
Abstrak
- Memahami dampak penuh dari penekanan spesies yang diperkenalkan dan invasif pada komunitas ekologi memerlukan eksperimen penekanan jangka panjang di mana spesies asli dengan riwayat hidup yang berbeda dapat mencapai keseimbangan populasi baru. Respons yang didokumentasikan dalam studi jangka pendek cenderung bersifat sementara.
- Studi ini mengevaluasi kelimpahan dan struktur ukuran 11 spesies ikan pesisir selama 20 tahun sebagai respons terhadap penangkapan ikan smallmouth bass tahunan di seluruh danau, predator non-asli yang bersaing dengan dan memakan spesies asli.
- Banyak perubahan dalam kelimpahan relatif spesies terlihat jelas dari perbandingan komposisi komunitas sebelum, segera setelah, dan 20 tahun setelah eksperimen penekanan penyerbu. Dinamika jangka panjang diklasifikasikan sebagai menunjukkan perubahan mendadak, perubahan bertahap, atau tidak ada perubahan yang terlihat, berdasarkan analisis titik perubahan.
- Kelimpahan relatif sebagian besar anggota komunitas ikan meningkat pada tahun setelah dimulainya penghilangan ikan bass tetapi menjadi lebih bervariasi setelahnya. Setelah peningkatan awal, sebagian besar spesies mengalami penurunan kelimpahan, dengan beberapa menurun hingga mendekati atau di bawah tingkat sebelum percobaan dan yang lainnya tetap berada di atas tingkat sebelum penghilangan.
- Banyak spesies menunjukkan penurunan awal ukuran tubuh setelah ikan bass disingkirkan, yang menunjukkan peningkatan kelimpahan ikan muda, tetapi pola jangka panjang bervariasi di antara spesies.
- Fluktuasi populasi yang terus-menerus selama penelitian ini menunjukkan bahwa arah, tempo, dan stabilitas respons populasi terhadap penghilangan penyerang dominan sering kali bersifat sementara. Perubahan dalam kelimpahan populasi dan ukuran tubuh selama eksperimen 20 tahun kami tidak berkorelasi antar spesies, yang menunjukkan bahwa komunitas ikan memerlukan banyak generasi untuk mencapai keadaan stabil baru setelah hilangnya atau penekanan predator dominan.
- Sintesis dan aplikasi. Upaya pemulihan komunitas ikan jarang diikuti oleh evaluasi jangka panjang, oleh karena itu pemahaman saat ini tentang respons terhadap penghilangan spesies yang diperkenalkan dan invasif mungkin sebagian besar didasarkan pada dinamika sementara. Sebaliknya, kelimpahan populasi dan struktur ukuran harus didokumentasikan setidaknya untuk beberapa generasi spesies yang berumur paling panjang. Wawasan tentang hasil pengelolaan dengan menekan atau menghilangkan predator yang diperkenalkan dan invasif memerlukan penghitungan jeda waktu demografis karena interaksi spesies menjadi terstruktur ulang dalam jangka waktu yang lama.
1. PENDAHULUAN
Pengenalan dan invasi ikan telah memengaruhi ekosistem perairan di seluruh dunia melalui dampak ekologis dan ekonominya. Penyebaran ikan non-asli yang luas (Dawson et al., 2017 ) dimungkinkan oleh perikanan rekreasi dan pelepasan ikan dari akuarium, serta penyebaran setelah pembentukan awal (Coucherousset & Olden, 2011 ; Gozlan et al., 2010 ). Ikan yang diperkenalkan dan invasif mengganggu ekosistem melalui pengecualian kompetitif fauna asli (Bohn et al., 2008 ; Coulter et al., 2019 ; Sampson et al., 2009 ) dan pemangsaan terhadap mangsa asli (Green et al., 2012 ; Knapp et al., 2001 ; Tiberti et al., 2014 ). Perubahan-perubahan ini mengubah struktur jaring makanan (misalnya komposisi komunitas; Carlisle & Hawkins, 1998 ; Flecker & Townsend, 1994 ) dan fungsi (misalnya perubahan dalam struktur trofik dan aliran energi; Detmer & Lewis, 2019 ; Vander Zanden et al., 1999 ), dan perubahan-perubahan ini dapat meluas melewati batas-batas ekosistem (Detmer et al., 2017 ; Epanchin et al., 2010 ; Knight et al., 2005 ).
Dampak upaya pemulihan ekosistem akuatik biasanya dievaluasi dengan perbandingan biner antara periode pra dan pascaintervensi. Namun, mengkarakterisasi tempo dan pola respons dalam periode pascaintervensi penting untuk menilai pengelolaan berkelanjutan, terutama mengingat respons variabel oleh spesies yang diperkenalkan dan invasif (Abrams, 2009 ; Zipkin et al., 2009 ). Meskipun penelitian jangka panjang jarang dilakukan, kepentingannya terdokumentasi dengan baik (Detmer et al., 2024 ; Likens, 1989 ; Magnuson, 1990 ) dan tinjauan komprehensif oleh Prior et al. ( 2018 ) menunjukkan bahwa respons ekologis terhadap upaya penghapusan spesies yang diperkenalkan dan invasif jarang dinilai selama lebih dari 5 tahun. Mengkarakterisasi lintasan respons jangka panjang penting untuk memahami berapa lama ekosistem perlu mencapai keadaan baru setelah eliminasi atau penekanan penyerbu, dan bagaimana keadaan itu dibandingkan dengan kondisi ketika penyerbu dominan. Respons ekosistem awal dapat terjadi dengan cepat karena perbedaan dalam pemilihan mangsa oleh predator asli versus predator yang diperkenalkan (misalnya Lepak et al., 2006 ). Sebaliknya, jeda waktu dalam respons terhadap upaya pengelolaan atau pemindahan yang intensif dapat terjadi karena mekanisme kompensasi yang tidak terduga (Zipkin et al., 2008 ). Ikan dapat menunjukkan waktu generasi yang panjang, hingga beberapa dekade (Tabel 1 ). Oleh karena itu, sebagian besar spesies hanya mengalami beberapa generasi selama durasi penilaian yang khas setelah upaya pengendalian invasi. Yang lebih memperumit pemahaman kita tentang penekanan spesies yang diperkenalkan dan invasif adalah potensi evolusi yang cepat (Zarri et al. dalam revisi ) atau adaptasi perilaku (Detmer et al. dalam tinjauan ). Lebih jauh lagi, invasi sekunder dapat memperumit respons, karena perubahan dalam kelimpahan satu spesies invasif dapat meningkatkan kondisi untuk spesies lain yang diperkenalkan atau invasif (O’Loughlin & Green, 2017 ). Oleh karena itu, untuk mencapai kondisi stabil baru setelah penyingkiran yang terkelola atau bahkan pemberantasan spesies yang diperkenalkan atau invasif bisa memakan waktu puluhan tahun, tergantung pada komposisi komunitas, seberapa dalam fungsi ekosistem diubah oleh spesies non-asli, dan respons terhadap penekanan spesies target.
