
Abstrak
- Hampir setiap aspek sistem biologi memiliki fenologi—pola aktivitas atau fungsi yang terkait dengan siklus tahunan.
- Sebagian besar penelitian fenologi terestrial berfokus pada daun, awal keluarnya daun atau penuaan. Ini adalah bentuk fenologi yang paling mudah diukur dan paling jelas. Meskipun demikian, daun bukanlah satu-satunya aspek sistem terestrial dengan dinamika musiman.
- Pola musiman baik di atas maupun di bawah tanah, lintas berbagai sifat dan organ, penting bagi siklus global air, karbon, dan nutrisi lainnya, serta interaksi antarspesies.
- Melalui plastisitas dalam alokasi sumber daya dan interaksi trofik, peristiwa fenologi tidak selalu sinkron, tetapi memiliki ketergantungan antara berbagai organ dan organisme, di atas dan di bawah tanah.
- Namun, pola lintas sifat fungsional, organ, dan organisme lebih sulit diparameterisasi dibandingkan fenologi daun dan karenanya direpresentasikan secara lebih abstrak dalam komponen vegetasi model permukaan tanah yang digunakan untuk memprediksi planet masa depan.
- Dalam perspektif ini, kami mengajukan pertanyaan berikut: Apa yang kita ketahui tentang seluruh sistem fenologi tanaman dan heterotrof tanah, dan bagaimana keduanya saling terkait? Dapatkah kita membuat prediksi tentang keterkaitan fenologi ini dan fleksibilitasnya melalui sifat, interaksinya, dan karakteristik lingkungan? Bagaimana keterkaitan ini dapat diuji dengan baik dalam kondisi lingkungan yang sebenarnya?
- Kami menantang bidang ini untuk menghasilkan data yang memungkinkan prediksi tentang penggabungan dan pemisahan fenologi seluruh sistem pada planet kita yang berubah.
Baca Ringkasan Bahasa Sederhana gratis untuk artikel ini di blog Jurnal.
1 TANGGAPAN FENOLOGI TERHADAP PERUBAHAN IKLIM
Fenologi, perkembangan musiman dan tahunan dari proses biologis siklik, adalah salah satu respons dan umpan balik yang paling terlihat dan penting terhadap perubahan iklim (Panel Antarpemerintah tentang Perubahan Iklim, 2023 ). Fenologi tanaman mendorong kenaikan dan penurunan tahunan konsentrasi karbon dioksida (CO 2 ) atmosfer, saat hutan belahan bumi utara yang luas memasuki dan keluar dari musim tanamnya (Keeling et al., 1976 ; Tucker et al., 1986 ). Saat perubahan iklim meningkatkan suhu rata-rata global, fenologi tanaman juga bergeser (Piao et al., 2019 ). Musim semi semakin awal, dan periode fotosintesis lebih panjang (Linderholm, 2006 ). Ini memiliki efek langsung pada siklus karbon (C) dan umpan balik terhadap perubahan iklim global (Cleland et al., 2007 ). Perubahan dalam fenologi tajuk terlihat jelas dari tampilan atas-bawah dalam data satelit global (Cong et al., 2012 ) dan dari bawah-atas dalam catatan jangka panjang tentang keluarnya daun dan pembungaan (Fuccillo Battle et al., 2022 ; Menzel et al., 2006 ). Namun, hubungan dengan biokimia seperti siklus C tidak dikendalikan oleh daun saja; jaringan berkayu, akar, rizosfer dan mikroorganisme tanah semuanya memainkan peran penting dalam menentukan pertukaran C daratan-atmosfer (misalnya Gambar 1 ). Dengan demikian, pemahaman yang luas tentang pola dan saling ketergantungan komponen fenologi diperlukan untuk memprediksi dan memodelkan umpan balik terestrial di masa depan terhadap perubahan iklim.
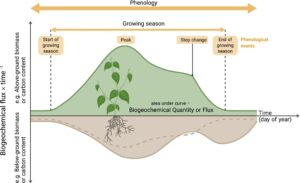
Fenologi umumnya diabstraksikan sebagai pola tahunan dalam aktivitas vegetasi—perubahan absolut dalam ukuran kumpulan komponen vegetasi utama seperti daun—dalam model Sistem Bumi (misalnya Dronova & Taddeo, 2022 ; Reed et al., 2009 ). Karena data penginderaan jauh digunakan sebagai masukan model, parsimoni mengasumsikan bahwa waktu tutupan daun atau peristiwa yang ditentukan seperti perluasan daun (misalnya Tang et al., 2016 ) menangkap aspek ekosistem fungsional fenologi di atas dan di bawah tanah. Namun, plastisitas yang sama yang memungkinkan tanaman menyesuaikan fenologi daunnya sebagai respons terhadap perubahan iklim juga memfasilitasi perubahan pada sifat tanaman lain dan alokasi sumber daya. Sekarang ada banyak penelitian yang menunjukkan fenologi pertumbuhan akar (Abramoff & Finzi, 2015 ) dan batang (Delpierre, Berveiller, et al., 2016 ) mungkin tidak berpasangan secara tepat dengan daun, dan bahkan di dalam daun, fenologi antara sifat yang berbeda tidak sepenuhnya disinkronkan (Fajardo & Siefert, 2016 ). Karena batang mengandung sebagian besar C hidup, dan akar mengendalikan sebagian besar transfer C ke penyimpanan tanah jangka panjang, representasi yang buruk dari fenologi mereka dapat menyebabkan kesalahan dalam prediksi model. Misrepresentasi ini mungkin sangat akut di bawah tanah, di mana akar berkontribusi hingga 20% hingga 33% dari serapan C vegetasi global (Jackson et al., 1996 ; McCormack et al., 2015 ) dan tergantung pada tanaman individu, hingga 95% dari total biomassa tanaman (Mokany et al., 2006 ). Lebih jauh lagi, pertukaran tanaman dengan dan di dalam tanah dapat mencakup umpan balik dua arah pada fenologi. Dengan demikian, ketergantungan heterotrofik pada autotrof (Chaparro et al., 2014 ; Jackson et al., 2019 ), ketergantungan autotrofik pada heterotrof (O’Brien et al., 2021 ; Van Nuland et al., 2021 ), dan interaksi kompetitif (Averill et al., 2014 ; Xu et al., 2015 ) semuanya dapat memengaruhi siklus dan fungsi biogeokimia (Finzi et al., 2015 ; Sobral et al., 2017 , 2023 ). Secara keseluruhan, setiap komponen ekosistem yang ingin kami prediksi di bawah perubahan global memiliki respons fenologi tingkat organisme dan/atau organ terhadap perubahan yang membuat respons tingkat sistem fungsional muncul dan berpotensi tidak terduga (Gambar 1 ).
Banyak dari komponen-komponen ini, terutama yang berada di bawah daun, bersifat samar—sulit untuk diamati atau diinterpretasikan pada skala fenologi (Albert et al., 2019 ). Misalnya, respirasi tanah merupakan campuran komponen autotrofik dan heterotrofik (Phillips et al., 2017 ) dan tertinggal dari produktivitas primer bruto (Baldocchi et al., 2006 ), sehingga sintesis berbasis penginderaan jauh mungkin tidak menangkap atau mungkin salah mengatribusikan perubahan dalam fluks ini. Namun, jika pola umum dapat diidentifikasi, hipotesis mekanistik yang mewakili komponen samar seperti akar, eksudat akar, dan mikroba tanah dapat diimplementasikan ke dalam model permukaan lahan berbasis proses dan diuji melalui pengamatan (Caldararu et al., 2023 ; Collier et al., 2018 ). Di sini, kami menyajikan bukti pola fungsional dalam komponen di bawah daun, yang dapat digunakan untuk menghasilkan hipotesis ini tentang fenologi ekosistem secara keseluruhan dan memajukan prediktabilitas sistem Bumi.
