
Abstrak
- Pohon merupakan penyerap karbon di atas permukaan tanah yang penting dalam ekosistem sabana, namun konsumsi kayu internal oleh pengurai (misalnya rayap dan mikroba) menimbulkan ketidakpastian dalam penghitungan biomassa pohon. Masih belum jelas apakah kerusakan batang internal bersifat konstan atau bervariasi di seluruh pohon, sehingga tidak pasti apakah satu sampel mencerminkan kerusakan di seluruh pohon. Lebih jauh, kerusakan total dan lokasi kerusakan kemungkinan dipengaruhi oleh tekanan kerusakan eksternal (misalnya rayap, mikroba, dan kebakaran), spesies pohon dan sifat pohon (misalnya diameter setinggi dada [DBH], kepadatan kayu) dan interaksinya.
- Kami mengambil sampel kerusakan internal di batang bawah (<2 m) pohon sabana di Queensland Utara, Australia untuk memeriksa profil vertikal jumlah kerusakan proporsional dan absolut. Kami membandingkan estimasi kerusakan dari metode sampel tunggal, dengan asumsi kerusakan konstan, dengan metode multi-sampel, dengan asumsi kerusakan variabel, untuk menguji seberapa baik satu sampel mewakili batang bawah. Kami menyelidiki apakah spesies pohon mengakumulasi kerusakan secara berbeda karena DBH, kepadatan kayu atau kerentanan terhadap tekanan kerusakan eksternal (misalnya bekas luka akibat kebakaran dan keberadaan rayap). Akhirnya, kami menguji apakah tekanan kerusakan eksternal memengaruhi spesies pohon secara berbeda dan apakah ini dimediasi oleh DBH atau kepadatan kayu.
- Jumlah absolut, tetapi bukan proporsi, kerusakan menurun dengan posisi vertikal yang lebih tinggi pada batang. Tidak ada perbedaan dalam total kerusakan batang antara metode sampel tunggal dan multi-sampel. Variasi spesifik spesies dalam kerusakan batang internal dipengaruhi oleh DBH dan kepadatan kayu. Total kerusakan paling besar terjadi pada pohon besar, terutama yang terdapat rayap eksternal. Terakhir, keberadaan rayap eksternal, tetapi bukan bekas luka bakar, berbeda di antara spesies pohon dan kemungkinan besar terjadi pada pohon besar dan padat.
- Kami menunjukkan bahwa satu sampel tunggal secara efektif menangkap kerusakan internal total pada batang bagian bawah. Meskipun spesies berbeda dalam kerusakan total, tingkat akumulasi kerusakan dengan peningkatan tinggi pada batang konsisten, yang menunjukkan hubungan umum. Dengan mengintegrasikan pengaruh tekanan kerusakan eksternal dan sifat pohon, temuan kami menggarisbawahi pentingnya mempertimbangkan elemen-elemen ini untuk memperkirakan karbon yang tersimpan dalam biomassa pohon di atas tanah secara akurat.
Baca Ringkasan Bahasa Sederhana gratis untuk artikel ini di blog Jurnal.
1. PENDAHULUAN
Sabana menutupi 20% permukaan daratan Bumi dan merupakan ekosistem yang kaya secara ekologis dengan biogeokimia yang unik (Scholes & Archer, 1997 ), namun sering diabaikan perannya dalam siklus karbon global (Dobson et al., 2022 ; Parr et al., 2014 ). Di sabana, tanaman berkayu menyimpan karbon baik saat hidup maupun mati; pohon yang hidup menyerap sekitar 0,39 Pg C setiap tahun (Grace et al., 2006 ), dan kayu yang mati menyimpan 9,6 Pg C lebih lanjut secara global (Wijas, Allison, et al., 2024 ). Dalam hal penyimpanan dan penyerapan karbon sabana, biomassa tanaman berkayu di atas tanah (AGB) menjadi perhatian khusus karena umur panjangnya di lanskap. Namun, AGB akhirnya didaur ulang kembali ke atmosfer dan tanah melalui proses dekomposisi (Zhou et al., 2007 ), di mana rayap dan api merupakan kontributor yang sangat penting dalam ekosistem sabana (Murphy et al., 2023 ; Zanne et al., 2022 ).
Sementara banyak penelitian berfokus pada penyimpanan karbon di pohon hidup (Sullivan et al., 2017 ) dan pergantian karbon melalui dekomposisi kayu mati yang tumbang (Wijas, Flores-Moreno, et al., 2024 ), kerusakan batang pohon internal merupakan komponen ekosistem sabana yang sebagian besar diabaikan dalam model siklus karbon (Calvert et al., 2024 ). Kerusakan batang internal terjadi ketika kayu di dalam pohon hidup membusuk (Janzen, 1976 ) dan telah didokumentasikan mempengaruhi hingga sepertiga AGB pohon sabana (Flores-Moreno et al., 2024 ), yang mengakibatkan ketidakpastian dalam estimasi AGB pohon (Calvert et al., 2024 ). Pembusukan rayap (Eleuterio et al., 2020 ; N’Dri et al., 2011 ; Werner & Prior, 2007 ; Yatsko et al., 2024 ) dan ‘heartrot’ mikroba (Eleuterio et al., 2020 ; Heineman et al., 2015 ; Lee et al., 1988 ; Yatsko et al., 2024 ; Zeps et al., 2017 ) berkontribusi terhadap kerusakan batang internal, namun sifatnya yang tersembunyi membuat pengukuran menjadi sulit. Untuk mengukur total kerusakan dan menentukan di mana kerusakan itu terjadi, penebangan pohon secara destruktif terbukti berguna (Calvert et al., 2024 ); namun, hal ini seringkali tidak layak secara logistik dan tidak dapat dilakukan secara ekstensif. Pengambilan sampel nondestructive lebih mudah diakses, dicapai dengan menggunakan increment corers (Heineman et al., 2015 ), resistograph drills (Flores-Moreno et al., 2024 ) dan sonic tomography (Gilbert et al., 2016 ) untuk menargetkan pengambilan sampel di batang bawah atau pada diameter setinggi dada (DBH) (Brown et al., 1995 ; Eleuterio et al., 2020 ; Flores-Moreno et al., 2024 ; Werner & Prior, 2007 ; Zeps et al., 2017 ). Namun, mungkin bermasalah untuk berasumsi bahwa satu pengukuran mewakili kerusakan di seluruh pohon (Flores-Moreno et al., 2024 ). Menangkap profil vertikal kerusakan memerlukan strategi pengambilan sampel yang direvisi, dan berpotensi untuk meningkatkan akurasi pemodelan kerusakan internal.