| Nama umum | (Genus dan spesies) | Usia saat dewasa | Sumber | ||
|---|---|---|---|---|---|
| Rata-rata (rentang) a | Sumber | Usia maksimal (tahun) | |||
| Ikan Trout Danau | Salvelinus namaycush | 11.3 (7,0–20,0) | Basis Ikan ( 2023 ) | 50 | Kekuasaan ( 1978 ) |
| Pengisap putih | Catostomus komersoni | 2.0 | Trippel dan Harvey ( 1991 ) | 17 | Bahasa Indonesia : Beamish ( 2011 ) |
| Ikan trout sungai | Bunga Salvelinus fontinalis | 2.3 (1,5–3,5) | Basis Ikan ( 2023 ) | 24 | Reimers ( 1979 ) |
| Ikan putih bulat | Prosopium silindris | 4.1 (3.0–5.5) | Basis Ikan ( 2023 ) | 16 | Furnis ( 1974 ) |
| Ikan bass mulut kecil | Mikropterus dolomieu | 5.5 | Basis Ikan ( 2023 ) | 15 | Scott dan Crossman ( 1973 ) |
| Ikan lele coklat | Burung pipit Ameiurus nebulosus | 2,5 (2,5–2,5) | Basis Ikan ( 2023 ) | 18 | Freyhof dan Kottelat ( 2007 ) |
| Ikan mas biji labu | Lepomis gibbosus | 2.2 (2.0–2.5) | Basis Ikan ( 2023 ) | 12 | Hubbel ( 1966 ) |
| Ikan pari biasa | Luxilus cornutus | 1.0 | Basis Ikan ( 2023 ) | 6 | Altman dan Dittmer ( 1962 ) |
| Ikan chub sungai | Semotilus atromaculatus | 3.2 (2,5–3,5) | Basis Ikan ( 2023 ) | 8 | Pelukan ( 1996 ) |
| Ikan smelt pelangi | Osmerus mordax | 3.0 (2.0–6.0) | Basis Ikan ( 2023 ) | 7 | Bailey ( 1964 ) |
| Lumpur tengah | Batasan Umbra | 1.0 | Robinson dan kawan-kawan ( 2010 ) | 9 | Mahon ( 1984 ) |
| Patung berlendir | Cottus cognatus | 3.0 (2.0–4.0) | Basis Ikan ( 2023 ) | 7 | Craig dan Wells ( 1976 ) |
Rata -rata dan rentang usia saat dewasa dihitung dari studi yang disertakan dalam Fishbase. Studi ini mencakup usia rata-rata saat ikan dari populasi tertentu menjadi dewasa untuk pertama kalinya. Usia tersebut dihitung dari panjang saat pertama kali dewasa menggunakan kebalikan dari fungsi pertumbuhan von Bertlanffey: tm = t0 – ln (1 – Lm / Linf ) /K ( https://www.fishbase.se/manual/key%20facts.htm ).
Selain ketidakpastian tentang berapa lama respons ekosistem terhadap penekanan spesies yang diperkenalkan atau invasif harus dipantau, tidak ada konsensus tentang metrik apa yang harus dilacak. Penting untuk menentukan tujuan berdasarkan pemahaman tentang bagaimana dampak ekologis berskala dengan ukuran populasi spesies target (Green & Grosholz, 2021 ), karena upaya pengendalian yang dikelola dan non-kimia jarang memberantas spesies target. Program pengendalian spesies yang diperkenalkan dan invasif sering kali berfokus pada demografi spesies target mereka, tetapi berfokus pada hasil untuk populasi spesies asli sering kali merupakan target yang lebih tepat (Lambert et al., 2023 ). Namun, bahkan setelah populasi fokus diidentifikasi, peneliti juga harus memutuskan apakah akan fokus pada metrik perilaku, demografi, komunitas atau ekosistem dari respons sistem.
Bahasa Indonesia: Untuk memperjelas domain waktu yang dibutuhkan untuk memahami respons ekosistem terhadap penekanan spesies yang diperkenalkan dan invasif, kami mengkarakterisasi perubahan dalam kelimpahan dan struktur ukuran ikan litoral selama upaya dua dekade untuk menekan populasi ikan predator yang diperkenalkan (smallmouth bass, Micropterus dolomieu ) yang mendominasi biomassa komunitas. Setelah upaya awal pemberantasan penuh, tujuan percobaan ini selama 20 tahun terakhir adalah pemberantasan fungsional smallmouth untuk meningkatkan ukuran ikan olahraga asli dan komunitas ikan asli. Evaluasi awal menunjukkan bahwa smallmouth bass mempertahankan ukuran populasi yang besar melalui perekrutan yang terlalu kompensasi, tetapi struktur usia dan ukuran bergeser secara dramatis untuk memasukkan lebih banyak juvenil dan lebih sedikit dewasa besar (Zipkin et al., 2008 ). Respons demografis ikan litoral lainnya juga cepat; spesies ini mencapai 4 hingga 90 kali kelimpahan sebelum pemindahan dalam 2 tahun pertama setelah pemindahan smallmouth bass dimulai (Weidel et al., 2007 ). Smallmouth bass menunjukkan pergeseran demografis yang jelas dari individu berukuran sedang ke populasi yang didominasi oleh individu yang lebih kecil, sebagian besar masih muda (Weidel et al., 2007 ). Peningkatan jangka pendek pada ikan bertubuh kecil memungkinkan predator puncak asli, ikan trout danau ( Salvelinus namaycush ), menjadi lebih pemakan ikan (Lepak et al., 2006 ). Semua pengamatan tersebut didasarkan pada data tidak lebih dari 6 tahun setelah dimulainya percobaan penghilangan bass, durasi yang mirip dengan sebagian besar studi penekanan penyerbu (Prior et al., 2018 ). Tim kami telah melanjutkan percobaan pers ini dengan menghilangkan bass empat kali setahun dan mencatat tangkapan per unit upaya semua spesies dalam komunitas ikan ini, dengan demikian memberikan kesempatan untuk menguji apakah respons demografis awal telah bertahan, dan perubahan dalam struktur ukuran bass predator juga telah mendorong perubahan ukuran pada spesies lain.
Black bass ditetapkan secara global sebagai pengganggu jaring makanan komunitas asli, terutama melalui konsumsi ikan asli (Cerrilla et al., 2022 ; Long & Seguy, 2024 ; Van Der Walt et al., 2016 ; Vander Zanden et al., 1999 ). Di sini, kami mengukur perubahan dalam kelimpahan dan struktur ukuran ikan asli dan non-asli untuk menentukan apakah dan kapan anggota komunitas ikan Little Moose Lake mencapai keadaan stabil baru selama dua dekade penekanan smallmouth bass, spesies black bass. Pertama, kami membandingkan respons jangka pendek yang diamati oleh Weidel et al. ( 2007 ) dengan hasil jangka panjang, kemudian mengkarakterisasi perubahan antara keadaan pra-eksperimen dan tahun pertama setelah penekanan eksperimental; tahun pertama penghilangan smallmouth bass dianggap sebagai bagian dari keadaan pra-eksperimen. Kedua, kami menggunakan analisis titik perubahan untuk mengidentifikasi titik waktu ketika pergeseran distribusi yang tiba-tiba dalam kelimpahan relatif terjadi untuk setiap spesies. Jika tidak ada titik perubahan yang dapat diidentifikasi, kami menggunakan regresi untuk mengevaluasi apakah terjadi pergeseran kelimpahan secara bertahap. Kami mengantisipasi bahwa penekanan ikan bass mulut kecil akan mengakibatkan peningkatan kelimpahan dan pengurangan ukuran tubuh anggota komunitas ikan lainnya setelah penekanan eksperimental ikan bass, yang mungkin berdampak terutama pada tahap muda tahun spesies lain melalui predasi dan kompetisi. Penilaian tahunan telah memberikan peluang unik untuk mengevaluasi respons yang tertinggal di antara spesies yang timbul baik dari riwayat hidup yang lambat dan/atau bervariasi (Tabel 1 ) atau pengaturan ulang beberapa jalur interaksi yang memerlukan waktu untuk menstabilkan kembali sebagai respons terhadap eksperimen pers.