2 DI LUAR FENOLOGI DAUN HINGGA AKAR DAN BATANG
Kerangka kerja pemodelan sudah ada untuk merepresentasikan dinamika daun ekosistem (Hufkens et al., 2018 ). Data penginderaan jauh dari satelit atau jaringan kamera digital proksimal (Brown et al., 2016 ) berarti dinamika top-down diamati secara langsung. Ini termasuk menggunakan indeks properti permukaan untuk merepresentasikan fenologi daun pada skala lanskap, masuk akal karena bagian atas kanopi juga terdiri dari spesies yang paling produktif (Zhao et al., 2023 ). Variasi jangka pendek dalam kondisi lingkungan juga berarti bahwa daun bereaksi secara plastis, mengoptimalkan penyerapan C (Mäkelä et al., 2006 ; Xie et al., 2018 ) dan penginderaan jauh kanopi dapat digunakan untuk memparameterisasi model (Caldararu et al., 2014 ; MacBean et al., 2015 ; Manzoni et al., 2015 ).
Dinamika fungsi akar adalah kandidat kuat untuk peningkatan representasi fenologi atau validasi asumsi model yang ada. Namun, pengamatan fenologi akar jarang dan jarang berkelanjutan karena pengambilan sampel akar menuntut. Banyak metode penilaian akar juga membingungkan biomassa hidup dan mati jika akar tetap ada dan terurai di situ (Trumbore & Gaudinski, 2003 ). Lebih jauh, biomassa akar sering diukur pada biomassa pucuk puncak, yang secara implisit mengasumsikan sinkronisitas (Poorter et al., 2012 ) antara bagian tanaman di atas dan di bawah tanah. Sebagai organ multifungsi untuk mencari nutrisi dan penyerapan air, ada banyak alasan untuk pola fenologi yang unik pada akar. Representasi fenologi akar fungsional yang lebih baik sangat dibutuhkan untuk memperkirakan perubahan global dengan tepat, tidak hanya dalam hal siklus C tetapi juga untuk merepresentasikan hidrologi, termasuk banjir, kekeringan, dan tekanan air (Cheng et al., 2021 ; Drewniak & Gonzalez-Meler, 2017 ; Niu et al., 2020 ).
Keterkaitan antara aktivitas akar dan pucuk sulit diprediksi karena ketidakpastian dalam rentang hidup akar dan strategi perolehan sumber daya yang membatasi kemampuan peramalan pertumbuhan akar (Schwieger et al., 2019 ). Variasi mungkin disebabkan oleh plastisitas lingkungan, kondisi yang langsung menguntungkan dalam jangka pendek atau penghindaran ekstrem musiman. Namun, ada juga efek bioma dan strategi kehidupan yang jelas pada musim tumbuh akar dan fenologi hingga tingkat spesies (Abramoff & Finzi, 2015 ; Balogianni et al., 2016 ; Blume-Werry et al., 2016 ; McCormack et al., 2015 ). Misalnya, di hutan dan padang rumput, pertumbuhan akar tertinggal dari pertumbuhan pucuk (Steinaker & Wilson, 2008 ) tetapi di bawah salju Arktik dapat mempercepatnya (Blume-Werry et al., 2017 ). Inisiasi akar kemungkinan didorong oleh suhu atau asimilasi C, tetapi puncak biomassa akar mungkin didorong oleh kompetisi (Radville, Bauerle, et al., 2016 ). Beberapa penelitian kini juga menunjukkan bahwa akar juga dapat tumbuh ‘di luar musim’ di musim dingin (Nieman et al., 2018 ; Radville, McCormack, et al., 2016 ; Shane et al., 2009 ). Meskipun penelitian dilakukan pada spesies individu, penjelasan mekanistik umum dan kepentingan relatif dari faktor-faktor ini masih kurang (Erktan et al., 2018 ).
Kayu, yang menyokong daun, merupakan penyimpan C utama di atas tanah, yang menjadikan representasi akurat komponen berkayu penting untuk memprediksi serapan C di masa mendatang. Model umumnya mendasarkan pertumbuhan kayu pada paradigma ‘pertumbuhan gubal yang cukup untuk mendukung transpirasi’ yang direpresentasikan oleh gubal yang tumbuh, lapisan luar kayu tipis yang hidup, yang proporsional dengan daun. Fenologi diameter batang dapat divalidasi pada skala kasar dari dendrometer, tetapi jaringan berkayu memiliki fenologi pertumbuhannya sendiri dan variasi antara komponen internal (Lehnebach et al., 2018 ) setidaknya sebagian terpisah dari daun (Dox et al., 2022 ). Produksi meristem kambium terkait erat dengan diferensiasi menjadi xilem (Rossi et al., 2013 ), didukung oleh hubungan antara fenologi daun dan produksi pembuluh untuk mengangkut air (Guada et al., 2019 ; Lavrič et al., 2017 ). Perkembangan kayu ini mungkin lebih dikendalikan oleh sink daripada daun, tergantung pada kendala lain selain fotosintat yang baru saja difiksasi (Delpierre, Berveiller, et al., 2016 ). Memang, inisiasi pertumbuhan kayu secara umum diprediksi oleh suhu pada pohon-pohon beriklim sedang dan boreal, tetapi penghentiannya kurang terkontrol dengan baik (Delpierre, Vitasse, et al., 2016 ). Keterkaitan antara pertumbuhan batang dan respirasi batang juga tidak 1:1 (Chan et al., 2018 ) karena respirasi mungkin didorong oleh stres daripada pertumbuhan (Horna et al., 2011 ), aliran keluar massal tergantung pada aliran getah (Hölttä & Kolari, 2009 )—itu sendiri fenologis, dan pertumbuhan itu sendiri mungkin lebih terkait dengan air daripada ketersediaan C (Zweifel et al., 2006 ). Ketika struktur tegakan, alometri, demografi dan kondisi lingkungan berubah di bawah iklim masa depan, umpan balik yang tidak terduga dapat terjadi. Misalnya, perubahan iklim dapat mengubah alometri (Fu et al., 2017 ; Rudgers et al., 2019 ) dan efisiensi penggunaan air, seperti yang diamati sepanjang gradien iklim (Lines et al., 2012 ), yang mengarah pada hubungan yang berkorelasi dengan kontrol lingkungan kasar yang merespons dengan cara yang tidak dapat diprediksi.
Di luar batang, cabang sering kali membentuk seperempat dari total biomassa di atas tanah, tetapi kontrol lingkungan, ekologi, dan ontologis pada waktu dan alokasi tidak jelas (Fu et al., 2017 ; Rudgers et al., 2019 ). Dominasi apikal yang terkenal mengimbangi inisiasi tunas lateral baru, dan pemimpin pohon mendahului pertumbuhan tunas lateral. Pada vegetasi arktik, pertumbuhan cabang dan batang memiliki respons yang berbeda terhadap suhu (Buchwal et al., 2023 ), yang menunjukkan offset asinkroni atau fenologi yang tidak berpasangan dalam musim tanam yang terbatas. Fenologi pertumbuhan cabang mungkin kurang fleksibel daripada batang (Rossatto et al., 2009 ), meskipun pola percabangan keseluruhan mungkin cukup kekal (Bentley et al., 2013 ). Dalam kasus apa pun, perluasan kayu pohon mendahului peningkatan biomassa yang ketat (Cuny et al., 2015 ), dan kontrol jangka pendek pada pertumbuhan diel masih belum jelas (Steppe et al., 2015 ), jadi pemahaman tentang hubungan ini terbatas. Selain itu, beberapa tetapi tidak semua pohon secara sistematis menggugurkan cabang (cladoptosis) dan bahkan ketika ini tidak terjadi, kerusakan lingkungan dapat menyebabkan kematian cabang dalam skala waktu yang lebih pendek daripada umur pohon. Jadi, secara umum, fenologi jaringan berkayu tampaknya dapat diprediksi berdasarkan kombinasi kebiasaan ekologi tingkat spesies, kondisi lingkungan, dan fenologi daun, tetapi masih ada ketidakpastian yang cukup besar pada hubungan sebab akibat antara ketiganya dan oleh karena itu kemampuan perkiraan masa depan.