Bahasa Indonesia: Menuju sasaran memahami profil vertikal kerusakan batang internal, penting untuk mempertimbangkan bagaimana pelaku biotik (terutama mikroba dan rayap, Yatsko et al., 2024 ) menyebabkan kerusakan batang internal dan memanfaatkan sumber daya kayu di dalam pohon. Harapan hipotesis nol adalah bahwa pengurai internal secara acak mengonsumsi kayu di dalam pohon, membuat profil kerusakan vertikal juga acak. Dari hanya dua studi tentang profil vertikal kerusakan internal (Calvert et al., 2024 ; Yatsko et al., 2024 ), dua hipotesis alternatif non-eksklusif muncul mengenai variasi kerusakan vertikal, yang bergantung pada bagaimana kerusakan diukur. Hipotesis pola kerusakan internal pertama adalah bahwa kerusakan (diukur sebagai jumlah kayu absolut) terkonsentrasi di pangkal pohon (lihat H1 , Gambar 1a ). Di sini, ada lebih banyak kayu yang tersedia, karena diameter batang paling besar, dan dapat diakses oleh organisme kolonisasi tanah. Misalnya, rayap (agen kerusakan batang internal yang dominan di sabana, Werner & Prior, 2007 ) biasanya mencari makan di tanah dan dapat menggunakan bagian bawah batang pohon sebagai titik akses ke kayu internal atau masuk melalui akar, memusatkan kerusakan di batang bagian bawah (Yatsko et al., 2024 ). Lebih jauh lagi, jumlah kayu yang dikonsumsi oleh mikroba berkorelasi positif dengan jumlah kayu yang tersedia (Gilbert et al., 2016 ), yang dapat mengikat kerusakan pada pangkal pohon tempat batang paling besar. Umumnya, pengurai menargetkan kayu teras pohon (Perry et al., 1985 ), yang paling besar di bagian bawah pohon, berkurang dari batang ke cabang karena meruncing dan dapat sepenuhnya menghilang di cabang-cabang halus (Climent et al., 2003 ).
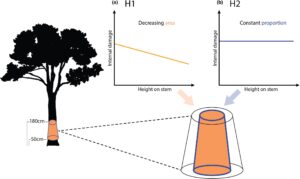
Hipotesis pola kerusakan internal kedua adalah bahwa kerusakan, sebagai proporsi, tetap konstan melalui batang bawah untuk mempertahankan struktur yang kuat. Kerusakan yang diukur sebagai proporsi kontras dengan jumlah kerusakan absolut (dari H1 ) karena tidak tergantung pada ukuran batang. Hipotesis ini sebagian besar berkaitan dengan kerusakan internal yang disebabkan rayap, karena beberapa rayap pemakan kayu hidup di pohon yang sama dengan yang mereka konsumsi (Werner & Prior, 2007 ), yang mengarah pada kebutuhan untuk mempertahankan rumah yang kuat secara struktural (lihat H2 , Gambar 1b ). Kami berharap bahwa rayap akan menggali kayu dalam proporsi yang konstan di seluruh posisi vertikal di batang bawah, sehingga bagian batang yang lebih besar di pangkal pohon akan memiliki dinding yang lebih tebal, meninggalkan struktur yang kuat untuk ditinggali (Adams et al., 1979 ; Lee et al., 2014 ). Hal ini telah dijelaskan oleh pohon rayap ‘piping’, di mana batang dilubangi seperti sedotan (Werner & Prior, 2007 ), yang kemungkinan melindungi rayap yang bersarang di dalamnya. Meskipun pengeroposan yang luas (proporsi >0,6) dapat meningkatkan kematian pohon (Werner & Prior, 2007 ), telah ditunjukkan bahwa pohon dengan kerusakan internal yang sedang dan proporsional konstan dapat mempertahankan stabilitas struktural (Mattheck et al., 1994 ), terutama jika kepadatan kayu batang luar tinggi (Larjavaara & Muller-Landau, 2010 ; Osazuwa-Peters et al., 2014 ).
Bahasa Indonesia: Untuk memperkirakan secara akurat total kerusakan batang internal dan profil vertikalnya, perlu mempertimbangkan variasi antar spesies pohon, dan jika variasi akumulasi kerusakan antar spesies didorong oleh sifat pohon yang mendasarinya, khususnya kepadatan kayu dan ukuran batang. Flores-Moreno et al. ( 2024 ) menemukan bahwa spesies dengan kepadatan kayu tinggi mengakumulasi kerusakan batang internal yang lebih besar ketika diukur melintasi gradien curah hujan tropis. Kayu padat lebih disukai oleh Coptotermes acinaciformis (Oberst et al., 2018 ), yang diketahui melubangi pohon di sabana Australia. Preferensi seperti itu dapat menyebabkan spesies pohon dengan kayu padat mengakumulasi lebih banyak kerusakan. Ukuran pohon (di sini, DBH) juga penting; beberapa penelitian menemukan bahwa pohon yang lebih besar mengalami kerusakan yang lebih sering (Heineman et al., 2015 ; Nogueira et al., 2006 ), namun yang lain menemukan hubungan yang lemah antara DBH dan total biomassa yang rusak (Flores-Moreno et al., 2024 ). Pada akhirnya, memahami bagaimana kerusakan batang internal bervariasi di berbagai spesies pohon, dan mengidentifikasi ciri-ciri pohon yang bertanggung jawab atas perbedaan ini, tetap menjadi tantangan utama dalam mengkarakterisasi kerusakan batang internal.
Di luar sifat-sifat pohon, tekanan kerusakan eksternal seperti aktivitas api dan rayap juga dapat memengaruhi akumulasi kerusakan dengan memfasilitasi akses ke kayu internal. Api penting untuk pergantian biomassa kayu di sabana (Grace et al., 2006 ; Wijas, Allison, et al., 2024 ), dan bukaan eksternal dari bekas luka api dapat memungkinkan pengurai mengakses kayu internal (N’Dri et al., 2011 ). Ada atau tidak adanya bekas luka api dapat menyebabkan perbedaan baik dalam profil vertikal maupun akumulasi kerusakan total. Sebuah studi di sabana Afrika menyimpulkan bahwa api adalah ‘agen pembuka rongga’ utama di batang pohon, yang memungkinkan masuknya rayap atau mikroba berikutnya (N’Dri et al., 2011 ). Bekas luka api basal bahkan merupakan strategi pengelolaan yang mempercepat pembentukan rongga untuk kepentingan fauna penghuni rongga (Adkins, 2006 ). Kehadiran rayap eksternal juga dapat mengindikasikan pohon yang mengalami kerusakan internal. Rayap membangun tabung lumpur, terowongan, atau ‘landasan pacu’ di bagian luar pohon, yang mengindikasikan adanya serangan hama (Li et al., 2016 ), dan koloni rayap yang dibangun di pangkal pohon dapat mengindikasikan adanya cekungan internal (Eleuterio et al., 2020 ). Oleh karena itu, keberadaan bekas luka bakar eksternal dan aktivitas rayap pada batang pohon dapat menjadi prediktor penting kerusakan batang internal.