2 BAHAN DAN METODE
2.1 Lokasi dan latar belakang penelitian
Studi ini dilakukan di Little Moose Lake (43.692242° Lintang, −74.925711° Bujur) di New York, Amerika Serikat. Danau ini memiliki luas permukaan 271 ha, kedalaman maksimum 44 m, dan dialiri oleh lima anak sungai orde pertama. Tingkat gangguan antropogenik di danau ini rendah untuk ukurannya; danau ini terletak di cagar alam pribadi yang tidak dapat diakses oleh publik dan memiliki sekitar 45 bangunan rumah perahu pribadi di sepanjang garis pantainya yang berisi satu atau lebih jalur perahu kayu. Kualitas air tidak berubah secara dramatis sejak diperkenalkannya ikan bass mulut kecil, dengan model linier nilai Juli dari tahun 1998 hingga 2020 yang menunjukkan tidak ada perubahan pada kedalaman Secchi Juli ( F (1,21) = 0,06; p = 0,81) atau total fosfor ( F (1,21) = 0,98, p = 0,33). Meskipun ada sedikit peningkatan karbon organik terlarut dari sekitar 2 mg/L menjadi sekitar 3,5 mg/L ( F (1,21) = 18,37, p < 0,001), seperti yang umum terjadi di Adirondacks selama periode ini (Jane et al., 2024 ). Perubahan suhu air permukaan pada bulan Juli dan tanggal keluarnya es selama periode tersebut tidak berubah (suhu air Juli F (1,47) = 2,823, p = 0,10; tanggal keluarnya es F (1,20) = 1,021, p = 0,24). Little Moose Lake memiliki tekanan memancing yang rendah relatif terhadap ukurannya untuk perikanan ikan trout (Cox & Walters, 2002 ) karena telah melakukan rata-rata 195 perjalanan per tahun sejak 1998, dengan upaya terbesar dilakukan pada ikan trout danau diikuti oleh ikan bass mulut kecil. Namun, untuk kedua spesies, sangat sedikit individu yang dipelihara setiap tahun: 40 untuk ikan trout danau dan 9 untuk ikan bass mulut kecil. Ikan bass mulut kecil diperkenalkan selama tahun 1940-an dan dianggap ‘melimpah’ pada tahun 1950-an (Weidel et al., 2007 ). Di wilayah tersebut, ikan bass mulut kecil diperkenalkan sebagai ikan olahraga. Ada rencana pengelolaan yang menjelaskan pengenalan ikan bass mulut kecil di Little Moose Lake, meskipun tidak jelas apakah itu dilakukan secara tidak sengaja sebagai ikan umpan atau sengaja untuk menjebak ikan olahraga lain. Spesies non-asli lainnya telah diperkenalkan dan mempertahankan populasi yang persisten selama penelitian ini, termasuk ikan smelt pelangi ( Osmerus mordax ), ikan salmon Atlantik yang terkurung daratan ( Salmo salar ) dan ikan mudminnow tengah ( Umbra limi ). Selain beberapa spesies asli, Little Moose Lake juga memiliki nilai konservasi yang unik karena berisi ikan putih bulat ( Prosopium cylindraceum ) (Departemen Konservasi Lingkungan Negara Bagian New York, 2023 ), yang dianggap terancam punah di negara bagian New York. Bahasa Indonesia: Penangkapan ikan listrik dengan perahu digunakan untuk memperkirakan kelimpahan relatif (tangkapan per unit usaha, selanjutnya CPUE) dan struktur ukuran komunitas ikan pesisir (untuk tinjauan metodologis, lihat Weidel et al., 2007 ) setiap tahun selama musim semi (Mei–Juni; Lampiran S1 dalam Informasi Pendukung). Karena sifat jangka panjang dari penelitian ini dan variabilitas kondisi antar-tahunan selama pengambilan sampel, kami mengevaluasi dan mengamati tidak ada pengaruh kondisi pada komunitas yang diambil sampelnya (Lampiran S1 ). Semua pengambilan sampel dilakukan sesuai dengan IACUC di bawah Cornell 2008–0009 dan di bawah izin Lisensi untuk Mengumpulkan Ilmiah dari Thomas Detmer, LCP 2700. Sebelum tahun 2003, jumlah lokasi pengambilan sampel (9 hingga 30) dan persentase garis pantai (91 ± 3%) yang dicakup bervariasi dari tahun ke tahun. Kami mengecualikan data dari tahun 2002 karena lokasi pengambilan sampel pada tahun itu hanya mencakup 37% dari garis pantai. Setelah tahun 2003, jumlah lokasi meningkat menjadi 32, dan 100% garis pantai disurvei. Analisis awal menunjukkan bahwa pengecualian tahun-tahun tambahan dengan upaya pengambilan sampel di bawah rata-rata hanya menyebabkan sedikit perubahan dalam kesimpulan tentang respons ikan pumpkinseed dan bullhead (lihat Lampiran S2 ); oleh karena itu, kami menganalisis data dari semua tahun kecuali tahun 2002. Untuk menghilangkan ikan bass mulut kecil dan mengkarakterisasi seluruh komunitas ikan pesisir, kami menggunakan perahu standar yang dialiri listrik untuk menarik ikan kira-kira setiap 30 meter di sepanjang garis pantai. Upaya dicatat sebagai waktu perahu tersebut secara aktif menyetrum ikan. Kami mengumpulkan setiap ikan yang diamati, dan semua individu kecuali bass diidentifikasi, diukur, dan dilepaskan dengan segera (lihat Marcy-Quay et al., 2019 untuk informasi lebih lanjut). Bass di-eutanasia, dibekukan, dan kemudian diukur panjang dan beratnya. Rincian lebih lanjut tentang sejarah percobaan penekanan bass kami dirangkum dalam Weidel et al. ( 2007 ), Zipkin et al. ( 2008 ) dan Lepak et al. ( 2006 ). Survei elektrofishing musim semi pada tahun 1998, 1999, dan 2000 digunakan untuk menilai keadaan pra-penekanan komunitas ikan. Penghapusan smallmouth bass dimulai pada musim semi 2000, tetapi data dari tahun itu mencerminkan kondisi pra-penghapusan karena komunitas belum menanggapi pemusnahan bass. Penekanan smallmouth bass bervariasi antar tahun dalam upaya dan waktu (Lampiran S2–S3 ); penangkapan ikan dengan alat elektro pada tahun 2000 dan 2001 berlangsung dalam jangka waktu yang lebih panjang setiap tahunnya dibandingkan pada tahun 2003 dan tahun-tahun berikutnya karena kami berusaha keras untuk menghilangkan sebanyak mungkin ikan bass mulut kecil (Lampiran S3 ). Sejak tahun 2003 dan seterusnya, penangkapan ikan dengan alat elektro untuk menekan ikan bass mulut kecil di sekitar seluruh tepi danau telah terjadi empat kali setahun: satu malam musim semi (pertengahan Mei), dua siang musim semi (akhir Mei hingga pertengahan Juni), dan satu malam musim gugur (pertengahan September). Semua ikan bass dimatikan dengan cara eutanasia selama setiap putaran upaya, tetapi data pada semua spesies lainnya dikumpulkan hanya selama survei malam hari. Di sini, kami menganalisis data komunitas ikan hanya dari survei elektrofishing malam musim semi karena itu adalah catatan terlengkap kami dari tahun 1998 hingga 2020. Perbedaan dalam waktu dan intensitas elektrofishing musim semi di antara tahun-tahun dapat menciptakan bias dalam rangkaian waktu kami, jadi kami menilai efek tanggal pengambilan sampel pada tangkapan semua spesies, dan apakah pemindahan smallmouth bass menciptakan efek penipisan dari peristiwa pemindahan berturut-turut (lihat Lampiran S4 ) di mana pengambilan sampel smallmouth bass pada peristiwa kedua akan dipengaruhi oleh peristiwa pengambilan sampel awal di mana ikan dipindahkan. Kami menemukan sedikit efek waktu musiman; hanya pumpkinseeds pada tahun 2000 dan slimy sculpins pada tahun 2001 yang menunjukkan efek negatif yang lemah dari hari dalam setahun. Kami membatasi analisis data smallmouth bass pada tahun 2000 hingga hari dalam setahun hari 135–153 untuk menghindari efek penipisan, tetapi tidak ada bukti penipisan pada tahun 2001. Secara keseluruhan, perbedaan tanggal pengambilan sampel tidak secara substansial mempengaruhi kelimpahan ikan yang diamati; Oleh karena itu, kami menggunakan semua data yang tersedia dari survei penangkapan ikan listrik musim semi pada tahun 1998–2001 dan 2003–2019.
