
Studi ini memberikan wawasan awal tentang distribusi bulu babi diadematoid asli ( Centrostephanus longispinus ) dan asing ( Diadema setosum ) di Laut Mediterania, yang mewakili dasar penting untuk penilaian biogeografis status konservasi bulu babi yang dilindungi, serta untuk mengevaluasi risiko invasi dan mengoptimalkan upaya pengelolaan untuk spesies invasif. Area prioritas yang diidentifikasi oleh penelitian ini untuk menerapkan kegiatan pengawasan dan langkah-langkah konservasi meliputi pantai Aegea, Ionia, dan Levantina. Mirip dengan spesies invasif lainnya, penyebaran D. setosum dimulai di wilayah ini, dan kelimpahannya yang meningkat dapat berkontribusi pada hilangnya keanekaragaman hayati. Variasi kepadatan populasi echinoid sering dikaitkan dengan gangguan antropogenik dan perubahan iklim (Uthicke et al. 2009 ); dengan demikian, ekosistem laut di wilayah Ionia dan Aegea-Levantina berada di bawah tekanan kumulatif dan sangat membutuhkan tindakan perlindungan (Fanelli et al. 2021 ).
D. setosum tampaknya tersebar luas di hampir semua negara Mediterania tenggara, dengan titik-titik panas di Turki, Aegea selatan, dan Siprus, yang membentang dari pantai Levant Timur ke arah Lebanon dan Israel. Batas paling utara di pantai Ionia telah diperbarui, yang menunjukkan bahwa spesies asing tersebut mungkin juga ada di Corfu atau dekat perbatasan Albania. Mengingat bahwa beberapa spesies invasif yang tercatat di Laut Ionia telah diamati di Laut Adriatik tengah timur dalam beberapa tahun (misalnya, Pterois miles (Bennett, 1828), Dragičević et al. 2021 ), protokol peringatan dini harus diterapkan di bagian cekungan ini untuk memverifikasi keberadaan D. setosum . D. setosum menunjukkan ciri-ciri penyerbu sukses berukuran sedang dan berumur sedang (Quell et al. 2021 ): Ia memiliki masa hidup melebihi 3 tahun (Vafidis et al. 2021 ; Zirler, Leck, et al. 2023 ), mencapai diameter uji sekitar 97 mm (Zirler, Leck, et al. 2023 ), sangat subur (menghasilkan sekitar 3 juta telur per betina dalam kondisi terkendali) dan dapat menyesuaikan siklus hidupnya sebagai respons terhadap faktor lingkungan (Muthiga dan McClanahan 2007 ). Lengan postoral yang panjang dan berfenestrasi dari larva planktotrofik D. setosum (Pearse dan Cameron 1991 ; Uthicke et al. 2009 ) dapat meningkatkan peluang kelangsungan hidup plutei di kolom air (dengan rentang hidup larva 40–45 hari, Aminur Rahman et al. 2015 ) dan mendorong pertumbuhan eksponensial D. setosum (Pechenik 1999 ). Bulu babi membentuk agregasi besar yang mungkin menghalangi predasi ikan dan menyinkronkan gametogenesis jantan dan betina (Muthiga dan McClanahan 2007 ). Pematangan gonad terjadi pada 25°C (Bronstein et al. 2016 ), sedangkan suhu optimal untuk perkembangan larva adalah 28°C (Sarifudin et al. 2016 ; Aminur Rahman et al. 2015 ), yang menunjukkan bahwa keberhasilan imigran Lessepsian ini juga terkait dengan dampak perubahan iklim dan peningkatan suhu (Hernández et al. 2020 ). Sifat utama lain yang berkontribusi terhadap sifat invasif D. setosum adalah kemampuannya untuk hidup di pelabuhan (Pearse 1998 ; dan pengamatan pribadi), yang menyoroti peran pelabuhan tidak hanya sebagai batu loncatan bagi spesies asing (Outinen et al. 2021 ) tetapi juga sebagai inkubator untuk tahap awal kehidupan mereka. D. setosum dan C. longispinus menunjukkan sifat ekologi yang berbeda, sedangkanD. setosum terutama menghuni perairan dangkal pada substrat yang terbuka dan cenderung berkumpul (Muthiga dan McClanahan 2007 ), C. longispinus umumnya menyendiri, peka cahaya, nokturnal (Andrew dan Byrne 2007 ) dan terkait dengan singkapan koraligen yang dalam. Mengingat sifat menguntungkan D. setosum di perairan hangat, kemungkinan besar bulu babi invasif ini dapat bersaing dengan C. longispinus pada kedalaman mulai dari 10 hingga 40 m (misalnya, di mana distribusinya tumpang tindih) dan dengan bulu babi air dangkal asli lainnya seperti Paracentrotus lividus (Lamarck, 1816) dan Arbacia lixula (Lamarck, 1758) (Zirler, Leck, et al. 2023 ). Sebaliknya, penipisan spesies asli dapat dipercepat oleh pemanasan cepat di cekungan (Yeruham et al. 2015 ).