Dalam studi ini, kami mengukur profil kerusakan internal vertikal pada batang bawah pohon sabana, yang didominasi oleh flora Myrtaceae, di Queensland Utara, Australia. Dalam sistem studi ini, api, rayap, dan mikroba semuanya berkontribusi terhadap pembusukan kayu (Clement et al., 2021 ; Law et al., 2023 ; Wijas, Flores-Moreno, et al., 2024 ) dan terdapat kerusakan batang internal yang diketahui (Flores-Moreno et al., 2024 ). Kami menjawab pertanyaan-pertanyaan berikut:
(Q1) Apakah kerusakan internal berubah dengan (a) posisi vertikal pada batang bawah di berbagai spesies atau (b) posisi vertikal pada batang bawah untuk pohon dengan kepadatan kayu dan DBH yang berbeda? Kami menyatakan kerusakan sebagai jumlah absolut untuk menguji H1 (‘kerusakan di pangkal’), dan kerusakan sebagai proporsi untuk menguji H2 (‘rumah yang kuat’).
(Q2) Apakah total biomassa batang bawah yang rusak berubah jika profil kerusakan vertikal diperhitungkan (melalui metode multi-sampel) atau tidak (melalui metode sampel tunggal)?
(Q3) Apakah spesies berbeda dalam total biomassa batang bawah yang rusak, dan, jika demikian, apakah ini disebabkan oleh sifat pohon (kepadatan kayu, ukuran batang) dan/atau tekanan kerusakan eksternal?
(Q4) Apakah tekanan kerusakan eksternal memengaruhi spesies pohon secara berbeda, dan apakah perbedaan yang terdeteksi disebabkan oleh kepadatan kayu atau ukuran batang?
2 BAHAN DAN METODE
2.1 Lokasi penelitian dan pengambilan sampel
Pengambilan sampel dilakukan di dua lokasi sabana di Queensland Utara Jauh, Australia, Station Creek (STCK, -16,61 S, 145,24 E) dan Pennyweight Station (PNW, -16,57 S, 144,92 E), yang terletak di Suaka Margasatwa Brooklyn milik Australian Wildlife Conservancy (AWC) (Gambar 2 ). PNW adalah ekosistem sabana yang lebih kering (curah hujan 812 mm per tahun) dibandingkan dengan STCK (curah hujan 1728 mm per tahun; Cheesman et al., 2018 ). Kedua lokasi mengalami musim hujan dan kemarau yang berbeda, dengan 77% curah hujan terjadi antara November dan April (Cheesman et al., 2018 ). Izin untuk kerja lapangan diberikan oleh Australian Wildlife Conservancy.
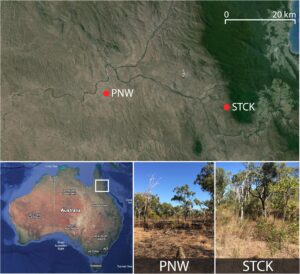
Pada tahun 2018, plot berukuran 50 × 50 m dibuat di setiap lokasi dan pengukuran DBH serta identifikasi spesies dilakukan untuk semua pohon di setiap plot. Nilai kerapatan kayu untuk setiap spesies diekstraksi dari Tree Functional Attributes and Ecological Database (Tabel S2 , Harja et al., 2019 ). PNW didominasi oleh Eucalyptus cullenii dan Melaleuca stenostachya (Myrtaceae), sedangkan STCK lebih beragam tetapi sebagian besar didominasi oleh E. cullenii dan Corymbia clarksoniana (Myrtaceae) (Tabel S1 ).
Kami memilih spesies pohon yang mencakup >85% dari biomassa plot dan dengan kejadian kerusakan batang internal yang diketahui (hingga 30% dari biomassa rusak, Flores-Moreno et al., 2024 ) untuk memeriksa profil kerusakan vertikal. Kami memilih tiga spesies Myrtaceae: E. cullenii (terjadi di kedua lokasi: STCK, 65% dari biomassa tingkat plot, PNW, 96% dari biomassa tingkat plot), M. stenostachya (PNW, 2,3% dari biomassa tingkat plot) dan C. clarksoniana (STCK, 20% dari biomassa tingkat plot, Tabel S1 ). Kami mengukur profil kerusakan vertikal pada 2 m bagian bawah batang untuk 45 pohon ( E. cullenii n = 21, C. clarksoniana n = 14 dan M. stenostachya n = 10), mengambil sampel sedikitnya 10 individu per spesies dalam empat posisi vertikal (Tabel S2 ).
2.2 Mengukur kerusakan batang internal
Kami menggunakan bor listrik IML-RESI untuk mengukur kerusakan internal (residrill; PD-500, IML, Jerman, Gambar 3a ). Residrill mengukur resistansi saat stylus berputar, menghasilkan pengukuran resistograf di mana resistansi tinggi menunjukkan kayu yang sehat dan resistansi rendah menunjukkan kayu yang rusak atau membusuk (lihat Gambar S1 ). Kami mengebor sepanjang bidang utara-selatan pada empat posisi vertikal pada setiap batang pohon (menargetkan pengambilan sampel sekitar 50 cm, 95 cm, 120 cm dan 180 cm dari pangkal pohon, Gambar 3a ). Kami mencatat DBH pohon, diameter batang di titik pengeboran, serta jarak dari titik pengeboran ke tanah.

Untuk setiap titik pengukuran, kami mengukur kerusakan sebagai jumlah absolut (melalui area penampang sampel, untuk menjawab H1 ) dan sebagai proporsi (untuk menjawab H2 ). Kami menghitung kerusakan sebagai proporsi berdasarkan area dari data keluaran residrill mengikuti metodologi dari Flores-Moreno et al. Singkatnya, ambang batas kayu sehat diklasifikasikan untuk setiap pengukuran residrill (‘jejak’, Gambar S1 ). Setiap jejak disegmentasi menggunakan fungsi sepotong-sepotong untuk meminimalkan jumlah kuadrat residual (RSS). Segmen dibandingkan dengan ambang batas kayu sehat menggunakan uji-z ekor bawah (alfa = 0,05) untuk mengukur kerusakan internal. Dari sini, kami menentukan total proporsi kerusakan berdasarkan area, yang menangkap efek nonacak dari akumulasi kerusakan pada jarak yang berbeda dari empulur (lihat Flores-Moreno et al., 2024 untuk detail pemrosesan data). Kemudian, untuk menghitung jumlah kerusakan absolut, kami mengalikan luas penampang (dihitung menggunakan luas lingkaran = π(d/2) 2 , d adalah diameter batang pada ketinggian pengukuran) dengan proporsi kerusakan berdasarkan luas untuk mendapatkan total luas kerusakan (cm 2 ).
2.3 Menghitung kerusakan pada batang utama menggunakan estimasi sampel tunggal: Kerusakan konstan
Kami memodelkan segmen batang bawah yang diambil sampelnya dengan residrill sebagai kerucut terpotong, atau frustum (Gambar 1 ). Frustum meruncing secara linear dari bawah ke atas, yang mencerminkan bentuk batang pohon yang meruncing. Kami menghitung volume frustum yang tidak rusak (yang mewakili segmen batang pohon bagian bawah) menggunakan persamaan berikut (Larsen, 2017 ):
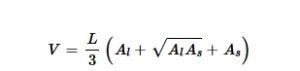
di mana V merupakan volume batang bawah, L merupakan panjang frustum, A l merupakan luas dasar frustum (titik pengambilan sampel terendah) dan A s merupakan luas atas frustum (titik pengambilan sampel tertinggi).