2.2 Analisis statistik
Kami menggunakan tangkapan per unit usaha (CPUE) untuk mewakili kelimpahan relatif untuk 11 dari 16 spesies di Little Moose Lake. Kelimpahan relatif, diukur sebagai jumlah individu yang ditangkap per waktu untuk setiap kunjungan ke setiap lokasi, harus menjadi metrik yang andal untuk mendeteksi perubahan kelimpahan dari waktu ke waktu dan di seluruh lokasi, meskipun tidak dapat digunakan untuk memperkirakan ukuran populasi total. Ketika ada beberapa putaran penangkapan ikan listrik musim semi di lokasi yang sama dalam satu tahun, kami merata-ratakan hasilnya untuk memperkirakan kelimpahan relatif setiap spesies. Kami mengecualikan ikan dace perut merah utara ( Chrosomus eos ) dan ikan dace hidung hitam ( Rhinichthys atratulus ) karena mereka muncul pada kelimpahan yang sangat rendah hanya dalam satu dan enam dari 20 tahun, masing-masing. Untuk fokus pada ikan yang memberikan kontribusi yang berarti pada jaring makanan dan tidak dimanipulasi secara artifisial, kami mengecualikan tiga salmonid yang populasinya ditambah dengan penebaran ikan selama percobaan: ikan trout pelangi ( Oncorhynchus mykiss ; 1–6 juvenil per ha selama 13 dari 20 tahun), ikan salmon Atlantik yang terkurung daratan ( Salmo salar ; 1–6 juvenil per ha selama 13 dari 20 tahun) dan ikan trout sungai ( Salvelinus fontinalis ; 3–5 juvenil per ha pada tahun 2000 dan 2001, dan 20–28 per ha sebelum tahun 1998). Ikan trout pelangi dan ikan trout sungai menunjukkan rekrutmen yang buruk untuk memancing, yang menunjukkan kontribusi yang terbatas pada jaring makanan. Akibatnya, penebaran mereka menjadi jarang dan akhirnya dihentikan karena biaya tinggi dan pengembalian rendah. Penebaran ikan salmon Atlantik yang terkurung daratan terus berlanjut karena minat yang tinggi dalam pengembangan perikanan salmon trofi. Meskipun demikian, kecil kemungkinan mereka berdampak berarti pada jaring makanan, karena perbandingan ikan bersudut sebelum dan sesudah pemindahan menunjukkan bahwa rasio bersudut ikan yang ditebar (ikan trout sungai, ikan trout pelangi, dan ikan salmon Atlantik daratan) terhadap ikan trout danau tidak berubah ( uji -t dua sisi yang tidak berpasangan dengan varians yang tidak sama, semua p > 0,05, tidak dipublikasikan), dan oleh karena itu kemungkinan besar lebih erat kaitannya dengan upaya penebaran daripada respons terhadap dinamika jaring makanan.
Untuk mengkaji pergeseran antara kelimpahan relatif sebelum pemindahan dan segera setelah pemindahan, kami melakukan uji peringkat bertanda Wilcoxon non-parametrik untuk tahun 2000 dan 2001; uji peringkat bertanda Wilcoxon digunakan sebagai uji non-parametrik yang kuat untuk mengkarakterisasi perubahan dalam arah dan besarnya perbedaan yang dapat diterapkan pada semua spesies, termasuk yang tidak memenuhi asumsi kenormalan. Analisis titik perubahan juga digunakan untuk mengidentifikasi perubahan dalam kelimpahan relatif selama penelitian ini (Matteson & James, 2014 ). Analisis titik perubahan non-parametrik multivariat digunakan untuk mengidentifikasi waktu dan arah perubahan dalam kelimpahan relatif spesies dari waktu ke waktu (R versi 4.2.1, paket ‘ecp’). Pendekatan ini memperlakukan setiap lokasi sebagai replikasi dalam danau setiap tahun, kemudian mengidentifikasi pergeseran dalam mean atau varians distribusi untuk mengidentifikasi kelompok tahun dengan distribusi yang sama. Kami mendefinisikan titik perubahan sebagai pergeseran yang berlangsung setidaknya 2 tahun. Perubahan tahun ke tahun yang tidak berlanjut selama setidaknya 2 tahun tidak dihitung sebagai titik perubahan. Respons cepat antara tahun pertama dan tahun setelah penekanan, yang mungkin tidak bertahan cukup lama untuk diidentifikasi sebagai titik perubahan, dinilai dengan uji peringkat bertanda Wilcoxon untuk tahun pertama dan setelah penekanan, dengan mengakui bahwa pergeseran tersebut tidak mewakili status distribusi baru yang dicirikan oleh rata-rata atau variasi baru. Analisis titik perubahan dijalankan lebih dari 1000 pengulangan, dengan pengaturan alfa = 1. Alfa adalah metrik divergensi yang mewakili divergensi dalam distribusi antara vektor independen yang membentuk urutan nilai yang dipisahkan oleh setiap titik perubahan. Alfa = 1 memungkinkan estimasi titik perubahan untuk mengidentifikasi perubahan dalam rata-rata dan distribusi (Matteson & James, 2014 ) . Untuk memahami pengaruh potensial perubahan jumlah situs seiring waktu, kami juga menjalankan analisis kelimpahan relatif dengan alfa 2, di mana hanya pergeseran rata-rata yang diidentifikasi, tetapi tidak ada perubahan substantif yang tampak (Lampiran S2 ).
