
ABSTRAK
Banyak spesies burung penyanyi menggunakan pengenalan vokal individual dalam perilaku sosial mereka. Peneliti biasanya menggunakan tugas diskriminasi individual, seperti tugas pengkondisian operan Go/No-go, untuk menilai pengenalan vokal individual. Beberapa vokalisasi chickadee bertopi hitam ( Poecile atricapillus ) mengandung fitur-fitur yang berbeda secara individual yang dapat digunakan untuk diskriminasi individual. Namun, tidak semua vokalisasi tersebut telah diuji untuk pengenalan individu dengan burung hidup. Selain itu, generalisasi vokalisasi silang dari diskriminasi individu yang dipelajari belum diuji. Generalisasi seperti itu akan menguntungkan bagi chickadee, karena chickadee sering berkomunikasi di luar kontak visual dan menggunakan komunikasi vokal untuk memandu interaksi sosial mereka. Di sini kami menguji apakah chickadee bertopi hitam dapat membedakan identitas individu pemanggil dalam panggilan chickadee black-capped chick-a-dee . Kami juga bertujuan untuk menjawab apakah chickadee dapat menggeneralisasi diskriminasi individu yang dipelajari menggunakan panggilan chick-a-dee ke lagu fee-bee , dan sebaliknya. Burung chickadee bertopi hitam dilatih untuk membedakan beberapa panggilan chick-a-dee atau lagu fee-bee yang dihasilkan oleh satu burung chickadee jantan dan satu burung chickadee betina dari yang dihasilkan oleh burung chickadee jantan dan betina yang berbeda dalam eksperimen pengkondisian operan Go/No-go. Kami kemudian menguji generalisasi di seluruh jenis vokalisasi dengan menyajikan rekaman dari empat individu yang sama kepada burung, kali ini dari jenis vokalisasi yang berlawanan. Burung chickadee mampu membedakan antara individu menggunakan panggilan chick-a-dee atau lagu fee-bee tetapi tidak dapat menggeneralisasi pembelajaran ini ke jenis vokalisasi yang berlawanan. Sementara temuan kami menunjukkan bahwa burung chickadee dapat menggunakan diskriminasi individu dalam setidaknya dua jenis vokalisasi, mekanisme yang dengannya lagu dan panggilan dikenali sebagai milik individu yang sama masih belum jelas. Isyarat kontekstual eksternal dapat memainkan peran penting dalam menjembatani informasi identitas di seluruh jenis vokalisasi tersebut.
Pengenalan vokal individu (IVR) mengacu pada kemampuan penerima sinyal untuk mendapatkan informasi identitas dari vokalisasi produsen sinyal sejenis, sehingga penerima dapat membedakan satu produsen dari yang lain (Carlson et al. 2020 ). IVR dianggap bermanfaat bagi penerima dengan memungkinkan mereka untuk dengan cepat menentukan apakah, bagaimana, dan sejauh mana informasi yang terkandung dalam vokalisasi produsen relevan bagi mereka sebagai penerima (Carlson et al. 2020 ). Penerima kemudian dapat mengubah perilaku mereka dalam menanggapi konten dan konteks informasi yang diberikan oleh produsen yang sesuai, seperti menghindari agresor teritorial atau sejenis yang dominan (Lambrechts dan Dhondt 1995 ), mendekati orang asing yang berpotensi mengancam tetapi mengabaikan tetangga yang dikenal (Brooks dan Falls 1975 ), atau mendekati kerabat atau pasangan yang dalam kesulitan (Warrington et al. 2014 ). Pada gilirannya, produser mendapat manfaat dari vokalisasi mereka yang berbeda secara individual dengan mendorong kerja sama keluarga atau kelompok dari penerima (Carlson et al. 2020 ; Kastein et al. 2013 ; Zsebők et al. 2017 ). Karena IVR menggambarkan proses internal, hal itu tidak dapat diukur secara langsung (Carlson et al. 2020 ). Oleh karena itu, pengamatan eksternal terhadap perilaku individu diperlukan untuk menyimpulkan IVR, yang dapat diasumsikan ketika seorang individu secara perilaku menunjukkan kemampuan untuk membedakan antara dua atau lebih individu berdasarkan vokalisasi individu tersebut (Carlson et al. 2020 ). Diskriminasi vokal individu (ID), perilaku yang dapat diamati yang sering digunakan sebagai proksi untuk IVR, telah banyak diamati di seluruh kerajaan hewan (misalnya, Amphibia- Chuang et al. 2017 ; Feng et al. 2009 ; Mammalia- Ceugniet dan Izumi 2004 ; Kastein et al. 2013 ; Lemasson et al. 2013 ; Aves- Brooks dan Falls 1975 ; Montenegro et al. 2020 ; Weary dan Krebs 1992 ). Terlebih lagi, terdapat banyak contoh kapasitas ID yang diduga berdasarkan variabilitas vokalisasi antar individu yang diamati (misalnya, Bertucci et al. 2012 ; Bloomfield et al. 2004 ; Charrier et al. 2004 ; Frommolt et al. 2003 ; Gasser et al. 2009 ; Guillette et al. 2010 ; Mammen and Nowicki 1981 ; Volodina et al. 2006 ; Warrington et al. 2014 ; Zsebők et al. 2017).); namun, fitur akustik yang khas secara individual, meskipun diperlukan untuk ID, tidak cukup untuk menunjukkan ID, karena ID memerlukan persepsi dan penggunaan fitur individualistis yang berbeda tersebut di pihak penerima (Carlson et al. 2020 ).