Untuk menghitung volume yang rusak di batang bagian bawah menggunakan estimasi kerusakan sampel tunggal, kami mengalikan volume frustum yang tidak rusak dengan proporsi kerusakan dari pengukuran residrill ~120 cm dari dasar pohon. Pengukuran 120 cm memungkinkan perbandingan dengan metode dari Flores-Moreno et al. ( 2024 ), dengan peringatan bahwa kami menggunakan pengukuran terdekat dengan DBH (1,2 m), karena desain pengambilan sampel kami tidak mencatat kerusakan internal tepat pada jarak 1,3 m dari tanah (pengukuran DBH sebenarnya). Untuk mengubah volume yang rusak menjadi biomassa yang rusak (dalam kg) di batang bagian bawah, kami mengalikan volume yang rusak dari estimasi sampel tunggal dengan nilai kepadatan kayu tingkat spesies (Tabel S2 ).
2.4 Menghitung kerusakan pada batang utama menggunakan estimasi multi-sampel: Kerusakan variabel
Untuk menghitung volume yang rusak pada batang bagian bawah menggunakan estimasi kerusakan multi-sampel, kami mendefinisikan hubungan yang meruncing untuk bagaimana kerusakan berubah antara bagian bawah dan atas batang bagian bawah. Kami memodifikasi persamaan frustum dari Larsen ( 2017) sehingga Al dan As mewakili area kerusakan dari pengukuran terendah (~50 cm) dan tertinggi (~180 cm) pada batang. Kami mengalikan volume yang rusak dengan nilai kepadatan kayu tingkat spesies untuk mengonversikannya ke biomassa (kg).
2.5 Menilai tekanan kerusakan eksternal: Kehadiran rayap dan bekas luka akibat kebakaran
Pohon diperiksa untuk mengetahui keberadaan rayap eksternal, yang didefinisikan sebagai jalur rayap pada batang (Gambar 3b ) atau gundukan yang dibangun di pangkal pohon (Gambar 3c ). Selain itu, kami mencatat ada atau tidaknya bekas luka kebakaran sedang hingga parah, yang diidentifikasi sebagai kerusakan pada kulit pangkal atau permukaan hangus yang menutupi setidaknya 25% batang bagian bawah (Gambar 3d ).
2.6 Pernyataan replikasi
| Skala inferensi | Skala penerapan faktor minat | Jumlah replikasi pada skala yang sesuai |
|---|---|---|
| Pohon individu | Spesies, kepadatan kayu | 4, 4 |
| Seluruh pohon | Spesies, kepadatan kayu | 45, 45 |
2.7 Analisis
2.7.1 Variasi kerusakan batang internal berdasarkan posisi vertikal di seluruh spesies pohon
Kami menguji bagaimana kerusakan, baik sebagai jumlah absolut ( H1 ) dan sebagai proporsi ( H2 ), berubah dengan posisi vertikal (yaitu tinggi residrill dari tanah, cm) dan jika ini bervariasi menurut spesies pohon. Untuk menentukan hubungan antara kerusakan proporsi, posisi vertikal pada batang bawah dan spesies pohon, kami menggunakan model regresi beta dengan tautan logit dari paket glmmTMB (Brooks et al., 2017 ). Kerusakan batang proporsi adalah variabel respons, posisi vertikal dan spesies pohon adalah efek tetap, dan individu pohon dan lokasi adalah efek acak. Untuk menentukan hubungan antara jumlah kerusakan absolut, posisi vertikal pada batang bawah dan spesies, kami menggunakan model efek campuran linier dari paket lmer (Bates et al., 2015 ). Luas kerusakan batang (cm 2 ) adalah respons yang ditransformasikan secara log (untuk memenuhi asumsi kenormalan) sementara posisi vertikal dan spesies pohon adalah efek tetap, dan individu pohon dan lokasi adalah efek acak. Untuk kedua model, uji perbandingan berganda post hoc berpasangan Tukey HSD digunakan untuk menentukan spesies mana yang berbeda secara signifikan.
2.7.2 Variasi kerusakan batang internal dengan posisi vertikal di seluruh DBH dan kepadatan kayu
Kami menguji apakah DBH dan kerapatan kayu memengaruhi perbedaan tingkat spesies dalam profil vertikal baik proporsi maupun jumlah absolut kerusakan batang internal. Kami menggunakan model regresi beta dengan tautan logit di mana proporsi kerusakan batang adalah variabel respons, posisi vertikal pada batang bawah, DBH, dan kerapatan kayu tingkat spesies adalah efek tetap, dan pohon individu dan lokasi sebagai efek acak. Untuk menguji efek DBH dan kerapatan kayu pada jumlah absolut kerusakan, kami menggunakan model efek campuran linier di mana area yang rusak adalah variabel respons yang ditransformasikan secara logaritmik (untuk kenormalan) dan posisi vertikal, DBH pohon, dan kerapatan kayu tingkat spesies adalah efek tetap, dengan pohon individu dan lokasi sebagai efek acak.
2.7.3 Biomassa yang rusak di batang bagian bawah: Membandingkan metode sampel tunggal dengan metode sampel jamak
Kami menggunakan uji peringkat bertanda Wilcoxon berpasangan non-parametrik (karena distribusinya tidak normal) untuk menentukan apakah estimasi sampel tunggal dari total biomassa yang rusak di batang bagian bawah berbeda dari estimasi multi-sampel.
2.7.4 Perbedaan total biomassa yang rusak berdasarkan spesies, sifat pohon dan adanya tekanan kerusakan eksternal
Kami pertama kali menguji perbedaan tingkat spesies dalam total biomassa yang rusak di batang bawah (dalam kg) menggunakan model efek campuran linier dengan total biomassa yang rusak sebagai variabel respons (ditransformasikan secara logaritma), spesies sebagai efek tetap, dan lokasi sebagai efek acak. Uji perbandingan ganda post hoc berpasangan Tukey’s HSD digunakan untuk menentukan spesies mana yang berbeda secara signifikan. Untuk menguji apakah dua sifat yang kami minati, DBH dan kepadatan kayu, menjelaskan perbedaan tingkat spesies, kami menjalankan model efek campuran linier dengan total biomassa yang rusak sebagai variabel respons (ditransformasikan secara logaritma), DBH pohon dan kepadatan kayu tingkat spesies sebagai efek tetap, dan lokasi sebagai efek acak. Kami juga menjalankan ANOVA untuk menguji perbedaan tingkat spesies dalam DBH, diikuti oleh uji post hoc berpasangan Tukey HSD. Terakhir, kami menguji apakah tekanan kerusakan eksternal berkontribusi pada biomassa yang rusak lebih besar menggunakan model efek campuran linier di mana biomassa yang rusak adalah respons (ditransformasikan secara logaritma), keberadaan rayap eksternal atau bekas luka kebakaran adalah efek tetap dan lokasi sebagai efek acak. Kami juga menguji interaksi antara keberadaan rayap eksternal dan bekas luka kebakaran.