Untuk spesies yang CPUE-nya tidak dapat dikarakterisasi dengan titik perubahan, kami mengkarakterisasi tren jangka panjang dalam kelimpahan relatif dengan memasang model regresi binomial negatif yang digelembungkan nol menggunakan paket R ‘pscl’. Struktur model ini dipilih karena prevalensi hitungan nol dalam data, karena spesies tertentu jarang atau tidak pernah muncul di lokasi tertentu di seberang danau. Binomial negatif sering digunakan untuk memodelkan data hitungan, sementara binomial negatif yang digelembungkan nol digunakan untuk menangani penyebaran berlebih dalam data (Vaudor et al., 2011 ). Kami menguji penyebaran berlebih dengan menghitung parameter penyebaran berlebih (phi) dan menemukan semua spesies yang dimodelkan menunjukkan phi > 1, yang menunjukkan penyebaran berlebih (Lampiran S3 ). Selain itu, kami menggunakan Kriteria Informasi Akaike (AIC) untuk menguji kinerja model antara regresi binomial negatif yang digelembungkan nol dan binomial negatif. Dalam semua kasus, binomial negatif yang digelembungkan nol berkinerja lebih baik daripada binomial negatif. Kedua model tersebut disusun dengan hasil tangkapan sebagai variabel respons dan tahun sebagai variabel prediktor. Regresi ini dijalankan hanya untuk tahun-tahun pasca-penghapusan (2001 dan seterusnya) untuk mencerminkan respons terhadap durasi upaya penghapusan ikan bass mulut kecil. Dengan demikian, regresi tersebut tidak selalu mencerminkan pergeseran antara keadaan sebelum penghapusan dan keadaan pasca-penghapusan.
Kami juga meneliti perubahan ukuran tubuh (diukur sebagai panjang total rata-rata semua individu yang ditangkap dan diukur pada tahun tertentu) dari setiap spesies ikan sepanjang waktu. Analisis struktur ukuran kami mencerminkan analisis kelimpahan populasi, dengan analisis titik perubahan diikuti oleh model regresi, kecuali bahwa kami menyesuaikan regresi linier dengan data panjang total. Mengingat kepadatan spesies tertentu yang rendah sebelum pemindahan, data panjang ikan tidak dikumpulkan secara ekstensif sebelum tahun 2000, yang kami gunakan untuk mewakili ukuran sebelum pemindahan. Oleh karena itu, kami membandingkan data tahun 2000 dan 2001 dengan uji peringkat bertanda Wilcoxon. Data untuk setiap tahun mencakup panjang total semua individu yang diukur yang dikumpulkan melalui penangkapan ikan dengan alat elektro.
3 HASIL
3.1 Dampak pemindahan ikan terhadap kelimpahan relatif dan struktur ukuran ikan bass mulut kecil
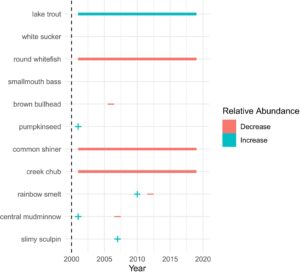
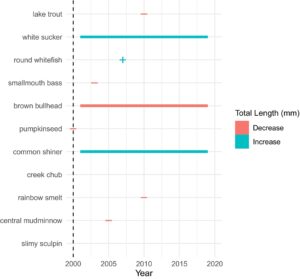
Penurunan jangka pendek dalam kelimpahan relatif smallmouth bass dan tidak ada perubahan dalam panjang total diamati pada tahun pertama setelah penghilangan smallmouth bass (Gambar 1 dan 2 ; Tabel 2 dan 3 ). Data tangkapan smallmouth bass terdispersi berlebihan (phi > 1), dan binomial negatif yang digelembungkan nol memberikan kecocokan yang lebih baik daripada binomial negatif, seperti yang ditunjukkan oleh nilai AIC yang lebih rendah (Lampiran S3 ). Mengingat hal ini, binomial negatif yang digelembungkan nol untuk Smallmouth bass tidak menunjukkan titik perubahan yang signifikan atau pergeseran bertahap selama percobaan dalam kelimpahan relatif, yang menunjukkan pola stabilitas jangka panjang (Gambar 3 dan 5 ). Pada tahun 2003, bass menunjukkan pengurangan dalam panjang totalnya (Gambar 4 dan 5 ; Tabel 3 ).


| Tahun 2000 dibandingkan dengan tahun 2001 | ||||
|---|---|---|---|---|
| CPU tahun 2000 | CPUE tahun 2001 | nilai p | Statistik W | |
| Ikan lele coklat | 1.76 | 3.11 | 0.468 | 374.5 |
| Ikan chub sungai | 1.51 | 57.02 | <0,0001* | 146 |
| Ikan pari biasa | 3.01 | 61.70 | 0,037* | 302.5 |
| Ikan trout danau | 1.07 | 1.25 | 0.254 | 355.5 |
| Lumpur tengah | 0.16 | pukul 16.00 | 0,014* | 300.5 |
| Biji labu | 31.98 | 60.36 | 0,055 | 296 |
| Ikan smelt pelangi | 0,03 | 0.56 | 0,026* | 333.5 |
| Ikan putih bulat | 0.00 | 0.42 | 0,026* | 350 |
| Ikan bass mulut kecil | 173.41 | 71.03 | <0,0001* | 727 |
| Patung berlendir | 1.23 | 6.64 | 0,065 tahun | 307 |
| Pengisap putih | 3.70 | tanggal 23.09 | <0,0001* | 92 |
Catatan : Tanda bintang menunjukkan signifikansi pada α = 0,05.
| Tahun 2000 dibandingkan dengan tahun 2001 | ||||
|---|---|---|---|---|
| 2000 TL (miliar) | 2001 TL (mm) | nilai p | Statistik W | |
| Ikan lele coklat | 243.39 | 174.42 | <0,0001* | 1792.5 |
| Ikan chub sungai | 127.15 | 76.70 | <0,0001* | 23273.5 |
| Ikan pari biasa | 94.78 | 54.20 | <0,0001* | 22436.5 |
| Ikan trout danau | 455.52 | 276.47 | 0,003* | 389 |
| Lumpur tengah | 71.67 | 59.17 | Bahasa Indonesia | Bahasa Indonesia |
| Biji labu | 93.04 | 82.17 | <0,0001* | 852.306 orang |
| Ikan smelt pelangi | Rp 117.000 | 134.00 | Bahasa Indonesia | Bahasa Indonesia |
| Ikan putih bulat | Bahasa Indonesia | 179.43 | Bahasa Indonesia | Bahasa Indonesia |
| Ikan bass mulut kecil | 170.54 | 169.62 | 0,787 tahun | 1.715.677 orang |
| Patung berlendir | 65.50 | 67.43 | 0,360 | 2453.5 |
| Pengisap putih | 304.81 | 137.35 | <0,0001* | 24.198 orang |
Catatan : Tanda bintang menunjukkan signifikansi pada α = 0,05.