3 ASINKRONI FENOLOGI SEBAGAI STRATEGI GAYA HIDUP?
Salah satu cara untuk menggabungkan strategi fenologi tanaman utuh ke dalam prediksi yang dimodelkan adalah dengan memperluas kerangka spektrum ekonomi daun ke seluruh organisme (Reich, 2014 ). Kategori yang luas, tetapi tidak sepenuhnya berbeda, ini menyediakan mekanisme yang sederhana yang dapat dengan mudah diuji. Hipotesis yang paling jelas terletak di antara strategi gaya hidup ‘cepat’ dan ‘lambat’ (Rodrigues et al., 2021 ), yang seharusnya merespons dengan cara yang berbeda terhadap lingkungan yang bervariasi karena kemampuan yang berbeda untuk menyimpan sumber daya, tuntutan yang berbeda terhadap kelangsungan hidup jangka panjang, dan lingkungan yang berbeda itu sendiri (Gambar 2 ).
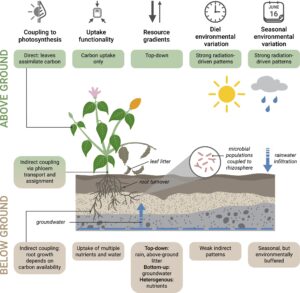
Spesies ‘cepat’ merespons variasi jangka pendek di lingkungannya, misalnya berkecambah dari bank benih dalam kondisi yang sesuai, yang menghasilkan aktivitas tahunan. Di satu sisi, kendala ukuran dari siklus hidup yang pendek berarti kemampuan terbatas untuk menyimpan sumber daya dan jalur vaskular yang pendek. Spesies ‘cepat’ tersebut dapat menghasilkan tunas dan akar secara bersamaan (Steinaker & Wilson, 2008 ). Di sisi lain, mereka juga harus merespons isyarat lingkungan jangka pendek (Abramoff & Finzi, 2015 ; Steinaker & Wilson, 2008 ) dan ketidakrataan dalam ketersediaan sumber daya. Secara umum, variasi sumber daya di bawah tanah lebih tinggi daripada sumber daya di atas tanah karena ketidakrataan spasial dan ketergantungan biologis yang kuat dari sumber daya dalam sistem tanah. Akibatnya, umur akar dan waktu pergantian mungkin perlu lebih fleksibel daripada padanan daun (Steinaker et al., 2010 ; Steinaker & Wilson, 2008 ). Dengan demikian, oportunisme fungsional dari gaya hidup ‘cepat’ mungkin justru mendorong desinkroni fenologis pada komponen tanaman di atas dan di bawah tanah.
Di ujung lain dari spektrum riwayat hidup, tanaman berumur panjang yang ‘lambat’ menyangga permintaan sumber daya melalui penyimpanan. Ini dapat menghasilkan peluang untuk desinkroni dalam ceruk tertentu. Efisiensi pertumbuhan dapat dicapai dengan mendapatkan nutrisi di luar permintaan sesaat, baik memanfaatkan periode ketersediaan tinggi, tetapi penggunaan rendah, seperti musim gugur (Muñoz et al., 1993 ; Tagliavini et al., 1998 ; Weinstein et al., 1991 ), atau dengan penyerapan nutrisi terus-menerus terlepas dari fenologi di atas tanah (Nadelhoffer et al., 1984 ). Spesies dapat menggunakan strategi pengaturan waktu sebelum atau sesudah pertumbuhan daun, atau pertumbuhan akar, yang mungkin dimodulasi oleh kondisi intra- dan inter-tahunan (McCormack & Guo, 2014 ). Beberapa cenderung mencapai puncak pertumbuhan akar di awal musim tanam ketika permintaan nutrisi tinggi (Eissenstat & Yanai, 1997 ). Periode ritmis pertumbuhan akar dan pucuk dikenal luas di Quercus (Herrmann et al., 2016 ; Reich et al., 1980 )—meskipun pola ritmis endogen terjadi pada panjang gelombang yang jauh lebih pendek daripada musim fenologis dan berpotensi dilandaskan oleh strategi sinkron atau asinkron yang lebih luas. Pasangan pertumbuhan juga dapat berubah seiring bertambahnya usia individu (Makoto et al., 2020 ), karena kombinasi ontogeni dan kendala fisik; organisme yang lebih besar dapat menyimpan lebih banyak sumber daya dan mengeksploitasi volume tanah yang lebih dalam. Jadi secara keseluruhan, tidak jelas sejauh mana strategi fenologis ini sendiri bersifat plastis. Akibatnya, sementara strategi kehidupan ini memberikan kerangka kerja yang berguna untuk merumuskan hipotesis tentang fenologi seluruh organisme tanaman, penelitian lebih lanjut diperlukan untuk menentukan sejauh mana (dan kapan) sinkronisasi di atas/bawah tanah selaras dengan strategi siklus hidup secara keseluruhan. Generalitas respons semacam itu penting untuk memprediksi perakitan komunitas tumbuhan (Weigelt et al., 2021 ) dan skalabilitas konsekuensinya terhadap perubahan global.
4 INTERAKSI BIOTIK DAN ABIOTIK DI BAWAH TANAH
Sebagian besar ketidakpastian dalam fenologi ekosistem secara keseluruhan didorong oleh plastisitas tanaman terhadap lingkungannya. Di bawah tanah, plastisitas ini memperluas kejadian lebih jauh karena penugasan C dapat digunakan untuk menumbuhkan akar, dipertukarkan dengan jamur mikoriza, atau digunakan untuk menghasilkan eksudat. Berbagai strategi di bawah tanah ini dapat menghasilkan hasil fungsional yang serupa, tetapi fenologi nyata yang berbeda dari komponen-komponennya. Dengan demikian, nasib yang bervariasi dalam sumber daya ini merupakan tantangan yang rumit dan kritis dalam memprediksi fungsi ekosistem.
4.1 Jamur mikoriza
Jamur mikoriza sangat penting untuk fungsi tanaman, memfasilitasi akses ke nutrisi dan air. Perubahan iklim mungkin telah mengubah fenologi siklus hidup mikoriza, karena pemanasan memperpanjang musim produksi sporokarp (Büntgen et al., 2012 ; Kauserud et al., 2012 ). Jamur mikoriza bersimbiosis untuk beberapa tanaman dan fakultatif atau didorong oleh lingkungan untuk yang lain (Heinemeyer et al., 2004 ). Memang, sumbu utama variasi akar lintas spesies adalah kolonisasi mikoriza dan alokasi sumber daya (Carmona et al., 2021 ). Dalam model, jamur mikoriza secara historis direpresentasikan sebagai ‘biaya’ C minor bagi inangnya. Konsep ini dapat diuraikan melampaui skalar linear melalui model ‘pasar’ pertukaran C dan nitrogen (N) dan jenis mikoriza variabel (Franklin et al., 2014 ) tetapi ini tidak divalidasi dengan baik oleh data. Memang, keseimbangan bukti menunjukkan bahwa abstraksi ke pertukaran pasar tidak realistis, dan pertukaran mikoriza lebih baik dijelaskan oleh fluks C surplus atau overflow (Bunn et al., 2024 ). Ketika permintaan C tanaman menurun selama musim tanam sedang, pergeseran terjadi ke arah pasangan jamur dalam hubungan mutualisme-parasitisme varian temporal (Neuhauser & Fargione, 2004 ). Oleh karena itu, fenologi mikoriza puncak mungkin umumnya tertunda relatif terhadap tanaman dan kekuatan hubungan ini mungkin lebih kuat dalam komunitas yang lebih dominan mikoriza. Di lintang tinggi, ini seharusnya berarti bahwa seiring akar menjadi lebih tipis dan tanaman lebih bergantung pada strategi ‘alih daya’ (Laughlin et al., 2021 ), fenologi akar mungkin lebih erat tersinkronisasi dengan fenologi mikoriza dan menyediakan cara untuk menguji hipotesis ini. Namun, kolonisasi akar mungkin merupakan kontrol semi-independen pada sinkronisasi yang bervariasi secara musiman, dengan arah perubahan yang bervariasi dalam sistem yang berbeda (Bohrer et al., 2004 ; Bouamri et al., 2014 ; Clark et al., 2009 ; Egerton-Warburton & Allen, 2000 ). Dalam beberapa kasus, mitra mikoriza dapat beroperasi saat pertumbuhan akar terbatas; misalnya, jamur mikoriza Cerrado Brasil lebih melimpah di musim kemarau daripada musim hujan (Vieira Junior et al., 2020 ). Kontrol lebih lanjut, terutama pada fenologi pembuahan, mungkin bergantung pada tanaman inang (Dickie et al., 2010 ), lingkungan atau topografi (DeMars & Boerner, 1995 ). Secara keseluruhan, interaksi antara fenologi akar dan mikoriza sulit untuk diurai tanpa pengamatan yang lebih baik (misalnya Allen & Kitajima, 2013 ; Defrenne et al., 2021) .). Hal ini khususnya tidak pasti mengingat kombinasi spesies inang dan simbion yang hampir tak terbatas. Mengembangkan metode penyaringan cepat (uji DNA berbiaya rendah atau fenotipe visual dengan analisis yang didukung AI di atas mikrofotografi yang ada untuk jamur ECM) dapat membantu menutup celah ini dan memungkinkan deskripsi fungsional jamur mikoriza.