Burung penyanyi menjadi model populer untuk mempelajari ID dan IVR (Carlson et al. 2020 ), setidaknya sebagian karena fakta bahwa burung penyanyi sering berkomunikasi di luar kontak visual dan sangat bergantung pada penyampaian informasi secara vokal di antara spesiesnya (Ficken et al. 1978 ; Kondo et al. 2010 ; Mammen dan Nowicki 1981 ). Dengan demikian, banyak spesies burung penyanyi telah terbukti mampu membedakan antar individu, dan melakukannya dengan menggunakan berbagai tingkat spesifisitas. Ketika penerima sinyal dapat mengidentifikasi produsen –seperti tetangga yang dikenal atau pasangan– di antara produsen lain, ini menunjukkan IVR- tunggal (Carlson et al. 2020 ). Misalnya, burung pipit berleher putih jantan ( Zonotrichia albicollis ) mendekati lebih dekat ke speaker playback, dan merespons lebih cepat, dengan tingkat nyanyian yang lebih tinggi dan lebih banyak respons terbang, terhadap lagu orang asing daripada tetangga yang dikenal dalam studi playback lapangan (Brooks dan Falls 1975 ). Demikian pula, burung pipit besar betina ( Parus major ) muncul dari sarang mereka secara eksklusif sebagai respons terhadap lagu pasangan mereka tetapi tidak terhadap lagu orang asing (Lind et al. 1996 ), dan hampir secara eksklusif terhadap lagu pasangan dibandingkan dengan lagu tetangga (Blumenrath et al. 2007 ). Dalam kasus ini, satu individu dikenali di antara yang lain pada waktu tertentu. Sebaliknya, kemampuan untuk membedakan di antara beberapa individu menunjukkan sistem pengenalan yang jauh lebih baik yang dikenal sebagai IVR- multiple , di mana penerima sinyal dapat mengenali beberapa produsen dalam satu kelompok, dan mengidentifikasi satu produsen dari yang lain (Carlson et al. 2020 ). Misalnya, panggilan kontak burung gagak hutan ( Corvus macrorhynchos ) menunjukkan variabilitas individu yang digunakan burung gagak hutan untuk membedakan antara individu yang dikenal dalam paradigma pengkondisian operan (Kondo et al. 2010 ). Demikian pula, induk burung black redstart ( Phoenicurus ochruros ) yang membelah induk merespons lebih andal terhadap panggilan mengemis dari anak burung yang mereka beri makan lebih disukai daripada terhadap anak burung yang tidak mereka beri makan lebih disukai (Draganoiu et al. 2006 ). Dalam kasus ini, penerima mampu membedakan antara vokalisasi beberapa individu, dan mengenali satu individu tertentu dari yang lain.
Burung chickadee bertopi hitam ( Poecile atricapillus ) adalah burung yang tahan dingin yang hidup hampir terus-menerus di seluruh Amerika Serikat dan Kanada (Smith 1991 , 1; Weisman dan Ratcliffe 2004 ), berkelompok secara eksklusif pada bulan-bulan musim gugur dan musim dingin, hidup monogami pada bulan-bulan musim semi dan musim panas (Smith 1991 , 6), dan komunikasi akustiknya telah dipelajari secara ekstensif. Burung chickadee bertopi hitam adalah spesies model yang ideal untuk menyelidiki IVR karena banyak perilaku teritorial dan perkembangbiakannya dimediasi oleh komunikasi vokal, dan mereka bergantung pada kemampuan penerima untuk membedakan antara individu, seperti menguping betina yang membedakan antara lagu dua jantan (Mennill dan Otter 2007 ; Wilson dan Mennill 2010 ). Satu vokalisasi umum yang diteliti dari chickadee bertopi hitam adalah panggilan chick-a-dee yang merupakan nama panggilannya , panggilan yang relatif rumit secara akustik dan sangat bervariasi yang paling sering digunakan dalam konteks sosial dan dalam koordinasi kawanan, yang terdiri dari hingga lima jenis nada (A, B, C, D, dan D-hybrid), yang mana salah satunya dapat diulang atau dihilangkan (Ficken et al. 1978 ; Smith 1991 , 66–74). Analisis bioakustik dari panggilan chick-a-dee menemukan bahwa panggilan ini lebih bervariasi antar individu daripada di dalam individu (Charrier et al. 2004 ; Mammen dan Nowicki 1981 ), yang mendukung fungsinya dalam ID. Secara khusus, Mammen dan Nowicki ( 1981 ) menemukan beberapa fitur spektral (frekuensi puncak nada D, frekuensi maksimum nada D, lebar pita nada D), temporal (durasi nada D, durasi nada pengantar, durasi panggilan total, interval antara nada pengantar, interval antara nada D, interval antara nada pengantar dan nada D), dan struktural (jumlah nada D dan jumlah nada pengantar) dari panggilan chick-a-dee berbeda secara signifikan antara individu. Demikian pula, Charrier et al. ( 2004 ) menemukan bahwa berbagai parameter akustik nada A, B, dan C berbeda secara signifikan antara individu, dan durasi nada D, frekuensi maksimum, dan frekuensi harmonik pertama yang terlihat juga berbeda secara signifikan antara individu. Sejauh pengetahuan kami, diskriminasi individu menggunakan panggilan chick-a-dee belum diuji secara empiris dengan burung.