3.2 Respon masyarakat pesisir terhadap penekanan populasi ikan bass
Semua spesies menunjukkan setidaknya satu jenis perubahan dalam periode survei setelah dimulainya penekanan ikan bass. Antara tahun pertama penekanan dan tahun berikutnya, enam dari 10 spesies yang disurvei mengalami peningkatan kelimpahan relatif (Gambar 1 ; Tabel 2 ). Dua dari enam spesies ini mempertahankan peningkatan ini dalam jangka pendek (<5 tahun; Gambar 3 dan 5 ; Weidel et al., 2007 ).
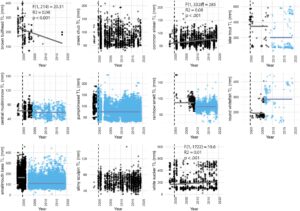
Meskipun terjadi peningkatan umum dalam kelimpahan oleh sebagian besar ikan, respons jangka panjang sangat bervariasi dan, dalam beberapa kasus, berbeda dari respons awal. Pergeseran mendadak dan perubahan bertahap seiring waktu adalah respons umum yang serupa selama durasi studi. Pergeseran cepat, dengan titik perubahan terjadi dalam lima tahun atau kurang setelah dimulainya pemindahan ikan bass, lebih jarang terjadi daripada titik perubahan yang tertunda (Gambar 3 ). Lima spesies tidak menunjukkan pergeseran cepat dan diuji untuk overdispersi dan kecocokan model. Semua spesies menunjukkan overdispersi (rentang phi 7,16–187,96) dan nilai AIC yang lebih rendah ketika dipasang dengan binomial negatif yang digelembungkan nol (Lampiran S5 ). Tiga spesies menunjukkan penurunan bertahap dalam kelimpahan relatif seiring waktu setelah peningkatan tahun pertama: common shiner ( b = 0,348, SE = 0,09, z = −3,99, p < 0,001), creek chub ( b = −0,365, SE = 0,08, z = −4,45, p < 0,001) dan round whitefish ( b = −0,55, SE = 0,19, z = −2,96, p < 0,01). Hanya lake trout ( b = 0,212, SE = 0,08, z = −2,68, p < 0,01) yang menunjukkan peningkatan bertahap dalam kelimpahan relatif seiring waktu.
3.3 Respon masyarakat pesisir dalam skala besar
Perubahan jangka panjang dalam ukuran ikan kontras dengan perubahan dalam kelimpahan relatif. Perbandingan tahun-ke-tahun antara tahun 2000 dan 2001 menunjukkan bahwa ukuran tubuh brown bullhead, creek chub, pumpkinseed, brook trout, common shiner, white sucker dan lake trout semuanya menurun segera setelah dimulainya penekanan bass (Gambar 2 ; Tabel 3 ). Pengurangan ukuran ini disertai dengan peningkatan kelimpahan relatif dalam setiap kasus. Slimy sculpin tidak menunjukkan perubahan dalam ukuran rata-rata. Tangkapan central mudminnow dan rainbow smelt terlalu rendah pada tahun 2000 untuk membandingkan panjang total tahun 2000 dan 2001. Rainbow smelt ( Osmerus mordax ), lake trout, smallmouth bass, central mudminnow, dan round whitefish menunjukkan titik perubahan dalam panjang total (Gambar 3 ). Semua titik perubahan ini negatif, kecuali untuk round whitefish, dan tidak ada spesies tersebut yang menunjukkan titik perubahan kedua. Seperti perubahan dalam kelimpahan relatif, ada beberapa perubahan bertahap, dan perubahan tersebut tidak konsisten secara arah; white sucker ( R 2 = 0,01, F (1,1722) = 19,5, p < 0,001) dan common shiner ( R 2 = 0,08, F (1,3328) = 285, p < 0,001) bertambah panjang seiring waktu, sementara brown bullhead berkurang ( R 2 = 0,08, F (1,214) = 20,3, p < 0,001). Baik slimy sculpins maupun creek chub tidak menunjukkan titik perubahan atau regresi signifikan seiring waktu (Gambar 6 ).
4 DISKUSI
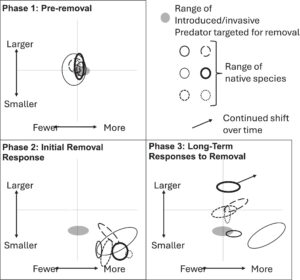
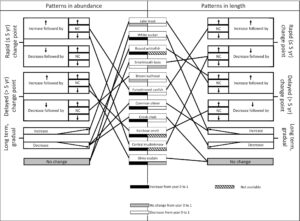
Penghapusan ikan bass mulut kecil di Little Moose Lake memiliki efek jangka pendek langsung pada sebagian besar spesies ikan, tetapi penilaian jangka panjang berikutnya menunjukkan bahwa respons jangka pendek tersebut sebagian besar bersifat sementara. Pola-pola ini menyoroti pentingnya mendefinisikan tujuan jangka panjang dan memantau pola sementara dan meletakkan kerangka kerja untuk kerangka kerja tiga fase konseptual yang mencakup fase pra-penghapusan, fase respons awal, dan fase jangka panjang (Gambar 7 ). Pada fase pertama, pra-penghapusan, predator yang diperkenalkan atau invasif mendominasi mangsa asli, yang mengakibatkan sedikit perubahan seiring waktu dalam ukuran atau kelimpahan taksa apa pun. Pada fase respons awal berikutnya, perekrutan anak-anak meningkat seiring dengan berkurangnya pemangsaan pada individu-individu ini. Akhirnya, pada fase respons jangka panjang, jaring makanan menjadi dinamis dengan beberapa penataan ulang saat hubungan predator-mangsa berubah dan spesies menjadi dewasa pada tingkat yang berbeda. Kerangka kerja ini menjelaskan mengapa peningkatan jangka pendek (<5 tahun) dalam kelimpahan relatif dan penurunan ukuran jarang selaras dengan pola jangka panjang dalam penelitian saat ini. Karena spesies asli yang ditekan meningkat dalam kelimpahan relatif setelah penghilangan penyerbu, respons yang tertunda mungkin terjadi karena lanskap persaingan dan risiko predasi berubah. Perubahan yang sedang berlangsung kemudian dapat terus terjadi karena hubungan predator-mangsa secara berurutan dibangun kembali dan diputus. Perbedaan spesies dalam sifat riwayat hidup, seperti rentang hidup dan waktu hingga dewasa, dapat semakin mengacaukan pembentukan hubungan predator-mangsa yang stabil (Tabel 1 ). Selain itu, metrik yang dievaluasi untuk menilai pemulihan dari penghilangan spesies yang diperkenalkan dan invasif akan memengaruhi bagaimana respons oleh organisme lain ditafsirkan. Seperti yang ditunjukkan di sini, perubahan dalam ukuran populasi atau struktur ukuran yang menyimpang dapat menunjukkan peningkatan sementara pada juvenil. Hasil kami menunjukkan bahwa penghilangan predator untuk mencapai tujuan pemulihan komunitas ikan perlu dinilai dalam jangka waktu yang lama, dengan mempertimbangkan kelimpahan relatif dan struktur ukuran populasi ikan yang ditargetkan untuk pemulihan.