4.2 Eksudat akar
Eksudat akar adalah cara tanaman memodifikasi lingkungan tanahnya dan memengaruhi mikrobiota lokal tanpa interaksi simbiosis langsung dengan organisme lain. Eksudat meliputi gula sederhana, asam organik, dan asam amino, tetapi saat ini tidak dapat diukur dengan mudah di lapangan (Williams & de Vries, 2020 ). Selain eksudat, pengelupasan sel selama pertumbuhan akar (Badri & Vivanco, 2009 ) berkontribusi pada ketersediaan sumber daya bagi mikroba tanah di rizosfer. Karena banyaknya mikroba yang tumbuh di sepanjang permukaan akar (Ling et al., 2022 ), substrat C ini dengan cepat diserap, menghubungkan tanaman dan populasi rizosfernya. Masih terdapat ketidakpastian yang besar dalam tingkat regulasi eksudat aktif, dan kemungkinan hal ini bervariasi di seluruh taksa dan kondisi lingkungan (Badri & Vivanco, 2009 ). Penelitian terbaru menunjukkan bahwa komposisi eksudat sebagian diatur secara aktif pada tanaman model (Pantigoso et al., 2020 ) dan mungkin memiliki pola fenologi yang berbeda di luar pertumbuhan akar yang terkait dengan metabolisme. Eksudasi juga telah terbukti memainkan peran penting dalam respons dan toleransi kekeringan tanaman (Williams & de Vries, 2020 ) dan oleh karena itu laju pelepasan dapat bervariasi sepanjang tahun dan dengan kondisi lingkungan. Dalam hal ini, perubahan eksudasi, konsentrasi, laju pelepasan dan komposisi dapat memiliki dampak penting pada siklus biogeokimia tanah karena perubahan lebih lanjut pada fungsi rizosfer (Cheng et al., 2014 ).
4.3 C-penugasan di bawah tanah
Karena C yang diangkut di bawah tanah memiliki tiga kemungkinan nasib, satu ketidakpastian utama dalam fenologi adalah bagaimana ‘peralihan’ antara akar, simbion mikoriza, dan eksudat ini bekerja (Wang et al., 2024 ). Ini termasuk apakah pola-pola ini mayoritas, berfungsi secara universal atau dapat diprediksi dan terkendali (dan karenanya dapat diprediksi karena permintaan), atau tidak. Jika tanaman berpindah antara keadaan kontrol fungsional dan surplus atas pertukaran C, maka potensi untuk menggunakan parameter partisi antara kumpulan ini terbatas. Di seluruh spesies, ada bukti terkini bahwa komposisi dan jumlah eksudat akar merupakan bagian dari strategi sifat keseluruhan (Wen et al., 2022 ; Williams et al., 2022 ). Namun, tidak diketahui apakah hubungan ini dipertahankan dalam spesies (Williams et al., 2022 ) atau apakah pola penugasan C berkorelasi dengan sifat yang lebih mudah diukur seperti diameter akar dan rasio C:N (Zadworny et al., 2015 ), yang dapat berubah seiring waktu. Ada kemungkinan juga bahwa plastisitas tanaman akan mengarah pada berbagai strategi sifat di bawah kondisi lingkungan yang berubah seperti kekeringan (Bunn et al., 2024 ) atau pemanasan (Yin et al., 2013 ). Di hutan yang didominasi oleh pohon ek merah utara dan maple merah, eksudasi akar bergantung pada spesies, tetapi tertinggi pada akhir musim panas dan gugur (Abramoff & Finzi, 2016 ), yang sesuai dengan analisis dari banyak lokasi bahwa penugasan C di bawah tanah meningkat pada akhir musim (Wolf et al., 2011 ). Secara umum, iklim yang menghangat dan musim tanam yang lebih panjang menyebabkan pergeseran dalam durasi dan besarnya masukan tanaman di bawah tanah (Järveoja et al., 2018 ). Oleh karena itu, respons pertumbuhan akar, seperti investasi tanaman dalam eksudat dan mikoriza, mungkin terkait dengan perubahan yang didorong oleh masukan ini sebanyak yang terkait dengan faktor lingkungan lainnya (Prescott, 2022 ).
4.4 Kompleks tanaman-mikroba-tanah
Bahasa Indonesia: Terlepas dari kontrol akhir masukan C rizosfer, masukan ini juga memberi makan mikroorganisme heterotrofik di zona akar (Philippot et al., 2013 ). Biomassa seluruh kumpulan mikroba tanah (baik rizosfer maupun pengurai serasah) relatif kecil (Bar-On et al., 2018 ), tetapi memainkan peran yang sangat besar dalam siklus biogeokimia dan dengan demikian memberikan umpan balik ke dalam ketersediaan nutrisi bagi tanaman dan pertumbuhan tanaman. Ketergantungan pada C autotrofik inilah mengapa sebagian besar fenologi heterotrofik tanah sering diasumsikan melacak vegetasi di atas tanah (Davidson & Holbrook, 2009 ; Hobbie, 2013 ). Secara global, pengamatan statis biomassa mikroba puncak C melacak pola lintang dalam pertumbuhan tanaman (Zhao et al., 2022 ). Namun, menerapkan logika yang sama pada variasi fenologi sebagian besar belum teruji. Memvalidasi asumsi yang masuk akal ini sangat penting untuk penskalaan. Dalam kombinasi dengan struktur tanaman di atas dan di bawah tanah serta mikoriza, mikroba tanah yang hidup bebas membuat suatu kesinambungan aktivitas biologis yang secara bersamaan merespons satu sama lain dan lingkungannya, tetapi berpotensi berbeda dalam pola fenologinya.
4.5 Mikroba tanah yang hidup bebas
Di luar rizosfer, terdapat relung metabolisme yang melimpah untuk mikroba tanah yang tidak bergantung pada kontribusi C tanaman (Malik et al., 2020 ; Yang et al., 2010 ). Misalnya, kemolitoautotrof melakukan proses-proses utama seperti nitrifikasi, prekursor biogeokimia untuk denitrifikasi dan dengan demikian produksi nitrogen oksida, tanpa memerlukan C yang difiksasi tanaman. Tidak bergantung pada masukan tanaman, fenologi kumpulan ini tidak pasti dan kemungkinan melacak pola musiman dalam suhu (Biederman et al., 2016 ) dan sumber daya yang diperlukan untuk metabolisme mereka. Studi limnologi dan berbasis laut telah memimpin bidang ini dalam melacak pola fenologi dalam komunitas mikroba (Nelson, 2009 ; Raes et al., 2022 ; Shade et al., 2007 ). Namun, sebagian besar penelitian tentang fenologi siklus biogeokimia terestrial berpusat pada tutupan vegetasi (misalnya Cooke & Weih, 2005 ; Morris et al., 2022 ; Portier et al., 2019 ).