Mengenai faktor-faktor yang membatasi perluasan D. setosum , spesies yang memakan bulu babi juga dapat memangsa Diadematidae, seperti Charonia tritonis (Moluska: Gastropoda) yang termofilik, yang baru-baru ini berkembang biak di Laut Mediterania (Hall et al. 2017 ), serta bintang laut (misalnya, Marthasterias glacialis ) dan spesies ikan asli ( Diplodus sargus dan Balistes capriscus Guallart dan Templado 2012 ) dan non-asli ( Lagocephalus sceleratus dan Torquigener hypselogeneion Zirler, Leck, et al. 2023 ; Zirler, Schmidt, et al. 2023 ). Selain itu, aktivitas herbivori intensif dari ikan kelinci invasif Siganus spp. (Vella et al. 2023 ) dapat semakin mengurangi sumber makanan untuk D. setosum di substrat dangkal. Kematian massal yang sedang berlangsung yang mempengaruhi D. setosum menipiskan populasinya di daerah yang terkena dampak (Zirler, Schmidt, et al. 2023 ). Sementara Dinçtürk et al. ( 2024 ) mengaitkan Vibrio spp. sebagai penyebab kematian di Turki, masih belum pasti apakah patogen potensial ini asli atau asing, dan tidak jelas apakah itu menimbulkan risiko bagi bulu babi endemik atau spesies lain di Mediterania. C. longispinus terutama telah diamati di Mediterania barat; sedikit catatan C. longispinus di laut Aegea, Ionia, Adriatik timur, Levantina dan Mediterania tengah-timur. Ini menunjukkan bahwa penelitian lebih lanjut diperlukan untuk menentukan apakah C. longispinus benar-benar langka di cekungan ini atau lebih umum pada kedalaman yang belum dieksplorasi (10–80 m). Titik panas utama untuk spesies yang diidentifikasi dalam studi ini adalah pantai Mediterania barat laut, yang harus diprioritaskan untuk perlindungan bulu babi dan untuk mempelajari dinamika populasinya. Distribusi C. longispinus sebagian mencerminkan habitat koralligen (EMODnet), tempat ia dapat menemukan tempat berlindung dan makanan, seperti rumput laut dan bryozoa serta spons yang berkerak (Guallart dan Templado 2012 ; Öndes et al. 2022 ; Andrew dan Byrne 2007 ). Selain itu, penemuan baru-baru ini tentang kepadatan tinggi C. longispinus di lapisan rhodolith (Tabone et al. 2024 ) menunjukkan bahwa penyelidikan lebih lanjut ke dalam habitat rhodolith/Maërl (Demestre et al. 2024 ) dapat menghasilkan catatan spesies baru.
Perubahan yang sedang berlangsung dalam sirkulasi termohalin di Laut Mediterania (Skliris et al. 2018 ) juga memengaruhi kemunculan bulu babi. Sementara Mediterania barat mengalami penguapan yang berlebihan, Laut Aegea Utara lebih rentan terhadap penyegaran (Skliris et al. 2018 ). Jika C. longispinus sensitif terhadap salinitas rendah dan suhu tinggi, mirip dengan kerabatnya C. rodgersii (Webb dan Byrne 2025 ), Mediterania barat laut dapat berfungsi sebagai tempat perlindungan iklim untuk bulu babi endemik ini. Hotspot untuk C. longispinus tumpang tindih dengan batas-batas Suaka Pelagos, Kawasan Lindung Khusus Penting Mediterania (SPAMI) untuk konservasi cetacea dan habitatnya, serta Kawasan Penting Secara Ekologis atau Biologis (EBSA) yang dikenal sebagai Ekosistem Bentik Mediterania barat laut ( www.cbd.int/ebsa ). Namun, inisiatif perencanaan tata ruang laut lintas batas yang komprehensif untuk wilayah ini, yang bertujuan untuk menyelaraskan tujuan ekonomi, sosial, dan ekologi, masih belum ada (Mackelworth et al. 2024 ). Wilayah ini juga memiliki sejumlah besar pusat penyelaman (Di Camillo et al. 2025 ), yang menunjukkan bahwa pemantauan spesies yang dilindungi dapat difasilitasi melalui keterlibatan penyelam rekreasi.