2.7.5 Efek tekanan kerusakan eksternal pada spesies pohon, DBH dan kepadatan kayu
Kami menggunakan uji pasti Fisher untuk menentukan apakah ada atau tidaknya rayap eksternal dan bekas luka kebakaran berbeda di antara spesies. Kami menyiapkan data sebagai jumlah ada atau tidaknya rayap eksternal dan kerusakan kebakaran eksternal ke dalam tabel kontingensi terpisah. Setelah hasil uji yang signifikan, kami menggunakan uji Fisher berpasangan sebagai perbandingan post hoc dengan uji False Discovery Rate (FDR) untuk mengoreksi nilai- p untuk beberapa perbandingan. Kemudian, kami menguji peran DBH dan kepadatan kayu tingkat spesies dalam memengaruhi tekanan kerusakan eksternal (ada/tidaknya) menggunakan model binomial dengan tautan logit. Ketika DBH dan kepadatan kayu disertakan sebagai prediktor dalam model gabungan, kolinearitasnya tinggi. Karena alasan ini, kami menjalankan model binomial terpisah yang menguji setiap sifat pohon (DBH, kepadatan kayu) terhadap keberadaan rayap eksternal dan bekas luka kebakaran. Semua analisis dilakukan di R 4.4.1 (Tim Inti R, 2024 ).
3 HASIL
3.1 Variasi kerusakan batang internal berdasarkan posisi vertikal di seluruh spesies pohon
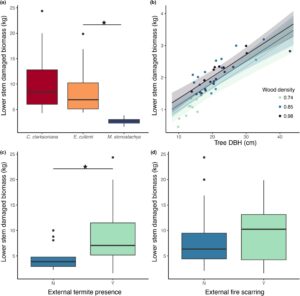
Jumlah absolut (dalam hal area) kerusakan batang internal bervariasi menurut spesies (Gambar 4a ) dengan perbedaan signifikan ditemukan antara ketiga spesies (Tabel S6 ). Jumlah absolut kerusakan internal menurun dengan posisi vertikal yang lebih tinggi pada batang ( β = −0,002, SE = 0,001, t = −3,383, Gambar 4c , Tabel S5 ). Kerusakan proporsional bervariasi menurut spesies ( C. clarksoniana > E. cullenii , M. stenostachya > E. cullenii , Gambar 4b , Tabel S4 ) tetapi tidak berubah dengan posisi vertikal pada batang utama (Gambar 4d , Tabel S3 ).

3.2 Variasi kerusakan batang internal dengan posisi vertikal di seluruh DBH dan kepadatan kayu
Luas kerusakan meningkat pada pohon dengan DBH lebih besar ( β = 0,064, SE = 0,008, t = 8,02) dan menurun pada posisi vertikal lebih tinggi pada batang ( β = −0,002, SE = 0,0005, t = −3,269, Gambar 4e , Tabel S8 ). Kepadatan kayu tidak mempengaruhi jumlah luas kerusakan (Tabel S8 ). Proporsi kerusakan tidak berubah secara signifikan berdasarkan posisi vertikal pada batang bawah (Tabel S7 ); kepadatan kayu tidak berpengaruh (Tabel S7 ), tetapi proporsi kerusakan lebih besar untuk nilai DBH lebih besar ( β = −0,017, SE = 0,007, z = −2,417, p = 0,016, Gambar 4f , Tabel S7 ).
3.3 Biomassa yang rusak di batang bagian bawah: Membandingkan metode sampel tunggal dengan metode sampel jamak
Tidak ada perbedaan dalam jumlah biomassa yang rusak ketika mengasumsikan kerusakan konstan (sampel tunggal) atau kerusakan variabel (multi-sampel) ( p = 0,14, Gambar S2 ).
3.4 Perbedaan total biomassa yang rusak berdasarkan spesies, sifat pohon dan adanya tekanan kerusakan eksternal
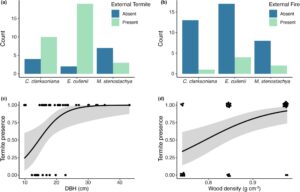
Total kerusakan internal batang bawah bervariasi menurut spesies (Tabel S10 , Gambar 5a ) dan pohon E. cullenii memiliki kerusakan yang secara signifikan lebih tinggi daripada pohon M. stenostachya ( p < 0,001) dalam analisis post hoc. Pohon yang lebih besar ( β = 0,068, SE = 0,008, t = 8,281, Tabel S11 ) dan berhutan lebat ( β = 1,516, SE = 0,565, t = 2,685, Tabel S11 ) memiliki lebih banyak biomassa total yang rusak (Gambar 5b ). Spesies berbeda secara signifikan dalam DBH (Tabel S9 ); pohon E. cullenii dan C. clarksoniana lebih besar daripada M. stenostachya (Gambar S3 ). Total biomassa yang rusak lebih tinggi pada pohon dengan kehadiran rayap eksternal ( β = −0,291, SE = 0,578, t = −0,504, Tabel S12 , Gambar 5c ), namun tidak pada jaringan parut kebakaran (Tabel S12 , Gambar 5d ).
3.5 Efek tekanan kerusakan eksternal pada spesies pohon, DBH dan kepadatan kayu
Spesies berbeda dalam kehadiran rayap eksternal ( p = 0,003), dengan perbedaan signifikan antara pohon E. cullenii dan M. stenostachya ( p = 0,01, Gambar 6a ). Tidak ada perbedaan signifikan antara spesies dan keberadaan bekas luka kebakaran eksternal ( p = 0,65, Gambar 6b ). Kehadiran rayap eksternal meningkat dengan DBH pohon ( p = 0,001, Gambar 6c ) dan kepadatan kayu ( p < 0,001, Gambar 6d ), tetapi bekas luka kebakaran eksternal pada batang tidak diprediksi oleh DBH pohon ( p = 0,79) atau kepadatan kayu ( p = 0,83).
4 DISKUSI
Dalam studi ini, kami meneliti profil vertikal kerusakan batang internal dan menguji bagaimana spesies, ciri pohon, dan tekanan kerusakan eksternal berkontribusi terhadap kerusakan batang internal. Jumlah absolut kerusakan batang internal menurun dengan posisi vertikal yang lebih tinggi pada batang, mendukung H1 , hipotesis ‘kerusakan pada pangkal’. Jumlah kerusakan bervariasi menurut spesies, yang kemungkinan besar disebabkan oleh pengaruh mendasar dari ukuran batang. Proporsi kerusakan tetap konstan di batang bawah, yang berarti bahwa di bagian batang yang lebih besar, ketebalan dinding akan lebih besar, meningkatkan stabilitas struktur. Tidak ditemukan perbedaan antara estimasi titik tunggal dan ganda dari total kerusakan, mendukung H2 , hipotesis ‘rumah yang kuat’. Total kerusakan di batang bawah bervariasi menurut spesies yang kemungkinan besar disebabkan oleh perbedaan DBH pohon dan kepadatan kayu; keberadaan rayap eksternal juga dikaitkan dengan peningkatan total biomassa yang rusak. Lebih jauh, keberadaan rayap eksternal bervariasi menurut spesies pohon dan terjadi lebih sering pada pohon yang besar dan padat. Di bawah ini kami membahas implikasi dari temuan kami pada pemodelan kerusakan batang internal dan mengusulkan bahwa memahami peran khusus rayap dapat membantu mengatasi ketidakpastian dalam estimasi karbon pohon di luar ekosistem sabana.