Perbedaan skala antara mikroba dan tanaman memiliki implikasi penting bagi plastisitas fenologis mereka dan menjadikan studi aktivitas mikroba tanah yang independen dari tanaman sebagai aspek penting dalam mengatasi ketidakpastian dalam fungsi ekosistem di masa mendatang. Bukti dari ketidakselarasan spasial antara biomassa tanaman dan mikroba dan hubungannya dengan pola fungsional respirasi tanah (Zhao et al., 2022 ) menunjukkan hal ini mungkin penting dalam fenologi ekosistem. Ini mungkin karena sifat fungsional seperti perubahan efisiensi penggunaan C komunitas mikroba dapat bervariasi terlepas dari ukuran populasi total (Tao et al., 2023 ). Asinkroni dapat menyebabkan hilangnya nutrisi tanah dan C dari ekosistem karena laju dekomposisi melebihi masukan C tanaman tahunan. Lebih jauh lagi, waktu respons yang berbeda juga memengaruhi fungsi tanaman hilir, seperti penyerapan nutrisi. Misalnya, peningkatan mineralisasi N bersih dan kehilangan N dari tanah (Contosta et al., 2011 ; Miller et al., 2009 ; Morris et al., 2022 ) selama periode dormansi atau aktivitas tanaman terbatas selanjutnya meningkatkan keterbatasan N tanaman (Vitousek & Howarth, 1991 ). Secara global, pola tersebut sudah tampak dalam sifat-sifat tanah yang berkorelasi erat dengan kelimpahan mikroba; tanah dengan kandungan C rendah di daerah tropis vs. tanah dengan C tinggi di zona boreal, sesuai dengan biomassa mikroba dan fluks C heterotrofik (Gao et al., 2020 ; Zhao et al., 2022 ). Dalam skala waktu yang lebih panjang, tidak jelas apa yang terjadi lebih cepat saat lingkungan berubah: perubahan yang berarti dalam masukan C karena pergeseran fenologi tanaman atau perubahan dalam kehilangan respirasi karena peningkatan aktivitas mikroba (Melillo et al., 2002 ; tetapi lihat juga Schindlbacher et al., 2012 ). Berbeda dengan kebanyakan manipulasi yang dilakukan hingga saat ini, dampak di dunia nyata tidak akan terdistribusi secara merata sepanjang tahun atau musim tanam, sehingga eksperimen pemodelan menjadi sangat berharga dalam memahami perubahan kompleks pada populasi berumur pendek (Winnick dkk., 2020 ).
5 BAGAIMANA FENOLOGI EKOSISTEM SESUAI DENGAN SPEKTRA SIFAT?
Kami telah menyinggung bahwa ketidakpastian yang kompleks dapat dikurangi melalui kerangka kerja sifat yang disempurnakan. Spektrum sifat telah membantu menghubungkan bentuk fenotipik dengan fungsi dengan mengelompokkan sifat-sifat ke dalam kompleks dan strategi, dibatasi oleh batasan fisik (misalnya Díaz et al., 2016 ; Kong et al., 2019 ; Weigelt et al., 2021 ; Wright et al., 2004 ). Sifat-sifat selanjutnya dapat membantu menggeneralisasi pola fungsional banyak individu dan spesies, semuanya berinteraksi satu sama lain (Ellwood et al., 2010 ; Picornell et al., 2023 ; Yamaguchi et al., 2022 ) dan sumber daya yang tersedia (Bönecke et al., 2020 ; Kuczynski et al., 2017 ; Taylor et al., 1982 ), ke skala ekosistem. Kami telah menyarankan mengapa koordinasi antara bagian tanaman di atas dan di bawah tanah dapat terkait dengan strategi cepat dan lambat, tetapi apakah pola musiman dalam musim tanam yang diamati juga berhubungan dengan sifat?
Hipotesis koordinasi sifat telah dikembangkan bersamaan dengan basis data sifat tanaman (misalnya TRY; Kattge et al., 2011 , 2020 ). Sementara fenologi disertakan sebagai sifat yang berdiri sendiri dalam definisi sifat paling awal (sensu Violle et al., 2007 ), pola musiman di luar daun tidak dikenali seperti itu dalam basis data sifat. Dalam basis data TRY, hanya jenis fenologi daun kategoris (yaitu hijau abadi, gugur) dan waktu pembungaan kalender yang merupakan fitur fenologi dengan lebih dari 1000 pengamatan yang direferensikan secara geografis di antara ribuan sifat. Dalam basis data lain yang lebih terspesialisasi seperti FRED, Basis Data Ekologi Akar Halus (Iversen et al., 2017 ) atau basis data sifat jamur (misalnya Nguyen et al., 2016 ; Põlme et al., 2020 ), sifat fenologi bahkan lebih jarang atau tidak ada. Di luar perluasan basis data ini secara paksa, pengembangan teori baru tentang bagaimana fenologi sesuai dengan spektrum sifat dapat memungkinkan hubungan untuk ditargetkan dalam pengamatan baru.
Pada tingkat ekosistem, kerangka kerja properti fungsional telah diusulkan dari perspektif atas-bawah (Migliavacca et al., 2021 ) yang mencerminkan spektrum sifat daun (Gomarasca et al., 2023 ). Properti seperti produktivitas primer bruto, efisiensi penggunaan air ekosistem, dan produktivitas ekosistem neto dengan demikian menawarkan satu rute melalui teka-teki ini pada skala yang berguna untuk model. Tetapi hubungan ini sekali lagi tidak terikat waktu dan tidak selalu terkait dengan fenologi. Seperti yang telah kami uraikan, aspek fenologi daun yang diindra jarak jauh sebagian terkait dengan organ lain, tetapi sifat hubungannya sering kali tidak jelas. Misalnya, fenologi daun terkait dengan sifat kayu hidrolik saat transpirasi berpasangan dengan anatomi xilem (de Lima et al., 2021 ). Hubungan ini terkadang bersifat searah, misalnya ukuran pembuluh berkorelasi baik dengan daun yang keluar (Savage & Chuine, 2021 ), tetapi terkadang juga lebih melingkar—misalnya pertumbuhan daun dan ekspresi sifat selama waktu fenologis bergantung pada pasokan C dan perkembangan arsitektur daun yang sama (Fajardo & Siefert, 2016 ). Secara keseluruhan, masalahnya mirip dengan tingkat spesies dalam menskalakannya ke dinamika musim tanam; strategi oportunistik mendikte respons jangka pendek, tetapi tidak selalu kebiasaan jangka panjang dan pola musim tanam fenologis.
Komponen penting dari inisiasi musim tanam yang nyata dan akibatnya panjang keseluruhan pada skala temporal dan emergen adalah mekanisme dormansi kuncup pada pohon beriklim sedang dan boreal, dan mekanisme dormansi biji. Kontrol dormansi kuncup yang lebih kuat merupakan indikator toleransi stres, dan posisi relatif pada spektrum sifat yang lambat. Dormansi biji yang lebih kuat terkait dengan strategi persistensi dan gaya hidup yang lebih lambat (Zhang et al., 2022 ), tetapi juga oportunisme pada tanaman tahunan ketika kondisi pemutusan dormansi merupakan relung temporal seperti pasca kebakaran (Pausas et al., 2022 ) dan dapat berhubungan dengan interaksi antara variasi fotoperiode musiman dan lingkungan (Norton et al., 2016 ). Upaya yang cukup besar (di luar cakupan tinjauan ini) telah dilakukan untuk memahami mekanisme dan bagaimana ini terkait dengan kontrol lingkungan dalam memengaruhi tanggal endodormansi (Chuine et al., 2016 ). Dalam mengintegrasikan kontrol yang kompleks, yang dapat berupa arah atau intensitas berdasarkan pemahaman sifat, sifat baru seperti ketahanan terhadap dingin mungkin penting (North & Kovaleski, 2024 ). Dalam elaborasi ini, menetapkan fitur dormansi mana yang umum di seluruh organ dan mana yang unik juga penting; ada bukti tidak adanya dormansi dalam pertumbuhan akar pada spesies ‘lambat’ (Malyshev et al., 2023 ), meskipun kehati-hatian harus dilakukan karena periode pertumbuhan dan penghentian juga terkait dengan pasokan sumber daya serta pemutusan dan permulaan dormansi.