Vokalisasi chickadee lain yang banyak diteliti adalah lagu fee-bee . Tidak seperti kebanyakan lagu burung penyanyi yang secara akustik lebih kompleks daripada panggilannya masing-masing, lagu fee-bee secara akustik lebih sederhana daripada panggilan chick-a-dee dan terutama digunakan dalam konteks teritorial dan reproduksi (Charrier et al. 2004 ; Ficken et al. 1978 ; Hahn et al. 2013 ; Smith 1991 , 57–61). Lagu fee-bee terdiri dari dua nada, dimulai dengan fee bernada tinggi yang mengamati penurunan frekuensi secara bertahap yang dikenal sebagai fee glissando, diikuti oleh lebah frekuensi yang lebih rendah (Montenegro et al. 2020 ; Hahn et al. 2013 ; Smith 1991 , 57–61; lihat ulasan dalam Weisman dan Ratcliffe 2004 ). Seperti halnya panggilan chick-a-dee , analisis bioakustik telah mengungkapkan individualitas yang signifikan dalam lagu fee-bee . Hahn et al. ( 2013 ) menemukan bahwa durasi lagu total, durasi proporsional not fee , glissando fee , frekuensi awal not lebah , rasio interval antar not, amplitudo relatif not fee dan lebah , dan rasio amplitudo akar rata-rata kuadrat not fee terhadap keseluruhan lagu semuanya merupakan fitur khas individual dari lagu fee-bee . Wilson dan Mennill ( 2010 ) juga menemukan bahwa lagu fee-bee jantan berbeda secara individual, tetapi mengambil satu langkah lebih jauh dengan menguji persepsi chickadee terhadap perbedaan individu ini. Dalam studi pemutaran ulang mereka, Wilson dan Mennill ( 2010 ) mengamati bahwa chickadee jantan tetap berada di dekat pemutaran lagu dalam wilayah mereka lebih lama, dan menghasilkan lebih banyak lagu, ketika disajikan dengan lagu dari jantan yang tidak dikenal daripada ketika disajikan dengan lagu dari jantan yang sudah disiapkan. Lebih jauh, Montenegro et al. ( 2020 ) mampu melatih chickadee bertopi hitam jantan dan betina untuk secara akurat membedakan antara lagu-lagu betina individu dalam paradigma pengkondisian operan.
Menariknya, tidak ada studi bioakustik atau perilaku yang disebutkan di atas yang menguji apakah variasi vokal individu, atau ID oleh burung, berkorelasi di seluruh jenis vokalisasi. Namun, masuk akal bahwa akan menguntungkan bagi chickadee, yang sering berkomunikasi di luar kontak visual (Ficken et al. 1978 ; Mammen dan Nowicki 1981 ), untuk dapat mengenali individu tertentu terlepas dari jenis vokalisasi tertentu yang dihasilkan, seperti yang sebelumnya telah ditunjukkan pada beberapa spesies mamalia (Cheney & Seyfarth), tetapi tidak pada burung. Sebagai contoh, salah mengidentifikasi tetangga yang dikenal yang memanggil dari dekat batas wilayah seseorang sebagai orang asing yang mengancam hanya setelah tetangga yang sama mulai bernyanyi dapat menyebabkan perilaku yang merugikan dan berpotensi berisiko yang mungkin dihindari oleh chickadee. Oleh karena itu, akan menguntungkan bagi chickadee untuk belajar mengidentifikasi individu berdasarkan satu jenis vokalisasi tunggal dan dapat menggeneralisasi pembelajaran ini di seluruh jenis vokalisasi lainnya. Karena burung chickadee sering berkomunikasi di luar kontak visual (Ficken et al. 1978 ; Mammen dan Nowicki 1981 ), akan sangat menguntungkan bagi burung chickadee untuk dapat membentuk generalisasi IVR ini berdasarkan informasi yang tersedia yang terkandung dalam vokalisasi.