4.1 Jumlah kerusakan internal menurun seiring dengan meningkatnya posisi vertikal pada batang bawah
Penurunan jumlah kerusakan absolut dengan ketinggian dari pangkal pohon mendukung hipotesis ‘kerusakan di pangkal’ ( H1 ) dan menunjukkan kendala biofisik di mana akumulasi kerusakan ditentukan oleh jumlah kayu pada titik mana pun pada batang. Karena pohon meruncing, ukuran batang berkurang dari pangkal ke cabang mahkota (McTague & Weiskittel, 2021 ). Oleh karena itu, lebih banyak kayu tersedia untuk dikonsumsi pada titik pengukuran terendah dibandingkan dengan pengukuran tertinggi dari tanah. Model kami menunjukkan bahwa pohon yang lebih besar memiliki kerusakan internal yang lebih besar, yang selanjutnya mendukung bahwa ada kendala biofisik pada akumulasi kerusakan, karena pohon-pohon ini memiliki lebih banyak biomassa. Kehadiran tekanan kerusakan (rayap dan bekas luka kebakaran) sering kali bertepatan dengan tempat sumber daya kayu tertinggi di pangkal pohon (Li et al., 2016 ; N’Dri et al., 2011 ). Oleh karena itu, pangkal pohon berfungsi sebagai titik akses bagi pengurai (Adkins, 2006 ) dan area di mana kerusakan terkonsentrasi.
Hasil kami juga mendukung hipotesis ‘rumah kuat’ ( H2 ), karena kerusakan batang internal proporsional tidak berubah melalui batang bawah. Ini mendukung asumsi utama yang digunakan dalam Flores-Moreno et al. ( 2024 ) untuk memperkirakan kerusakan batang internal: Proporsi tunggal digunakan untuk mewakili kerusakan yang terjadi pada seluruh pohon, yang kemudian diterapkan untuk menskalakan biomassa yang rusak secara internal ke tingkat plot. Flores-Moreno et al. ( 2024 ) mengambil sampel kerusakan batang internal di dua lokasi sabana yang sama dan memperkirakan bahwa 18%–37% AGB pohon rusak. Studi kami mengonfirmasi bahwa estimasi ini kuat. Lubang pohon akibat rayap (Werner & Prior, 2007 ) kemungkinan bertanggung jawab atas proporsi kerusakan yang konstan, karena rayap adalah pengurai kayu utama di lokasi studi (Clement et al., 2021 ; Flores-Moreno et al., 2024 ). Rayap lubang pohon sering kali tinggal di rongga yang mereka buat (Greaves, 1962 ). Mungkin strategis bagi rayap untuk menghilangkan kayu dalam proporsi yang konstan, karena struktur pohon masih dapat mempertahankan stabilitas selama rongga tidak melebihi 70% dari radius batang (Mattheck et al., 2006 ).
Namun, tetap penting untuk mengukur profil kerusakan vertikal di lokasi lain dengan dan tanpa tekanan dekomposisi rayap yang kuat, seperti sabana lain serta ekosistem non-sabana. Berbeda dengan temuan kami, dalam ekosistem sabana dengan curah hujan yang lebih tinggi di Australia, Calvert et al. ( 2024 ) menunjukkan pola penurunan kerusakan internal. Ekosistem ini memiliki tekanan dekomposisi internal yang kuat dari rayap dan mikroba (Yatsko et al., 2024 ), dan kerusakan dimodelkan menggunakan sampel dari batang utama dan cabang tajuk. Sementara penelitian saat ini tidak menunjukkan tren penurunan yang sama, variasi dalam kekuatan tekanan dekomposisi rayap dan kondisi lingkungan (yaitu curah hujan) berpotensi menjelaskan perbedaan ini. Curah hujan mungkin penting untuk memprediksi kekuatan relatif rayap versus mikroba sebagai pengurai internal, yang dapat mengubah pola kerusakan melalui pohon. Flores-Moreno et al. ( 2024 ) menunjukkan bahwa pohon hutan hujan memiliki kerusakan batang internal yang lebih sedikit daripada pohon sabana tetapi tidak dapat mengidentifikasi bagaimana setiap proses dipengaruhi oleh pengurai yang berbeda. Namun, pada kayu mati yang tumbang diketahui bahwa pembusukan mikroba memainkan peran yang lebih besar dalam dekomposisi kayu hutan hujan tropis basah dibandingkan dengan lokasi sabana (Wijas, Flores-Moreno, dkk., 2024 ). Oleh karena itu, jika pembusukan mikroba mendominasi, profil kerusakan vertikal mungkin terlihat berbeda dari yang diamati di sini untuk pohon sabana, sehingga hipotesis ‘rumah yang kuat’ kurang berlaku.
Kerusakan batang internal tidak terbatas pada pohon tropis, karena busuk jantung mikroba juga telah dicatat di hutan beriklim sedang (Aishan et al., 2024 ; Frank et al., 2018 ; Marra et al., 2018 ; Zeps et al., 2017 ). Tingkat pembusukan dalam sistem ini cukup bervariasi: Hauck et al. ( 2023 ) melaporkan kerusakan internal <1%, Frank et al. ( 2018 ) memperkirakan tingkat pembusukan <3% dan Marra et al. ( 2018 ) memperkirakan hingga 37% karbon pohon dapat hilang karena pembusukan internal. Menariknya, Schneider et al. ( 2008 ) menunjukkan bahwa keberadaan tubuh buah pada batang pohon terkait dengan peningkatan volume kerusakan internal pohon; karena dekomposisi mikroba mendominasi ekosistem beriklim sedang (Cornwell et al., 2009 ), akan menarik untuk menguji apakah tanda-tanda eksternal mikroba tersebut secara luas memprediksi kerusakan internal di hutan pada garis lintang yang lebih tinggi. Selain itu, Zeps et al. ( 2017 ) menunjukkan bahwa kumbang penggerek kayu, selain mikroba, memainkan peran penting dalam kerusakan internal pohon beriklim sedang, jadi sementara rayap diharapkan menjadi kurang berpengaruh sebagai pengurai kayu di garis lintang yang lebih tinggi, serangga lain dapat mengisi celah ini (Ulyshen, 2016 ). Dengan dinamika dekomposisi pohon internal yang berbeda yang terjadi di ekosistem beriklim sedang, upaya untuk mengambil sampel profil kerusakan vertikal di lingkungan tersebut akan memberikan wawasan yang berguna untuk menentukan apakah pola dalam penelitian ini dapat digeneralisasikan secara luas.