Di bawah tanah, sifat-sifat akar terbagi ke dalam beberapa sumbu lain, spektrum nonlinier lain yang memuat kedua sumbu gaya hidup lambat-cepat dan trade-off antara strategi penggunaan C ‘lakukan sendiri’ dan ‘alihdayakan’ (Bergmann et al., 2020 ; Carmona et al., 2021 ; Wang et al., 2024 ; Weigelt et al., 2021 ). Karena trade-off ini tidak ditemukan di atas tanah, dan C sebagai sumber daya dibagi antara metabolisme, pertumbuhan, eksudat, dan transfer ke mitra mikoriza, pola musiman sistem akar tidak selalu terkait baik dengan transfer C kotor di bawah tanah. Bahkan jika tingkat asimilasi C tanaman dijelaskan dari kombinasi sifat-sifat daun yang terwujud, fenologi akar yang prediktif akan terbatas karena pergantian. Secara global, tingkat pergantian akar lebih baik diprediksi oleh suhu daripada curah hujan (Gill & Jackson, 2000 ), tetapi pola musiman sudah sulit karena ambiguitas penampilan yang disebutkan di atas (Trumbore & Gaudinski, 2003 ). Pengamatan fenologi yang dapat diskalakan dan diintegrasikan di bawah kurva juga terbatas karena hanya sebagian kecil studi fenologi akar yang benar-benar kuantitatif (Radville, McCormack, et al., 2016 ). Sebuah meta-analisis baru-baru ini dari pengamatan fenofase di atas dan di bawah tanah menemukan bahwa pemanasan tidak mengubah fenofase di bawah tanah, tidak seperti kemajuan di atas tanah pada tanaman herba, tetapi pada tanaman berkayu fenologi di bawah tanah lebih sensitif (Liu et al., 2021 ). Pada skala ekosistem, efeknya tentu saja juga bergantung pada interaksi antar spesies. Secara umum dalam sistem musiman, puncak pergantian akar halus yang nyata terjadi di akhir musim tanam dalam sistem musiman (Abramoff & Finzi, 2015 ) yang berkorelasi dengan waktu puncak berbuah dari jamur ECM yang bergantung pada akar, sementara pengurai kayu mencapai puncaknya lebih awal (Kauserud et al., 2012 ; Sato et al., 2012 ). Pola-pola ini dapat diartikan sebagai bukti surplus C (Prescott, 2022 ; Trugman & Anderegg, 2025 ) yang mendorong aktivitas mikoriza. Pergeseran puncak dalam pergantian akar berpotensi memicu aktivitas mikroba, dan terkait lebih jauh dengan ketersediaan N tanah dalam beberapa sistem tahunan (Morris et al., 2019 , 2022 ) dan pola produksi serasah secara umum lebih penting untuk masukan C ke dalam tanah daripada eksudasi. Meskipun kami belum membahas ciri-ciri mikroba dalam tinjauan ini sejauh ini, ada upaya untuk mengukur respons heterotrofik tanah terhadap perubahan global dari perspektif berbasis sifat (Malik et al., 2020 ). Upaya ini akan dibantu dengan semakin banyaknya data tentang genom mikroba dan fungsi gen (Karaoz & Brodie, 2022 ), respons bakteri terhadap stresor lingkungan (Cébron et al., 2021 ), dan informasi berbasis sifat pada plasmid tanah (Dunivin et al., 2019 ). Akan tetapi, banyak dari upaya ini yang mensintesis data yang tersedia saat ini, dan pengukuran baru dari berbagai ekosistem tanah pada skala waktu yang lebih besar diperlukan untuk menangkap pola fenologi.
Variabilitas tinggi dalam fenologi di antara spesies individu dan komponen ekosistem dapat mengaburkan sinyal fenologi ekosistem secara keseluruhan yang mungkin selaras dengan spektrum sifat. Tumbuhan individu menunjukkan mekanisme dormansi dan pola fenologi yang berbeda, yang dipengaruhi oleh faktor lingkungan dan strategi kehidupan yang saling berinteraksi. Misalnya, sifat daun dan akar tertentu, termasuk kebiasaan hijau abadi dan toleransi kekeringan, dapat memperpanjang musim tanam dan memisahkan peristiwa fenologi satu sama lain. Di bawah tanah, pergantian akar dan aktivitas mikroba mengikuti pola yang kompleks dan bervariasi secara musiman yang semakin memperumit lanskap. Akibatnya, sementara pendekatan berbasis sifat dapat memprediksi perilaku fenologi tertentu, sifat respons spesies individu yang rumit dan tidak sinkron dapat menutupi pola fenologi ekosistem yang lebih luas.
6 ASPEK LEBIH LANJUT DARI FENOLOGI EKOSISTEM SELURUHNYA
6.1 Interaksi spesies
Tautan antara spesies yang berinteraksi satu sama lain dan lingkungannya memiliki kompleksitas tambahan yang mengarah pada efek yang muncul (Gambar 3 ). Fenologi pada skala ekosistem adalah hasil dari relung individu itu sendiri yang terdiri dari komponen bawaan (Donohue, 2005 ) dan plastik (Diamond et al., 2011 ; Inouye, 2008 ), bertambah melalui komunitas yang berinteraksi (Cleland et al., 2006 ; Elzinga et al., 2007 ; Miller-Rushing & Primack, 2008 ; Schwartz & Hanes, 2010 ). Interaksi spesies, seperti penyerbukan (Kharouba et al., 2018 ) dan partisi relung (Archibald & Scholes, 2007 ; Singh & Kushwaha, 2005 ), semuanya berulang kali ditemukan untuk merespons perubahan global. Struktur fisik (berpotensi dipengaruhi oleh interaksi ini) selanjutnya memainkan peran dalam tanggal fenologi melalui regulasi iklim mikro (Wu et al., 2024 ). Interaksi lain, seperti invasi, juga bergantung pada kebiasaan fenologi (Wolkovich & Cleland, 2011 ). Efek ini dapat berinteraksi dengan strategi kehidupan; misalnya, spesies yang lebih kecil mengalami lebih banyak kompetisi (Aarssen & Keogh, 2002 ). Kompetisi juga dapat mendorong respons pada beberapa organ di luar periode ‘optimal’. Dalam sistem sabana yang kaya spesies di Iberia, banyak pertumbuhan akar terjadi di musim dingin (Vieira Junior et al., 2020 ) ketika ketersediaan nutrisi tinggi tetapi masukan fotosintesis rendah, sementara vegetasi Arktik terbagi antara yang didominasi alang-alang dengan pertumbuhan akar awal musim dan komunitas tanaman berkayu yang tumbuh di bawah tanah di akhir musim (Sloan et al., 2016 ). Efek kompetisi tersebut dapat mengikuti fenologi mikroba tanah, dengan dampak hilir pada waktu ketersediaan nutrisi. Interaksi spesies dan pengaturan waktunya dalam komunitas mikroba tanah mungkin, bagaimanapun, memiliki sedikit efek pada fungsi, karena redundansi fungsional (Yang et al., 2010 ). Tetapi perubahan dalam komunitas mikroba sangat spesifik konteks (Williams & de Vries, 2020 ), dan perubahan global dapat mempengaruhi beberapa anggota kunci lebih dari yang lain. Misalnya, sensitivitas suhu metanogenesis umumnya lebih tinggi daripada oksidasi metana (Einola et al., 2007 ; Fan et al., 2022 ), dengan konsekuensi untuk anggaran metana ekosistem. Eksperimen yang ditargetkan dengan kuantifikasi anggota komunitas mikroba dan penanda fungsional (misalnya melalui metagenomik dan transkriptomik) diperlukan untuk menentukan apakah pergeseran dalam fenologi mikroba akan memodifikasi biogeokimia yang tampaknya digerakkan oleh tanaman.
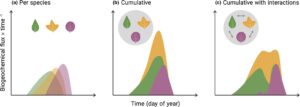
Jadi secara keseluruhan, tidak jelas apakah interaksi spesies atau komposisi komunitas dan kekayaan spesies memengaruhi fungsi fenologi ‘area-under-the-curve’ bersih dan sejauh mana larangan fenologi dari umpan balik lingkungan atau fisiologis terganggu oleh keanekaragaman (Gambar 3 ). Kehati-hatian harus diambil ketika mengekstrapolasi antara individu dan komunitas; pola fenologi yang diamati dari segala jenis pengukuran agregatif (penginderaan jarak jauh, hitungan akar minirhizotron) biasanya berada pada tingkat komunitas tetapi memiliki pengaruh yang bervariasi dari individu dengan adaptasi fenologi spesifik tambahan. Model berbasis kohort tempat dinamika komunitas dapat disimulasikan dapat memberikan rute untuk memahami pola-pola ini.