Sementara analisis bioakustik menunjukkan bahwa panggilan chick-a-dee dan lagu fee-bee keduanya khas secara individu, hanya yang terakhir yang telah diuji dengan burung. Tujuan pertama dari studi saat ini adalah untuk menjawab apakah asumsi lama ID dalam panggilan chick-a-dee benar-benar berlaku pada chickadee bertopi hitam. Tujuan kedua kami adalah untuk menjawab apakah chickadee bertopi hitam dapat menggeneralisasi ID yang dipelajari di seluruh jenis vokalisasi, hanya berdasarkan pada variasi khas individu yang terkandung dalam vokalisasi itu sendiri. Kami meramalkan bahwa chickadee bertopi hitam yang dilatih dalam paradigma go/no-go pengkondisian operan, berdasarkan metode Montenegro et al. ( 2020 ) akan dapat berhasil membedakan antara individu menggunakan panggilan chick-a-dee yang direkam atau lagu fee-bee . Kami selanjutnya meramalkan bahwa chickadee akan terus membedakan antara individu yang sama bahkan ketika disajikan dengan rekaman jenis vokalisasi yang awalnya tidak mereka latih.
1 Bahan dan Metode
1.1 Subjek
Sebelum penelitian, semua chickadee ditempatkan di 1/4 segmen kandang King’s Cages Superior Stack Breeder (segmen individu: 50,8 × 50,8 × 67,31 cm; King’s Cages, East Brunswick, New Jersey) di mana mereka memiliki kontak visual dan vokal dengan chickadee lain di dalam ruang koloni. Setiap segmen kandang dilengkapi dengan tiga atau lebih tempat bertengger, dan burung diberi akses gratis ke makanan (Mazuri Insectivore Diet; Mazuri, St. Louis, MO, AS), air, grit cangkang tiram, dan tulang sotong. Burung diberi suplemen enam hingga sepuluh biji bunga matahari tanpa kulit dan setengah kacang tanah setiap hari, telur rebus dicampur dengan sayuran cincang (bayam atau peterseli) dua kali seminggu, dan dengan ulat hongkong ( Tenebrio molitor ) yang ditaburi larutan vitamin (Prime Vitamin Supplement; Hagen Inc., Montreal, QB, Kanada) tiga kali seminggu. Ruang koloni diberi pencahayaan sesuai dengan siklus cahaya: gelap alami untuk Edmonton, Alberta, Kanada, dan dijaga pada suhu 20°C. Dua puluh burung chickadee bertopi hitam dimasukkan dalam studi pengkondisian operan ini, masing-masing secara acak dimasukkan ke dalam satu dari empat kelompok studi (lihat bagian Pelatihan Diskriminasi di bawah). Enam belas burung chickadee ditangkap dari EH Moss Forest Reserve di Lembah Sungai Edmonton, Edmonton, Alberta, Kanada, antara Februari 2020 dan Februari 2023. Empat burung lainnya ditangkap dari Mill Creek Ravine di Lembah Sungai Edmonton antara Januari 2019 dan Desember 2022. Burung-burung tersebut hidup bervariasi dari Oktober 2023 hingga April 2024, dengan rata-rata lebih dari 2 tahun di lab sebelum memulai studi. Burung-burung yang ditangkap sebelumnya berpartisipasi dalam studi lain yang tidak terkait selama waktu ini. Ke-20 burung tersebut ditangkap saat dewasa (setidaknya berusia 1 tahun) sebagaimana dinilai dengan memeriksa retrik ekor luar (Pyle 1997 ), dan jenis kelamin mereka ditentukan dengan analisis DNA dari sampel darah (Griffiths et al. 1998 ). Selama penelitian, burung chickadee melanjutkan Diet Mazuri Insectivore melalui pengumpan ketika merespons dengan benar terhadap uji coba pengkondisian operan. Burung chickadee juga diberi air, tulang sotong, dan grit cangkang tiram sepuasnya . Burung chickadee diberi suplemen dua setengah kacang tanah dan tiga ulat hongkong setiap hari.