4.2 Kerusakan antar spesies, ukuran pohon dan kepadatan kayu
Kami mendeteksi perbedaan dalam akumulasi kerusakan pada tingkat spesies, dengan pohon C. clarksoniana yang paling rusak dan M. stenostachya yang paling sedikit rusak. Perbedaan antar spesies paling kuat didorong oleh ukuran batang (terutama, pohon M. stenostachya adalah batang terkecil), kepadatan kayu dan keberadaan rayap eksternal; namun, pergeseran kerusakan yang luas antar spesies terjadi pada jumlah kerusakan keseluruhan (yaitu intersepsi) tetapi tidak pada akumulasi kerusakan di seluruh batang (yaitu lereng). Dari sini, kami menarik dua kesimpulan utama: (1) karakteristik kayu pada tingkat spesies merupakan kontrol penting pada seberapa banyak kayu yang dikonsumsi (yaitu menjelaskan perbedaan spesies dalam intersepsi), dan (2) antar spesies, mungkin ada hubungan umum antara akumulasi kerusakan internal dan ketinggian (yaitu konsistensi lereng antar spesies). Evaluasi yang lebih dekat terhadap sifat fisik dan kimia kayu lainnya yang dapat meningkatkan palatabilitas bagi pengurai merupakan langkah selanjutnya yang diperlukan. Misalnya, rayap mempertimbangkan faktor selain kepadatan kayu dan ukuran sumber daya dalam keputusan mencari makan, seperti kandungan kayu awal (kayu berwarna lebih terang, kurang padat), tingkat kelembapan dan bahkan seberapa baik substrat berfungsi sebagai media komunikasi (Oberst et al., 2018 ). Faktor-faktor tersebut dapat diukur menggunakan eksperimen pilihan rayap untuk menguji perbedaan tingkat spesies dalam palatabilitas kayu. Mengikuti kesimpulan kedua, kami mengusulkan bahwa jika pola kerusakan internal dapat digeneralisasikan di seluruh spesies atau lingkungan, mereka dapat dengan mudah diintegrasikan ke dalam model biomassa dengan mengasumsikan kemiringan konstan untuk mengkarakterisasi profil kerusakan vertikal. Ini akan menjadi langkah selanjutnya yang penting menuju penggabungan kerusakan batang internal ke dalam akuntansi AGB, yang belum tercapai (Calvert et al., 2024 ; Flores-Moreno et al., 2024 ). Namun, tetap penting untuk menguji apakah pola yang kita temukan di sini meluas ke tipe ekosistem dan spesies lain untuk menginformasikan model AGB secara lebih luas.
Menariknya, baik studi ini maupun Calvert et al. ( 2024 ) menemukan 20% kerusakan batang internal pada pohon Corymbia clarksoniana di dua lokasi sabana berbeda yang berjarak 600 km, meskipun ada variasi curah hujan tahunan. Konsistensi ini menunjukkan pola potensial, tetapi pengambilan sampel lebih lanjut di lokasi berbeda sangat penting untuk mengonfirmasi apakah tren tingkat spesies ini berlaku. Sejauh pengetahuan kami, belum ada studi yang membandingkan kerusakan batang internal dalam spesies di seluruh lokasi, tetapi data tersebut akan sangat berharga dalam mendokumentasikan ketahanan pola pada kerusakan batang internal khusus spesies. Memperluas upaya pengumpulan data untuk mengukur kerusakan batang internal pada tingkat spesies di seluruh rentang geografis dan ukuran dapat secara langsung dimasukkan ke dalam model AGB baru yang dibangun untuk menggabungkan efek kerusakan batang internal. Basis data komprehensif tentang jumlah kerusakan tingkat spesies dan profil vertikal akan sangat meningkatkan pemahaman kita tentang biomassa yang hilang di seluruh ekosistem.
4.3 Rumah bagi rayap di dalam pohon: Faktor penyebab dan tingkat penyebarannya secara global
Peran kepadatan kayu dan ukuran batang dalam memprediksi kerusakan internal berjalan beriringan dengan H1 dan H2 , di mana rayap menargetkan pohon yang dapat menyediakan rumah yang besar dan kuat. Misalnya, pohon C. clarksoniana mengalami kerusakan internal terbesar di batang bawah, yang tercermin dalam kayunya yang sangat padat (0,85 g cm −3 ) dan ukuran batang yang umumnya lebih besar. Kayu yang padat dan ukuran batang yang besar memberikan kekuatan pada struktur pohon (Larjavaara & Muller-Landau, 2010 ), dan karena kerusakan belum melewati ambang batas kerusakan kritis sebesar 70% (Mattheck et al., 2006 ), pohon tersebut tetap tidak terganggu secara struktural dan membentuk rongga yang terlindungi dengan baik. Di sini, kami menguji rentang kepadatan kayu yang terbatas (0,7–0,9 g cm −3 ) dan nilai DBH (10–43 cm), namun pola umumnya tetap kongruen dengan Flores-Moreno et al. ( 2024 ), yang menunjukkan kekuatan sedang dari kerapatan kayu dan DBH sebagai prediktor kerusakan internal pada rentang nilai yang lebih luas (kepadatan kayu: 0,4–0,9 g cm −3 , DBH: 10–125 cm). Sementara kerapatan kayu dan ukuran batang tampaknya menjadi prediktor yang kuat, ada berbagai sifat pohon lain yang dapat memengaruhi kerusakan batang internal (Frank et al., 2018 ). Investigasi lebih lanjut tentang peran ketebalan kulit kayu, arsitektur batang, rasio tajuk, dan kelas risiko kematian (yaitu status kesehatan) akan menjadi kontribusi yang berharga untuk memperluas pemahaman kita tentang kejadian dan kerentanan kerusakan batang internal.
Studi di sabana Australia utara telah menunjukkan bahwa pohon Myrtaceae berhutan lebat (termasuk spesies Corymbia dan Eucalyptus ) sering dilubangi oleh rayap Coptotermes (Greaves, 1962 ; Perry et al., 1985 ; Werner & Prior, 2007 ). Pohon-pohon ini mendominasi keanekaragaman spesies di sabana Australia (Crisp & Cook, 2013 ) dan mengukur tingkat kerusakan internal pada pohon-pohon ini di seluruh benua akan meningkatkan estimasi AGB pada skala lanskap (Calvert et al., 2024 ). Sementara lubang pohon telah dijelaskan secara paling luas di sabana Australia, masih menjadi pertanyaan terbuka apakah pola ini berlaku di ekosistem Australia lainnya atau bagian lain dunia tempat rayap bersarang kayu hadir. Kerusakan batang internal yang berasal dari rayap telah dilaporkan di ekosistem lain, seperti spesies Coptotermes di Amazon Brasil yang dijelaskan menjajah kayu teras pohon hidup (Eleuterio et al., 2020 ). Di Florida tenggara, AS, spesies hama invasif, C. gestroi , mengancam tajuk pohon perkotaan dengan menyebabkan kerusakan besar pada banyak spesies pohon (Chouvenc & Foley, 2018 ). Selain itu, C. formosanus di Louisiana, AS, menyerang dan merusak banyak spesies pohon perkotaan secara internal (Osbrink et al., 1999 ). Coptotermes gestroi dan C. formosanus adalah spesies yang sangat invasif dengan distribusi global (GBIF.org, 2024 ), yang memberikan dasar yang kuat untuk upaya di masa mendatang guna mengukur tingkat kerusakan batang internal secara lebih menyeluruh di luar sabana Australia.