Karena sifat invasif Diadematidae, dua spesies bulu babi lainnya yang hidup di sekitar Mediterania dapat memasuki cekungan: Diadema africanum dari Atlantik dan Diadema savignyi dari Laut Merah. Tahap larva D. africanum dapat menempuh jarak yang jauh (Hernández et al. 2020 ; Cano et al. 2025 ), dan meskipun penghalang biogeografis yang diciptakan oleh front Almeria–Oran dapat menghambat penyebarannya ke dalam cekungan (Patarnello et al. 2007 ), penghalang tersebut mungkin ada setidaknya di dalam batas front ini. Pemanasan global dapat berkontribusi untuk melemahkan penghalang ini dan memfasilitasi perjalanan spesies tropis (Albano et al. 2024 ). D. savignyi dan D. setosum bersifat simpatrik (Pearse 1998 ; Shahid et al. 2015 ), tetapi perbedaan modalitas reproduksi dan periode pemijahannya kemungkinan memungkinkan terjadinya isolasi dan koeksistensi genetik (Muthiga 2003 ). D. savignyi belum memasuki cekungan Mediterania, kemungkinan karena (i) potensi hambatan dalam gametogenesis di lintang yang lebih dingin (Muthiga 2003 ) atau (ii) perbedaan dalam ekologi trofik dan/atau kemampuan beradaptasi yang lebih rendah dibandingkan dengan D. setosum , karena lebih menyukai terumbu karang yang terbuka, dan larvanya mungkin tidak dapat bertahan hidup di perairan pemberat atau pelabuhan (Pearse 1998 ). Meskipun beberapa penulis melaporkan bahwa D. savignyi banyak terdapat di Laut Merah (Aljahdali dan Molla 2022 ), hanya ada sedikit catatan dari Teluk Suez (GBIF), dan hanya satu gambar yang tersedia dari iNaturalist (diambil di Sudan), yang menunjukkan bahwa kemunculan D. savignyi mungkin terbatas pada beberapa area atau bahwa bulu babi tersebut mungkin salah diidentifikasi sebagai D. setosum .
Pedoman identifikasi yang disediakan di sini akan membantu dalam membedakan antara bulu babi asli dan invasif dan secara akurat mencatat keberadaan atau ketidakhadiran mereka, yang sangat penting untuk mengambil tindakan konservasi yang efektif. Beberapa bulu babi diadematoid, termasuk D. setosum dapat dimakan, dan beberapa penelitian sedang mengeksplorasi potensi untuk membudidayakan Diadema spp. di daerah asalnya (Dautov et al. 2020 ). Jika menjadi perlu untuk mengendalikan perluasan bulu babi non-asli di Mediterania, mereka dapat dianggap sebagai produk kerang, mirip dengan kepiting biru Atlantik Callinectes sapidus (Azzurro et al. 2024 ). Mengenai sumber data, CS telah berkontribusi secara signifikan untuk mengumpulkan data kemunculan spesies target, menggarisbawahi pentingnya dan potensi ilmu partisipatif dalam meningkatkan pengetahuan tentang keanekaragaman hayati laut dan mengumpulkan data ketidakhadiran (Coppari et al. 2024 ). Sebaliknya, WEK menghasilkan lebih sedikit catatan tetapi menyediakan dokumentasi fotografi yang berharga di seluruh cekungan, penting untuk memvalidasi keberadaan bulu babi di area yang hanya memiliki catatan data. Catatan dari MBP terutama berasal dari pantai Mediterania Prancis, kemungkinan karena banyaknya infrastruktur (yaitu, pusat selam) yang ada di Côte d’Azur dan Corsica (Di Camillo et al. 2023 ). Catatan dari literatur lebih banyak untuk D. setosum daripada untuk C. longispinus yang dilindungi , kemungkinan karena (i) distribusi kedalaman yang lebih dangkal dari spesies invasif, (ii) tingkat paparannya yang lebih tinggi dan (iii) meningkatnya minat pada imigran Lessepsian sejak 2006 (Bronstein dan Kroh 2018 ; Zirler, Leck, et al. 2023 ; Zirler, Schmidt, et al. 2023 ; Vafidis et al. 2021 ). Pemantauan spesies invasif, yang memerlukan keterlibatan pemangku kepentingan dan publik (Kleespies et al. 2024 ), dapat menarik perhatian pada spesies asli dan meningkatkan tingkat perlindungan mereka, setidaknya di area yang dihuni oleh kedua spesies. Sebagian