1.2 Rangsangan Akustik
Sebanyak 176 stimulus, 88 panggilan chick-a-dee dan 88 nyanyian fee-bee , digunakan dalam penelitian ini. Dari jumlah tersebut, setiap burung diberikan 132 stimulus ini tergantung pada kelompoknya (lihat bagian Prosedur di bawah untuk alokasi stimulus khusus). Stimuli dibuat dari rekaman yang diambil dari dua burung chickadee bertopi hitam jantan dan dua betina yang ditangkap di Lembah Sungai Edmonton antara Januari 2009 dan Januari 2014 saat dewasa. Tidak ada burung stimulus yang ditempatkan di laboratorium pada waktu yang sama dengan burung-burung yang digunakan dalam penelitian ini. Burung stimulus direkam secara individual di dalam ruang peredam suara (1,7 × 0,84 × 0,58 m; Industrial Acoustics Company, Bronx, NY) menggunakan mikrofon AKG C1000S (AKG Acoustics, Vienna, Austria) yang diposisikan 10 cm di atas dan di bagian belakang kandang yang terhubung ke perekam digital Marantz PMD670 (Marantz America, Mahwah, NJ) (16 bit, laju pengambilan sampel 44.100 Hz). Rekaman diambil antara tahun 2012 dan 2017. Sebanyak 22 panggilan chick-a-dee dan 22 lagu fee-bee yang dihasilkan oleh setiap chickadee stimulus diisolasi dari rekaman ini menggunakan perangkat lunak SIGNAL 5.03.11 (Engineering Design, Berkley, CA, AS). Vokalisasi individual terlebih dahulu difilter bandpass (bandpass bawah 400 Hz, bandpass atas 1400 Hz) untuk mengurangi derau latar belakang (Montenegro et al. 2020 ) menggunakan GoldWave versi 6.31 (GoldWave Inc., St. John’s, NL, Kanada), sebelum 5 ms keheningan ditambahkan ke ujung awal dan akhir vokalisasi, dan stimulus dikurangi secara bertahap untuk mengurangi “popping” selama pemutaran dan dinormalisasi (RMS 1) menggunakan perangkat lunak SIGNAL 5.03.11.
1.3 Peralatan
Saat menyelesaikan penelitian, burung ditempatkan di kandang koloni Jupiter Parakeet yang dimodifikasi (30 cm × 30 cm × 40 cm; Rolf C. Hagen Inc., Montreal, QB, Kanada) yang terletak di dalam ruang operan peredam suara (lihat Gambar 1 ). Kandang Jupiter dimodifikasi dengan bukaan (11 cm × 16 cm) di sisi kiri kandang untuk memungkinkan burung mengakses pengumpan otomatis eksternal, di mana akses ke pakan dapat diakses secara kondisional. Kandang Jupiter juga dilengkapi dengan tempat bertengger atas permintaan yang dipasang dengan sensor cahaya inframerah yang diamankan 10 cm dari bukaan pengumpan, dua tempat bertengger tanpa permintaan (satu di tengah dan satu di sepertiga kanan kandang), dan tulang sotong, cangkir grit, botol air, dan pada hari pertama pra-pelatihan dan selama “hari istirahat” berkala (2 hari selama pra-pelatihan dan 1 hari setelah setiap uji probe), cangkir pakan gratis semuanya di dinding kanan kandang. Di dalam ruang operan juga terdapat pengeras suara rentang penuh Fostex (model FE108 Σ atau FE108E Σ; Fostex, Jepang; rentang respons frekuensi: 80–18.000 Hz) yang terletak di samping pengumpan di bagian depan sangkar. Pengumpan otomatis, pengeras suara, dan tempat bertengger permintaan dikontrol melalui komputer papan tunggal (Palya dan Walter 1993 ) yang terhubung ke komputer pribadi dan Penguat Terpadu Cambridge (model A300, rentang respons frekuensi: 25–60.000 Hz atau Azur 640A, rentang respons frekuensi 4–80.000 Hz; Cambridge Audio, London, Inggris; lihat Sturdy dan Weisman ( 2006 ) untuk deskripsi terperinci tentang pengaturan serupa) yang terletak di ruang yang berdekatan. Ruang operan diterangi berdasarkan cahaya alami: pola gelap untuk Edmonton, Alberta, Kanada menggunakan bohlam LED spektrum penuh (3 W, 250 lm E26, Not-Dim, 5000 K; Lohas LED, Chicago, Illinois, AS).
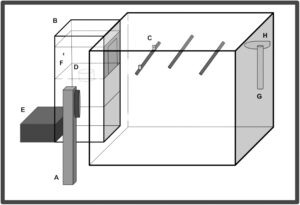
1.4 Prosedur
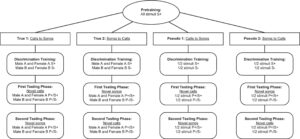
Singkatnya, burung dalam kelompok Sejati dilatih untuk membedakan suara chick-a-dee atau lagu fee-bee berdasarkan produser individu, sementara burung dalam kelompok Semu dilatih untuk menghafal suara chick-a-dee atau lagu fee-bee . Kami menguji pembelajaran ini terlebih dahulu dengan uji generalisasi tipe dalam-vokalisasi, dan kemudian dengan uji generalisasi tipe lintas-vokalisasi (Gambar 2 ).