Komunitas ikan pesisir sebelum dimulainya pemindahan mengindikasikan bahwa pengenalan ikan bass mulut kecil telah sangat mengurangi kelimpahan ikan mangsa asli, seperti yang telah didokumentasikan dalam penelitian sebelumnya (Vander Zanden et al., 1999 ). Meskipun degradasi komunitas disebabkan oleh ikan bass mulut kecil selama 50 tahun, respons dalam komunitas ikan asli menunjukkan bahwa pengelolaan aktif dapat memberikan perbaikan. Perbaikan yang berhasil dalam komunitas asli setelah penekanan ikan yang diperkenalkan telah didokumentasikan dalam respons komunitas makroinvertebrata dan zooplankton setelah penghentian penebaran ikan trout di danau California yang tinggi (Knapp et al., 2005 ). Selain itu, ikan trout cutthroat Yellowstone ( Onchorhynchus clarkia bouvieri ) telah meningkat kelimpahannya sebagai respons terhadap upaya untuk menekan ikan trout danau yang diperkenalkan (Koel et al., 2020 ).
Peningkatan jangka pendek pada ikan bass non-smallmouth muda setelah dimulainya pemindahan ikan bass smallmouth jarang diterjemahkan menjadi perubahan kelimpahan yang stabil dan jangka panjang; deskripsi sebelumnya tentang perubahan dalam struktur jaring makanan Little Moose Lake dalam beberapa tahun pertama pemindahan ikan bass (Lepak et al., 2006 ; Weidel et al., 2007 ) tidak mencerminkan dinamika jangka panjang yang dilaporkan di sini. Perubahan bertahap dan lambat setelah pengurangan kelimpahan spesies yang diperkenalkan atau invasif juga telah didokumentasikan dalam sistem lain yang mengalami perubahan mendadak pada spesies kunci. Misalnya, ikan asli terbukti terus berfluktuasi dalam kelimpahan beberapa dekade setelah pengurangan ikan alewife yang diperkenalkan ( Alosa pseudoharengus ; Bunnell et al., 2006 ). Demikian pula, dan yang mengejutkan, meskipun invertebrata memiliki tingkat pergantian yang tinggi, komunitas zooplankton dan makroinvertebrata bentik tidak mencerminkan sistem yang tidak terinvasi hingga hampir 20 tahun setelah penghentian penebaran di danau yang secara historis tidak memiliki ikan (Knapp et al., 2005 ).
Pengenalan centrarchid predator dapat diikuti oleh peningkatan ukuran tubuh mangsa asli karena predator terbatas pada individu yang lebih kecil dari batasan gape (Cerrilla et al., 2022 ). Meskipun sebagian besar spesies ikan di Little Moose Lake menunjukkan respons dalam kelimpahan dan ukuran, ada sedikit konsistensi dalam dan antara metrik karena responsnya bertahap dan tiba-tiba dan tidak seragam dalam arahnya (Gambar 8 ). Di luar peningkatan kelimpahan awal dan penurunan ukuran, responsnya sangat bervariasi. Misalnya, spesies dengan titik perubahan yang dapat dideteksi menunjukkan berbagai pola; setelah peningkatan awal, beberapa perubahan kelimpahan terjadi segera setelah pemindahan ikan bass dimulai sementara yang lain tertunda. Spesies yang diberikan tidak punah, respons cepat mungkin telah diharapkan karena ada perubahan cepat dalam demografi predator yang diperkenalkan, dan ikan predator yang diperkenalkan dan invasif sering terbukti menekan komunitas secara negatif melalui konsumsi langsung dan persaingan (Brown et al., 2009 ; Findlay et al., 2000 ; Jackson & Mandrak, 2002 ; Kirk et al., 2022 ; Weidel et al., 2000 ). Ikan muda kemungkinan besar akan sangat rentan karena smallmouth bass adalah predator anggota komunitas berbadan kecil (Weidel et al., 2000 ). Selain itu, kehadiran bass mungkin telah menyebabkan perubahan perilaku dalam penggunaan habitat (MacRae & Jackson, 2001) atau ekologi mencari makan (Kirk et al., 2022 ).
Pelepasan cepat dari konsumsi dan kompetisi kemungkinan besar berkontribusi pada peningkatan besar dalam kelimpahan relatif segera setelah dimulainya pemindahan. Namun, respons selanjutnya kemungkinan besar didorong oleh pergeseran dalam kompetisi homo- dan heterospesifik dinamis dan pemangsaan oleh spesies asli. Secara khusus, kemungkinan besar meskipun pemangsaan oleh smallmouth bass sangat berkurang, persaingan heterospesifik kemungkinan besar terjadi akibat peningkatan small bass yang diamati di danau ini karena kompensasi berlebihan (Zipkin et al., 2008 ). Selain itu, peningkatan ukuran predator puncak asli, ikan trout danau, dan perubahan pola makan yang dicatat setelah penekanan bass (Lepak et al., 2006 ) kemungkinan besar memiliki pengaruh tambahan pada lintasan anggota komunitas ikan asli dengan memberikan peningkatan pemangsaan dan perubahan perilaku pemangsaan yang mungkin telah menimbulkan perubahan dalam perilaku mangsa. Perubahan dalam kelimpahan spesies invasif dapat menyebabkan invasi sekunder dan perubahan dinamika predator-mangsa (Poos et al., 2010 ; Woodford et al., 2024 ). Dalam penelitian kami, pengurangan tekanan predasi dari smallmouth bass kemungkinan menyebabkan peningkatan rainbow smelt non-asli, yang menunjukkan pertumbuhan eksponensial selama hampir satu dekade sebelum mencapai puncaknya dan kemudian stabil pada tingkat yang lebih rendah dari puncaknya tetapi lebih tinggi dari sebelum penghilangan smallmouth bass yang dikelola. Lonjakan awal dalam invasi sekunder kemungkinan didorong oleh berkurangnya tekanan predasi, sementara penurunan berikutnya kemungkinan dipengaruhi oleh peningkatan predasi dari ikan trout danau, predator puncak asli. Dinamika kompleks tersebut, termasuk invasi sekunder dan penekanannya pada akhirnya oleh predator asli, menggarisbawahi pentingnya memahami interaksi predator-mangsa dan ketahanan ekosistem ketika mengembangkan strategi untuk pengelolaan spesies yang diperkenalkan dan invasif.
Mengingat sebagian besar studi penyingkiran spesies yang diperkenalkan dan invasif berfokus pada respons jangka pendek kurang dari 5 tahun (72%) (Prior et al., 2018 ), kemungkinan besar studi ini tidak mencerminkan kemungkinan hasil jangka panjang dari restrukturisasi komunitas yang dihasilkan dari upaya ini; pola jangka panjang yang menyimpang dari pola jangka pendek dalam menanggapi perubahan dalam kelimpahan predator dominan kemungkinan umum dan kurang terwakili dalam studi yang dipublikasikan. Studi saat ini menunjukkan bahwa meskipun banyak perubahan berkekuatan besar terjadi dalam 5 tahun pertama setelah dimulainya penyingkiran, banyak spesies terus mengalami perubahan seiring waktu, yang menunjukkan bahwa bahkan 20 tahun mungkin tidak cukup untuk mengidentifikasi apakah suatu populasi mencapai keadaan di mana ia beroperasi dalam periode jangka panjang setelah gangguan penyingkiran.