6.2 Ketersediaan nutrisi
Kondisi lingkungan memengaruhi fenologi, tetapi fenologi juga memengaruhi kondisi lingkungan lokal. Ketersediaan nutrisi sebagian dikendalikan oleh fenologi tanaman karena tanaman menggunakan sumber daya dan mendaur ulang nutrisi tersebut di dalam dirinya sendiri (Nord & Lynch, 2009 ). Desinkroni antara aktivitas tanaman dan ketersediaan nutrisi dapat meningkat karena perubahan iklim (Twining et al., 2022 ) dan respons cepat di bawah tanah dalam sistem perolehan sumber daya. Interaksi dengan fenologi kemungkinan berbeda antara N dan P karena nitrat sangat mobil, tetapi ortofosfat tidak. Nitrogen mencapai puncak ketersediaannya menjelang akhir proses dekomposisi, yang dapat meningkatkan aktivitas akar, khususnya dalam sistem yang terbatas N dan sangat musiman secara bersamaan (Radville, Bauerle, et al., 2016 ). Di sisi lain, defisiensi P menginduksi berbagai respons morfologi akar untuk memperoleh P (termasuk produksi eksudat) tetapi tidak boleh mengikuti siklus musiman yang kuat, karena sementara P dilepaskan dari pergantian, ia hanya dihilangkan melalui penyerapan. Respons fenologi komponen kriptik ini mungkin sangat penting dalam kondisi sumber daya rendah dan dengan alasan kuat untuk tidak sinkron dengan daun (Gambar 2 ). Interaksi antara nutrisi dan air juga memodulasi fenologi sistemik; misalnya, sistem Mediterania yang lebih subur menggunakan air lebih cepat, mempercepat fenologi (Luo et al., 2020 ), dan mengubah distribusi akar dan biomassa dibandingkan dengan perlakuan tanpa pupuk (Nair et al., 2019 ). Ini menunjukkan bahwa kehati-hatian harus dilakukan dalam sistem musiman untuk tidak salah menafsirkan respons alokasi C tanaman, seperti pergeseran dari biomassa di atas tanah ke di bawah tanah di bawah air dan keterbatasan N (Feng et al., 2023 ; Hermans et al., 2006 ; Puglielli et al., 2021 ), menjadi indikasi fungsi. Namun, bukti lapangan tentang bagaimana lingkungan memoderasi penugasan di atas/di bawah tanah pada skala musiman masih sangat terbatas. Secara umum, kami menekankan bahwa fenologi daun yang tampak kemungkinan besar terkait dengan alokasi fungsional ini dan respons mencari makan di bawah tanah dan harus dipertimbangkan bersama-sama, jika memungkinkan.
7 MENINGKATKAN MODEL DENGAN DATA BARU
Sementara banyak kemungkinan pengaturan eksperimental dapat menguji hubungan fenologis, urgensi perubahan global berarti ini perlu direpresentasikan sebaik mungkin dalam model segera. Secara keseluruhan, peran plastisitas, strategi gaya hidup, kompleks sifat (sensu Kramer-Walter et al., 2016 ; Reich, 2014 ), dan saling ketergantungan di antara mereka secara implisit direpresentasikan dalam model dalam cara mereka merepresentasikan ketergantungan lingkungan (Gambar 2 ) dalam strukturnya. Di wilayah tanpa arsip alami (seperti lingkaran pertumbuhan) atau pendanaan untuk upaya eksperimental besar yang menyediakan data berkelanjutan, model memungkinkan proses ini divalidasi dengan pengukuran yang lebih jarang (Caldararu et al., 2022 ).
Pendekatan seperti rasio tetap (misalnya gubal: luas daun, rasio akar: pucuk) sangat hemat dan diterapkan dalam model modern (misalnya Thum et al., 2019 ). Bahkan ketika lebih rumit, representasi didasarkan pada asumsi. Misalnya, akar telah direpresentasikan melalui rasio 1:1 dengan daun, kemudian dengan skalar linear (misalnya Lawrence et al., 2019 ; Wiltshire et al., 2021 ) dan/atau pemicu lingkungan seperti kekeringan atau embun beku yang mencegah atau memungkinkan pertumbuhan akar (misalnya Warren et al., 2015 ) yang secara umum mengikuti pengamatan (Alvarez-Uria & Körner, 2007 ; Kummerow et al., 1990 ) tetapi mungkin bukan mekanisme biologis. Dalam kebanyakan kasus, alokasi sumber daya di bawah tanah dibatasi oleh hubungan default di atas tanah–di bawah tanah dalam model. Bahkan ketika ‘alokasi dinamis’ dijelaskan, atau prinsip optimalitas diterapkan, dalam praktiknya sering kali berarti hanya mengubah skalar linier antara alokasi C daun dan akar halus (misalnya Thum et al., 2019 ). Optimalitas, yang semakin banyak digunakan dalam model, adalah penugasan sumber daya untuk memenuhi faktor yang paling membatasi pada fungsi biaya, biasanya menggunakan produktivitas primer bruto sebagai proksi untuk kebugaran. Pola fenologi, koordinasi sifat pada skala organ dan organisme, dan bahkan mungkin sifat emergen skala ekosistem dapat diprediksi dengan teori optimalitas (Caldararu et al., 2014 ; Harrison et al., 2021 ; Manzoni et al., 2023 ). Representasi ini juga menghasilkan keluaran frekuensi tinggi dan deret waktu dinamis yang meyakinkan, tetapi proses yang mendasarinya masih perlu divalidasi oleh data pada skala yang sesuai; ada kemungkinan untuk menjadi benar karena alasan yang salah. Jumlah situs yang relatif kecil dengan solusi berteknologi tinggi untuk mengukur fenologi bawah tanah seperti dendrometer akar (Alday et al., 2020 ), minirhizotron otomatis (Defrenne et al., 2021 ; Nair et al., 2022 ), dan pemindai terkubur (Nakahata, 2022 ) dapat memberikan peluang untuk menguji asumsi ini dengan cara yang sama seperti eksperimen FACE digunakan untuk memvalidasi respons model terhadap konsentrasi CO2 di masa mendatang .
Lebih jauh lagi, untuk representasi lengkap siklus C dalam ruang fenologi, fluks C bawah tanah yang terkait dengan akar—termasuk eksudat dan mikoriza—harus disertakan. Sementara beberapa estimasi total penugasan C bawah tanah dan dengan demikian data validasi untuk model telah dibuat melalui estimasi dari masukan di atas tanah (Gower et al., 1996 ; Raich & Nadelhoffer, 1989 ), penanganan kotak hitam ini tidak fleksibel. Bahkan jika penugasan di bawah tanah dibagi di antara beberapa kumpulan di bawah tanah, itu tidak berpasangan dengan dinamika di atas tanah. Saat ini ada upaya besar lebih lanjut untuk meningkatkan komponen ekosistem yang tersembunyi, seperti heterotrof, di ruang angkasa melalui meta-analisis statistik atau pembelajaran mesin (Gao et al., 2022 ; Huang et al., 2021 ; Poggio et al., 2021 ). Tanpa pemahaman dan validasi mekanistik, kelemahan seperti saling ketergantungan keluaran sintetis (misalnya Meyer & Pebesma, 2022 ) hanya akan menjadi lebih parah jika diperluas ke waktu fenologis, lintas spesies dan tipe ekosistem yang berbeda karena kelangkaan umum pengamatan fenologis.