1.5 Pra-pelatihan
Sebelum memulai pelatihan diskriminasi, burung pertama-tama dilatih untuk menggunakan tempat bertengger dan tempat makan melalui pembentukan yang berurutan. Burung belajar untuk mendarat di tempat bertengger yang diminta, dengan demikian menghalangi sinar inframerah, yang akan memicu stimulus untuk diberikan. Burung awalnya dilatih dengan stimulus visual (lampu isyarat merah) sebelum beralih ke stimulus akustik 1 detik (nada 1000 Hz). Burung akan menanggapi stimulus ini dengan meninggalkan tempat bertengger yang diminta dan bergerak ke tempat makan, di mana mereka akan menghalangi sinar inframerah lainnya, yang memicu wadah makanan untuk naik ke atas tempat makan, di mana ia dapat diakses oleh burung selama 2 detik. Setelah burung menguasai rutinitas ini, stimulus akustik diganti dengan serangkaian 132 stimulus yang akan didengar burung chickadee selama penelitian. Langkah ini mendorong tingkat respons yang tinggi dalam tahap-tahap yang berurutan dan mengoreksi respons diferensial yang tidak diinginkan. Di sini, burung memicu tempat bertengger yang diminta, yang mendorong salah satu dari 132 stimulus untuk dimainkan. Semua stimulus dimainkan dalam urutan acak tanpa penggantian di setiap blok percobaan. Chickadee diminta untuk duduk di tempat bertengger permintaan selama 900–1100 ms untuk memicu tempat bertengger tersebut dan tetap di tempat bertengger tersebut cukup lama untuk mendengarkan seluruh stimulus (1117–3891 ms) sebelum memasuki tempat makan untuk menerima akses 2 detik ke makanan. Jika burung meninggalkan tempat bertengger permintaan sebelum stimulus selesai dimainkan, mereka dihukum dengan kegelapan selama 30 detik. Periode kegelapan yang singkat umumnya digunakan untuk menandai dan mengurangi respons yang tidak tepat (misalnya, Montenegro et al. 2020 ; Sturdy dan Weisman 2006 ). Periode interpercobaan selama 30 detik, atau 60 detik jika burung gagal merespons stimulus dengan memasuki tempat makan, diperlukan sebelum burung dapat memicu kembali tempat bertengger permintaan. Karena tingkat respons yang tinggi diperlukan untuk memperoleh data yang andal pada tahap diskriminasi selanjutnya, semua chickadee diminta untuk merespons minimal 60% percobaan untuk minimal 3168 percobaan. Untuk memastikan burung tidak memasuki tugas diskriminasi dengan bias yang sudah ada sebelumnya, masing-masing burung harus secara konsisten menunjukkan, selama minimal 2112 kali percobaan, tidak lebih dari 10% perbedaan dalam tingkat respons antara panggilan dan lagu dan antara stimulus yang nantinya akan diperkuat (S+) atau dihukum (S−) selama setiap tahap diskriminasi.
1.6 Pelatihan Diskriminasi
Kelompok eksperimen dilatih untuk membedakan suara chick-a-dee (Benar 1) atau lagu fee-bee (Benar 2) berdasarkan identitas individu. Kelompok kontrol dilatih untuk membedakan suara chick-a-dee (Pseudo 1) atau lagu fee-bee (Pseudo 2) secara acak. Setiap kelompok Benar terdiri dari empat jantan dan dua betina, sedangkan setiap kelompok Pseudo terdiri dari tiga jantan dan satu betina.