Upaya pengelolaan dalam ekosistem akuatik sering kali dinilai berdasarkan perubahan kelimpahan (Koel et al., 2020 ; Weidel et al., 2007 ), dengan lebih sedikit penelitian yang mencakup perubahan dalam struktur ukuran (Sass et al., 2022 ). Pola dalam struktur ukuran, yang merupakan fungsi dari perubahan dalam struktur populasi dan pertumbuhan, awalnya merespons dengan cepat melalui peningkatan kelimpahan juvenil dan kemudian tertinggal ketika juvenil menjadi lebih melimpah atau terjadi pergeseran demografi (Gambar 8 ). Satu-satunya pengecualian adalah smallmouth bass, yang mengalami ukuran populasi yang stabil dengan penurunan tiba-tiba dalam panjang total, mendukung kompensasi berlebihan (Zipkin et al., 2008 ). Pergeseran demografi jangka panjang bergantung pada beberapa faktor termasuk kekuatan kelompok, usia kedewasaan, dan harapan hidup. Populasi yang menunjukkan penurunan panjang total yang cepat dan bertahap seiring waktu kemungkinan mengalami pergeseran dalam struktur usia populasi dan/atau perubahan pertumbuhan. Meskipun jarang, respons yang cepat dan lambat dalam struktur ukuran merupakan karakteristik klasik dari perubahan atau degradasi ekosistem (Broadway et al., 2015 ; Walters & Post, 2008 ). Respons yang lambat dalam struktur ukuran, misalnya, juga terlihat di Danau Yellowstone, di mana struktur ukuran ikan trout cutthroat asli Oncorhynchus clarkii belum stabil setelah lebih dari satu dekade pemindahan intensif ikan trout danau invasif (Koel et al., 2020 ). Oleh karena itu, pemantauan jangka panjang terhadap populasi dan struktur ukuran sangat penting untuk memahami pendorong pemulihan yang mendasarinya.
Replikasi dalam waktu dan ruang sangatlah ideal, tetapi ketika unit ekologi utama sulit untuk direplikasi, eksperimen skala besar pada satu ekosistem sangatlah berharga untuk memahami dinamika ekologi yang kompleks (Schindler, 1998 ). Hal ini ditunjukkan dalam beberapa studi penghilangan spesies yang diperkenalkan dan spesies invasif, seperti penghilangan ikan trout danau dari Danau Yellowstone (Koel et al., 2020 ). Seperti kebanyakan studi penghilangan, studi kami mencakup periode pra-intervensi yang singkat (Prior et al., 2018 ), tetapi tidak seperti kebanyakan studi penghilangan, studi kami mencakup periode respons yang lebih lama yang memungkinkan kami untuk lebih memahami respons jangka panjang yang potensial. Respons jangka pendek yang terdokumentasi dengan baik, tidak adanya perubahan signifikan sebelum dan sesudah penghilangan dalam kondisi fisik dan kimia (Weidel et al., 2000 ), dan kerangka konseptual kami untuk menafsirkan perubahan ekologi jangka panjang mendukung temuan kami. Pengujian lebih lanjut terhadap kerangka kerja ini, khususnya yang berkenaan dengan respon cepat ikan-ikan muda dan efek-efek yang rumit dan tertunda dari waktu ke waktu, akan meningkatkan strategi pengelolaan bagi ekosistem yang besar dan dinamis untuk semua upaya pengelolaan ekosistem dan bukan hanya upaya-upaya penyingkiran spesies yang diperkenalkan atau spesies invasif.
Akhirnya, penting untuk dicatat bahwa data yang dikumpulkan untuk studi saat ini hanya berasal dari daerah dangkal, dekat pantai, yang berpotensi mengaburkan respons beberapa spesies. Oleh karena itu, pergeseran penggunaan habitat dapat mengakibatkan pola yang, meskipun respons sebenarnya dalam kerangka pengambilan sampel kami dan terkait dengan penghilangan ikan bass, tidak mewakili atau bertentangan dengan pergeseran tingkat spesies dalam kelimpahan di seluruh danau. Meskipun di luar cakupan studi ini pada komunitas pesisir danau, perubahan perilaku, penggunaan habitat, dan distribusi perlu dipertimbangkan ketika mencoba memahami bagaimana ekosistem merespons penekanan spesies yang diperkenalkan atau invasif dan harus menjadi area penyelidikan di masa mendatang. Misalnya, ikan trout danau, spesies yang sebagian besar ditemukan di lepas pantai, beralih ke piscivory setelah dimulainya upaya penghilangan ikan bass smallmouth (Lepak et al., 2006 ). Pergeseran ini dapat secara aktif atau pasif mendorong ikan mangsa, termasuk ikan trout danau yang lebih kecil, ke perairan yang lebih dangkal untuk mengurangi risiko predasi. Perubahan dalam kehadiran predator dapat menciptakan ‘lanskap ketakutan’ akuatik (Ducsai et al., 2023 ; Hintz & Relyea, 2017 ; Wilson et al., 2021 ), yang memengaruhi perilaku mangsa. Kehadiran spesies invasif juga dapat mengubah dinamika ini (Shelton et al., 2008 ). Oleh karena itu, ada kemungkinan bahwa pemindahan yang terkelola dan perubahan selanjutnya dalam struktur ukuran dan ukuran populasi mengakibatkan perubahan dari waktu ke waktu dalam modifikasi perilaku.
5 KESIMPULAN
Evaluasi pengaruh jangka panjang penekanan spesies invasif pada komunitas ikan asli penting untuk memahami konsekuensi dari upaya ini, namun jarang dilaksanakan. Studi kasus ini menunjukkan bahwa perencanaan program pemulihan atau pengelolaan yang berhasil memerlukan pertimbangan baik jangka waktu maupun metrik yang relevan untuk setiap evaluasi keberhasilan. Hal ini khususnya penting jika jangka waktu tidak cukup panjang untuk mengamati perubahan dalam kelimpahan atau distribusi ukuran spesies yang berumur panjang atau yang tumbuh lebih lambat, yang mungkin dipengaruhi oleh pergeseran dalam ekologi mencari makan atau interaksi tidak langsung di antara spesies yang terdampak. Karena metrik dapat merespons secara berbeda dan berdasarkan respons yang berbeda tersebut menunjukkan pola pemulihan yang berbeda, strategi pengelolaan harus memperhitungkan respons yang tertinggal dan secara jelas menetapkan target respons ekosistem untuk upaya pemulihan. Penekanan fisik ikan bass mulut kecil telah mengubah lanskap komunitas ikan di Little Moose Lake, meningkatkan perikanan ikan trout danau dan, setidaknya pada awalnya, meningkatkan kelimpahan relatif banyak spesies asli. Namun, penurunan yang tertunda dalam kelimpahan relatif beberapa spesies telah terjadi sebagai respons terhadap upaya pengelolaan ini. Investigasi lebih lanjut terhadap lanskap persaingan dan dinamika komunitas skala kecil di seluruh lokasi dan habitat dari waktu ke waktu kemungkinan akan menyoroti pergeseran demografi dan jaring makanan kompleks yang mengatur dinamika komunitas dan aliran energi.