Kumpulan non-akar ini sangat jarang terwakili dalam model dan baru-baru ini terwakili baik secara implisit maupun eksplisit (misalnya Lawrence et al., 2019 ; Wang et al., 2022 ). Ketika teori menjadi lebih maju, kerangka kerja yang jelas (misalnya paradigma nilai tukar atau limpahan) dapat memungkinkan hal ini dielaborasi lebih lanjut. Kesulitan yang sangat akut adalah perbedaan antara peran absolut dan fungsional; C yang dimiliki dalam jamur mikoriza adalah biomassa hifa, tetapi tingkat penyerapan N diubah oleh tingkat simbiosis. Tidak jelas sejauh mana kolonisasi mikoriza pada akar, panjang hifa dan tingkat penyerapan fungsional berkorelasi satu sama lain sebagai sifat yang dapat diprediksi (Van Der Heijden & Scheublin, 2007 ), yang diperlukan untuk penskalaan melalui kerangka kerja sifat dari waktu ke waktu. Dalam kasus yang serupa, tetapi lebih ekstrem daripada hubungan akar: pucuk, representasi eksplisit akan memungkinkan pengukuran lapangan yang melelahkan untuk memvalidasi model. Satu rute bagi jamur untuk ini adalah dengan menilai waktu produksi sporokarp. Meskipun tidak mewakili semua jamur, pengukuran ini secara umum lebih mudah dan dapat menjadi tolok ukur yang masuk akal untuk menilai kinerja fenologi model.
8 TANTANGAN LEBIH LANJUT
Salah satu dari banyak tantangan untuk merepresentasikan fenologi tanah–tanaman secara keseluruhan dengan cara yang sederhana dalam model adalah bias spasial dalam data; singkatnya, sebagian besar pemahaman kita berasal dari daerah beriklim sedang dengan musim tanam yang berbeda, kondisi yang menguntungkan untuk kerja lapangan, dan bias historis terhadap observasi sejarah alam dan ketersediaan dana modern (Martin et al., 2012 ; Nuñez & Amano, 2021 ). Hal ini membatasi kemampuan kita untuk memodelkan tipe ekosistem lain dengan kuat dan kehati-hatian harus diberikan agar aturan umum tidak diterapkan secara keliru sebagai faktor spasial yang disamarkan. Misalnya, meskipun daerah tropis basah sering dianggap selalu hijau, hal ini berbeda dari fenomena tingkat ekosistem yang sama di garis lintang tinggi karena ada pola fenologi spesies dan daun yang dapat diamati pada skala sub-tahunan (Reich et al., 2004 ; van Schaik et al., 1993 ) dengan efek biogeokimia langsung (Alves et al., 2018 ; Girardin et al., 2016 ). Pola tahunan yang disimpulkan dari lingkaran pertumbuhan juga kurang dalam pengaturan ini karena pola pertumbuhan sub-tahunan, pertumbuhan radial sektoral yang berbeda, lingkaran palsu, pemisahan yang tidak jelas antara lingkaran pertumbuhan individu, atau tidak ada lingkaran sama sekali (Silva et al., 2019 ). Selain itu, di wilayah semi-kering, yang keduanya berkembang (Tercek et al., 2023 ) dan penting secara global (Ahlstrom et al., 2015 ), kelembapan membatasi musim tanam, yang menyebabkan periode aktivitas maksimum didorong oleh air daripada suhu. Dalam beberapa kasus, fenologi bahkan bimodal, dengan periode pembatasan yang berbeda di musim panas dan musim dingin karena faktor yang berbeda (Valeriano et al., 2023 ). Keterbatasan ini dapat berdampak kuat pada cara kerja fenologi. Misalnya, ketika air menjadi sumber daya pembatas utama dalam setahun, kayu akan tumbuh dominan pada musim hujan (Angoboy Ilondea et al., 2021 ) dan ketika air melimpah setiap tahun, panjang hari dan suhu dapat menentukan pertumbuhan kayu (Oliveira et al., 2009 ).
Kapasitas aklimatisasi organisme (beradaptasi dengan lingkungan yang berubah selama masa hidupnya) merupakan tantangan lain untuk memahami fenologi seluruh ekosistem. Kemampuan tanaman untuk beraklimatisasi dalam jangka panjang dapat mengubah respons terhadap fenologi, seperti untuk menghemat air (misalnya Luo et al., 2020 ; Zelikova et al., 2015 ) atau membatasi degradasi senyawa fotosintesis (Reich et al., 1991 , 2004 ). Secara keseluruhan, aklimatisasi ini dapat mengubah pola fenologi total (Marqués et al., 2023 ), terutama pada tanaman semusim berganti daun (Gallinat et al., 2015 ), tetapi dinamika tersebut saat ini tidak dimasukkan ke dalam kerangka kerja pemodelan (Fisher & Koven, 2020 ). Secara potensial, kemampuan daun untuk beraklimatisasi berkorelasi dengan sifat-sifat lain yang telah dipelajari lebih baik, sedangkan kemampuan aklimatisasi sendiri merupakan sifat yang berpotensi dapat diukur dan dapat dimasukkan ke dalam basis data sifat.
Akhirnya, kami sengaja membatasi pembahasan heterotrof pada populasi mikroba, tetapi hewan tentu juga memiliki respons fenologis terhadap perubahan iklim (Cohen et al., 2018 ) dan mungkin menjadi penggerak utama fungsi ekosistem (Bauer & Hoye, 2014 ), termasuk siklus C (Schmitz et al., 2018 ). Fenologi mereka tidak selalu samar, tetapi juga tidak terwakili dalam model. Mempelajari fenologi fungsional dari tingkat trofik motil yang lebih tinggi ini merupakan tantangan yang jauh lebih sulit daripada yang pernah kami coba di sini. Kemungkinan besar dalam beberapa kasus serangga pemakan floem dan penggugur daun mungkin jauh lebih penting untuk fungsi ekosistem skala luas daripada yang lain karena keterkaitannya yang kuat dengan tanaman dan populasi yang dinamis. Mengidentifikasi dan merepresentasikan umpan balik tersebut pada tingkat abstraksi yang tepat merupakan tantangan yang menarik dan kritis di masa mendatang.
9 KESIMPULAN
Kami telah menyoroti bahwa fenologi bukan hanya daun, dan pemahaman yang komprehensif tentang fenologi ekosistem diperlukan untuk memahami kopling penuh dari beberapa siklus unsur dan prediktabilitas fungsi ekosistem di masa depan yang tidak pasti. Eksperimentalis sering mengabaikan kumpulan kriptik ini, dan pemodel menyederhanakannya, yang dapat menyebabkan pemahaman yang bias dengan implikasi yang berpotensi besar bagi banyak pertanyaan perubahan global. Sementara kami menyusun kerangka kerja yang luas dan belum teruji untuk memahami fenologi, kami telah berusaha mengembangkan ide-ide yang dapat masuk ke dalam desain eksperimental. Secara umum, karena fenologi merupakan respons utama terhadap perubahan global dan tantangan untuk dipelajari dalam siklus penelitian reguler, kumpulan data jangka panjang sangat penting. Kumpulan data sains warga adalah alat yang berguna untuk fenologi daun (misalnya Kosmala et al., 2016 ; Purdy et al., 2023 ) tetapi akan jauh lebih sulit untuk dimanfaatkan untuk kumpulan ekosistem yang kriptik, tetapi tetap mendasar. Untungnya, data terbuka dengan cepat menjadi standar emas untuk ilmu sistem Bumi (Reichman et al., 2011 ), dan pengukuran yang saat ini langka dapat, dan kami percaya akan, dibuat tersedia dengan cepat bagi komunitas untuk mem-bootstrap pengetahuan penting ini. Untuk kumpulan yang kurang karismatik seperti akar dan mikroba, jaringan terdistribusi mungkin merupakan cara yang paling efisien untuk meningkatkan cakupan dengan upaya minimum (misalnya Worldwide Hydrobiogeochemistry Observation Network for Dynamic River Systems, WHONDRS, Garayburu-Caruso et al., 2021 ). Untuk mengatasi respons fenologi, pengambilan sampel ulang temporal juga harus dilakukan di lokasi individual dan ditargetkan untuk mengurangi ketidakpastian tentang kumpulan kriptik tanpa prior yang kuat berdasarkan kemudahan pengambilan sampel yang kurang kriptik. Oleh karena itu, kami meminta mereka yang memiliki sumber daya untuk melakukan pengukuran fenologi berulang dari kumpulan kriptik ini jika memungkinkan.