4.4 Menghubungkan kerusakan eksternal dengan kerusakan internal
Sementara kerusakan internal tersembunyi di dalam pohon dan karenanya sulit dideteksi tanpa pengukuran invasif, kami menemukan bahwa keberadaan rayap eksternal dapat menunjukkan apa yang terjadi di bagian dalam. Satu peringatan adalah bahwa ketika menggunakan bor resistograf, tidak sepenuhnya mungkin untuk memastikan rayap sebagai penyebab langsung tanpa memeriksa kayu bagian dalam secara visual (Yatsko et al., 2024 ). Namun, hubungan antara kerusakan eksternal dan internal juga telah ditunjukkan oleh Flores-Moreno et al. ( 2024 ), di mana penulis menemukan hubungan positif yang kuat antara aktivitas rayap di kayu mati di sekitarnya dan kerusakan batang internal, terutama untuk pohon sabana yang berhutan lebat. Penelitian lain di Australia utara mengaitkan peningkatan aktivitas rayap dengan jumlah lubang pohon yang lebih banyak (Woinarski & Westaway, 2008 ; Woolley et al., 2018 ) dan menunjukkan rayap, khususnya dari genus Coptotermes , sebagai agen kerusakan internal (Greaves, 1962 ; Werner & Prior, 2007 ; Yatsko et al., 2024 ). Namun, tidak cukup hanya mengandalkan keberadaan rayap; mengetahui spesies mana yang ada sangat penting. Misalnya, Nasutitermes graveolus , spesies bersarang arboreal yang umum di area penelitian kami, membuat landasan pacu eksternal di batang pohon tetapi tidak diketahui mencari makan di mahkota atau kayu batang. Sebaliknya, N. graveolus menggunakan landasan pacu untuk perlindungan saat mereka transit untuk mencapai kayu mati di tanah (Hill, 1942 ). Sebaliknya, C. acinaciformis sering membangun gundukan di pangkal pohon dan kemudian memakan kayu pohon bagian dalam (Yatsko et al., 2024 ). Sebagian besar rayap yang kami temukan berasosiasi dengan batang yang diambil sampelnya adalah C. acinaciformis . N. graveolus hadir dalam jumlah yang lebih sedikit, dan spesies Microcerotermes juga ditemukan, yang belum banyak dikaitkan dengan dekomposisi kayu internal. Secara keseluruhan, keberadaan rayap eksternal dapat mengindikasikan adanya cekungan pohon internal yang lebih besar, tetapi perlu mempertimbangkan spesies mana yang hadir.
Kami tidak mengamati peningkatan kerusakan internal untuk pohon dengan bekas luka api basal seperti yang diharapkan; kerusakan batang internal dapat difasilitasi oleh api, yang menciptakan kondisi untuk kolonisasi biotik lebih lanjut (N’Dri et al., 2011 ). Dalam sistem kami, kerusakan api eksternal mungkin tidak menunjukkan kerusakan batang internal yang lebih besar karena tingkat keparahan api. N’Dri et al. ( 2011 ) menemukan pembukaan basal yang lebih sering pada pohon yang terpapar kebakaran dengan tingkat keparahan yang lebih tinggi, namun wilayah lokasi studi dikelola dengan kebakaran dengan tingkat keparahan rendah melalui pembakaran awal musim kemarau. Oleh karena itu, intensitas api yang lebih rendah mungkin tidak menghasilkan bekas luka basal yang sesuai yang meningkatkan invasi biotik. Lebih jauh, kerusakan api eksternal tidak bervariasi di antara spesies atau ukuran pohon, yang menunjukkan bahwa pengaruhnya tidak dimediasi oleh pemilihan sifat pohon, setidaknya untuk spesies yang dipertimbangkan di sini.
4.5 Mengintegrasikan kerusakan batang internal ke dalam estimasi karbon pohon
Estimasi akurat karbon yang diserap pohon diperlukan untuk memvalidasi solusi perubahan iklim berbasis alam dan pasar karbon hutan (Chave et al., 2014 ). Dalam studi ini di antara beberapa studi lainnya (Calvert et al., 2024 ; Flores-Moreno et al., 2024 ), kami menunjukkan bahwa kerusakan batang internal menghilangkan AGB pohon di batang bawah, dengan konsekuensi mengurangi penyimpanan karbon di atas tanah. Studi masa depan perlu memperluas pengambilan sampel kerusakan internal ke tipe dan lokasi hutan lain, terutama untuk menangkap kerusakan di batang utama tempat sebagian besar biomassa pohon total tersimpan (Ribeiro et al., 2015 ). Selain itu, sedikit yang diketahui tentang kerusakan cabang tajuk karena kesulitan pengambilan sampel; penting untuk mengkarakterisasi jumlah kerusakan dan peran mikroba dan rayap dalam struktur ini. Perhatian khusus juga harus diberikan pada pohon-pohon besar dan padat yang mengandung biomassa tinggi di lanskap untuk jangka waktu yang lama (Lutz et al., 2018 ; Stephenson et al., 2014 ). Memang, Calvert et al. ( 2024 ) menunjukkan bagaimana kerusakan yang tidak diperhitungkan pada pohon-pohon tersebut dapat mengakibatkan estimasi biomassa yang berlebihan, menunjukkan bahwa bahkan dengan penggunaan estimasi biomassa dengan akurasi tinggi (misalnya pemindaian laser terestrial), kerusakan internal tidak dapat ditangkap, yang pasti mengakibatkan estimasi biomassa yang berlebihan.
Upaya pengambilan sampel kerusakan batang internal saat ini masih sangat jarang, dan upaya untuk memperluas pengukuran ke spesies dan ekosistem baru akan menentukan generalisasi pola yang diamati dalam penelitian saat ini. Praktik berikut akan membantu mengatasi keterbatasan saat ini dalam menggabungkan kerusakan batang internal ke dalam estimasi AGB untuk hutan dunia: Jika rayap merupakan pengurai kayu yang menonjol, survei pohon untuk keberadaan rayap eksternal harus menjadi prioritas, karena ini dapat menunjukkan di mana pengeroposan internal mungkin terjadi. Karakterisasi kerusakan internal yang meluas (misalnya menggunakan residrill pada DBH) dalam sistem yang didominasi rayap lainnya akan membantu menentukan tingkat fenomena ini. Pengambilan sampel kerusakan internal yang terfokus pada pohon besar dan padat adalah kuncinya, karena ini adalah reservoir karbon yang besar. Terakhir, penelitian di masa mendatang harus menguji apakah pengeroposan pohon merupakan perilaku genus Coptotermes secara luas dan menargetkan pengambilan sampel kerusakan internal di wilayah tempat Coptotermes terjadi, baik secara alami maupun sebagai spesies invasif. Memprioritaskan upaya ini dan meningkatkan cakupan global pengambilan sampel kerusakan batang internal akan memberikan wawasan yang lebih jelas tentang dampak ekologisnya dan membantu menyempurnakan biomassa hutan dan penghitungan karbon.