Selama pelatihan diskriminasi awal, chickadee diberikan 44 stimulus, 11 dari masing-masing dari empat burung stimulus yang terekam. Seperti dalam prapelatihan, chickadee mengoperasikan peralatan dengan terlebih dahulu mendarat di tempat bertengger yang diminta, yang memicu stimulus untuk bermain. Untuk memastikan seluruh stimulus diperhatikan, chickadee kembali dilatih untuk tetap berada di tempat bertengger selama seluruh durasi stimulus dan dihukum dengan kegelapan selama 30 detik jika mereka meninggalkan tempat bertengger lebih awal. Setelah stimulus selesai dimainkan, chickadee memiliki waktu 1 detik untuk memasuki tempat makan (Go) atau mengabaikan stimulus dengan meninggalkan tempat bertengger tetapi tidak memasuki tempat makan (No-go). Ketika chickadee memasuki tempat makan dengan benar selama uji coba S+, mereka segera diberi hadiah dengan akses selama 2 detik ke cangkir makanan mereka, diikuti oleh interval antar uji coba selama 30 detik. Ketika chickadee salah memasuki tempat makan selama uji coba S−, mereka segera dihukum dengan lampu padam selama 30 detik. Mengabaikan stimulus selama percobaan S+ atau S− menghasilkan interval antarpercobaan 0,5 detik. Tetap di tempat bertengger (tidak Lanjut atau Tidak Lanjut) menghasilkan interval antarpercobaan 60 detik untuk mendorong burung bergerak ke atau menjauh dari tempat makan (yaitu, membuat pilihan). Untuk burung chickadee yang secara acak ditetapkan ke Benar 1 (ID panggilan) atau Benar 2 (ID lagu), semua stimulus yang direkam dari Jantan A dan Betina A ditetapkan ke S+, sementara stimulus yang direkam dari Jantan B dan Betina B ditetapkan ke S−. Untuk burung chickadee yang secara acak ditetapkan ke Pseudo 1 (kontrol panggilan) atau Pseudo 2 (kontrol lagu), setengah dari setiap stimulus yang direkam burung stimulus ditetapkan secara acak semu sebagai S+ atau S−. Karena setiap burung stimulus menyumbangkan stimulus S+ dan S− dalam kelompok diskriminasi semu, burung chickadee tidak dapat menerapkan ID dengan andal untuk menyelesaikan tugas diskriminasi, mereka juga tidak dapat mengandalkan fitur identitas lain seperti jenis kelamin atau spesies, dan dengan demikian terbatas pada penggunaan hafalan (Herrnstein 1990 ). Burung chickadee diharuskan untuk mencapai rasio diskriminasi (DR), yang mewakili proporsi semua percobaan Go yang benar dari semua percobaan Go (Go S+ / (Go S+ dan Go S−)) minimal 0,80 untuk minimal enam blok percobaan, dengan dua blok terakhir berurutan, di mana blok percobaan terdiri dari enam penyajian setiap stimulus. Ke-44 stimulus dimainkan dalam urutan acak tanpa penggantian. Setelah burung chickadee mempelajari tugas tersebut, pembelajaran mereka diuji menggunakan stimulus baru.
1.7 Tahap Pengujian Pertama
Tahap pengujian terdiri dari dua pengujian untuk generalisasi, pertama pengujian probe, dan kemudian pengujian transfer penuh. Selama pengujian probe, empat stimulus baru dari masing-masing dari empat burung stimulus ditambahkan ke kumpulan stimulus. Ke-16 stimulus baru ini memiliki tipe vokalisasi yang sama seperti yang disajikan dalam tahap diskriminasi awal (panggilan untuk True 1 dan Pseudo 1, lagu untuk True 2 dan Pseudo 2). Untuk kelompok True 1 dan True 2, stimulus yang direkam dari Jantan A dan Betina A diberi label P +, dan yang direkam dari Jantan B dan Betina B diberi label P−. Label ini diacak semu untuk kelompok Pseudo 1 dan Pseudo 2. Namun, tidak seperti stimulus S+ dan S−, stimulus probe tidak diperkuat atau dihukum, terlepas dari bagaimana burung merespons, untuk menguji generalisasi pembelajaran tanpa penguatan diferensial. Karena stimulus probe tidak diberi imbalan maupun tidak diberi imbalan, dan untuk mencegah berkurangnya motivasi untuk menanggapi stimulus probe, burung chickadee diharuskan menyelesaikan dua blok percobaan berturut-turut dengan rasio diskriminasi minimal 0,80 sambil mempertahankan parameter diskriminasi awal, tetapi dengan probabilitas imbalan yang berkurang selama percobaan S+ dari 100% menjadi 85%. Dengan melakukan hal itu, burung chickadee tidak akan terlalu bias terhadap tanggapan setelah pemberian stimulus probe yang tidak diberi imbalan dan tidak akan kehilangan motivasi karena berkurangnya probabilitas imbalan selama pengujian probe. Setiap burung chickadee menyelesaikan 6 blok percobaan pengujian probe, di mana blok percobaan terdiri dari setiap stimulus probe yang dimainkan satu kali, dan setiap stimulus S+ dan S− dimainkan dua kali (untuk mempertahankan proporsi stimulus probe sekitar 15%).
Setelah pengujian probe, stimulus P + dan P− diubah menjadi stimulus S+ dan S− secara berurutan, dan tujuh stimulus baru tambahan (panggilan untuk True 1 dan Pseudo 1, lagu untuk True 2 dan Pseudo 2) dari setiap burung stimulus (total 88 stimulus) juga ditambahkan ke kumpulan stimulus, untuk uji transfer penuh. Selama pengujian ini, probabilitas hadiah selama uji coba S+ Go dikembalikan ke 100%, dan respons chickadee terhadap stimulus baru dibandingkan dengan stimulus yang dilatih. Chickadee lulus uji transfer ini setelah mereka mencapai rasio diskriminasi minimal 0,80 untuk minimal enam blok uji coba, dua blok terakhir harus berurutan. Seperti pada fase diskriminasi awal, blok uji coba terdiri dari enam penyajian setiap stimulus.