besar catatan bersifat baru, kemungkinan karena (i) meningkatnya jumlah inisiatif CS dalam beberapa tahun terakhir, (ii) tersedianya teknologi yang terjangkau untuk pencitraan bawah air, dan (iii) kemampuan untuk berbagi data secara daring. Memang, sebagian besar pengamatan berasal dari penyelaman skuba rekreasi, yang menunjukkan bahwa penyelaman teknis (baik oleh penyelam ilmiah maupun sukarelawan) dapat membantu menjembatani kesenjangan pengetahuan ini dengan memperluas eksplorasi manusia hingga kedalaman 90–100 m (Di Camillo et al. 2023 ), sementara pengumpulan data yang lebih dalam dan nondestruktif dapat dilakukan menggunakan ROV (Leonard et al. 2020 ; Bonhomme et al. 2014)).
The GAM provides valuable insights into the spatial dynamics of species presence/absence, highlighting significant periods of change and areas of high uncertainty. By incorporating the presence of diving centres in each grid cell, we accounted for potential sampling biases and ensured a more robust prediction. This approach allowed us to differentiate between true absence and lack of sampling effort, resulting in more accurate spatial prediction. However, due to the variability in data collection efforts and inconsistencies in presence and abundance records, the models are not robust enough to draw definitive conclusions about temporal trends in species presence/absence. It is crucial to refine data collection with a standardized approach and to include detailed information on the sightings, such as depth and temperature at the finding depth. Gathering further information would enhance the model’s predictive power. Nevertheless, other studies estimating the spatial distribution of sea urchin species in other regions (e.g., Canary Islands, Gonzàlez-Irusta et al. 2015) confirm that presence/absence models, such as binomial GAM, outperform better than those based solely on presence data. The lack of comprehensive information regarding sighting dates further undermines the reliability of the temporal predictions.
Kesimpulan
Studi ini memberikan penilaian tolok ukur distribusi spasial dan batimetri Diadematidae Centrostephanus longispinus dan Diadema setosum di Laut Mediterania. Temuan-temuan ini penting untuk meningkatkan efektivitas perencanaan konservasi untuk bulu babi yang dilindungi dan untuk mengelola spesies asing. Meskipun saat ini tidak mungkin untuk memperkirakan ukuran populasi bulu babi yang dilindungi, C. longispinus , bukti menunjukkan bahwa itu kurang langka daripada yang diyakini sebelumnya dan memiliki rentang distribusi yang menguntungkan. Kemunculan spesies asli ini pada biokonkresi berkapur autokton menyoroti bahwa konservasinya terkait erat dengan integritas habitat koraligen dan lapisan rhodolith. Mediterania barat laut telah muncul sebagai hotspot untuk C. longispinus , tetapi upaya pemantauan juga harus diprioritaskan di laut Aegea dan Ionia—di mana ia hidup berdampingan dengan D. setosum yang invasif —serta di wilayah Adriatik timur, Levantina, dan Mediterania timur-tengah, di mana datanya hilang. D. setosum tampaknya sudah mapan di Laut Mediterania bagian timur; namun, penelitian lebih lanjut diperlukan untuk mengeksplorasi distribusi batimetrisnya, penyebab wabah penyakit yang sedang berlangsung, dan dinamika ekologisnya di area yang tumpang tindih dengan C. longispinus . Meskipun jumlah catatan yang dikumpulkan tinggi, informasi ekologis tentang kedua bulu babi ini masih belum lengkap, terutama di ekosistem mesofotik. Untuk mengatasi hal ini, kami menyajikan protokol pemantauan standar yang dapat diterapkan oleh peneliti dan ilmuwan warga, yang bertujuan untuk menghasilkan data yang lebih konsisten tentang diadematoid dan, akibatnya, model prediktif yang lebih kuat.