1.8 Tahap Pengujian Kedua
Seperti pada fase pengujian pertama, burung chickadee pertama kali diuji dengan probabilitas hadiah yang dikurangi untuk uji coba S+ Go dari 100% menjadi 85%. Setelah burung chickadee mencapai dua blok uji coba berturut-turut dengan rasio diskriminasi minimal 0,80, empat stimulus probe baru dari masing-masing dari empat burung stimulus diperkenalkan ke kumpulan stimulus. Stimulus probe yang direkam dari Jantan A dan Betina A diberi label sebagai P+, dan yang direkam dari Jantan B dan Betina B diberi label P−. Meskipun stimulus S+ dan S− diperkuat atau dihukum tergantung pada respons burung chickadee, stimulus probe tidak diberi hadiah maupun hukuman. Kali ini, stimulus S+ dan S− hanya diputar sekali di setiap blok uji coba untuk mempertahankan proporsi stimulus probe sebesar 15%. Berbeda dengan uji coba pertama, 16 stimulus uji coba baru ini adalah jenis vokalisasi yang sebelumnya tidak disajikan dalam pelatihan diskriminasi awal dan fase pengujian pertama (lagu untuk True 1 dan Pseudo 1, panggilan untuk True 2 dan Pseudo 2). Burung chickadee menyelesaikan enam blok uji coba, di mana setiap stimulus disajikan sekali per blok uji coba.
Untuk tahap akhir penelitian, burung chickadee memasuki transfer penuh terakhir, di mana 16 stimulus probe diubah dari P+ dan P− menjadi S+ dan S− masing-masing, dan tujuh stimulus tambahan dari setiap burung stimulus, juga dari jenis vokalisasi yang tidak disajikan selama pelatihan diskriminasi awal (lagu untuk True 1 dan Pseudo 1, panggilan untuk True 2 dan Pseudo) juga ditambahkan. Probabilitas penghargaan selama uji coba S+ Go dikembalikan lagi ke 100%, dan respons burung chickadee terhadap stimulus baru sekali lagi dibandingkan dengan stimulus yang dilatih. Burung chickadee lulus uji transfer penuh ini setelah mereka menunjukkan rasio diskriminasi minimal 0,80 untuk minimal 6 blok uji coba, 2 terakhir di antaranya berurutan, di mana blok uji coba terdiri dari enam penyajian setiap stimulus.
1.8.1 Kriteria Gagal
Bahasa Indonesia: Agar tidak memperpanjang waktu burung percobaan di ruang operan, dan untuk memastikan bahwa kami menyediakan waktu yang cukup untuk menilai apakah burung akan mempelajari suatu tugas, kami menggunakan kriteria gagal spesifik yang ditetapkan untuk setiap tahap pelatihan. Dalam studi Montenegro et al. ( 2020 ), burung chickadee yang dilatih untuk menghafal lagu fee-bee individu (kelompok kontrol semu yang mirip dengan yang dijelaskan di atas) membutuhkan waktu kurang dari 18.000 kali percobaan rata-rata untuk mencapai kriteria lulus selama pelatihan diskriminasi awal, dan kurang dari 14.400 kali percobaan untuk mencapai kriteria lulus selama uji transfer generalisasi. Berdasarkan nilai-nilai ini dan memungkinkan percobaan tambahan untuk kemungkinan pembelajaran yang lebih lambat dalam kelompok panggilan chick-a-dee , burung dalam studi kami diharuskan untuk mencapai DR awal = 0,80 dalam 100 blok percobaan (26.400 percobaan), dan mencapai semua kriteria lulus dalam 115 blok percobaan (30.360 percobaan). Selama tahap pelatihan transfer pertama, burung dinyatakan gagal jika tidak mencapai DR = 0,80 dalam 50 blok percobaan (26.400 percobaan), dan semua lulus kriteria dalam 65 blok percobaan (34.320 percobaan). Pada tahap pelatihan akhir, burung dipensiunkan jika gagal mencapai DR = 0,80 dalam 33 blok percobaan (26.136 percobaan) dan lulus dalam 41 blok percobaan (32.472 percobaan). Selain itu, burung dipensiunkan dari penelitian setelah 100 hari di dalam bilik. Sebagian besar burung menyelesaikan penelitian jauh sebelum periode batas waktu ini, dengan rata-rata 67,2 hari dalam penelitian (SD = 18,66).
Sebanyak lima burung kelompok Pseudo mencapai kriteria gagal (yaitu, dikeluarkan dari penelitian sebelum mencapai kinerja tingkat kriteria). Dari jumlah tersebut, dua burung dari Pseudo 1 mencapai 100 hari dalam penelitian antara dua fase pengujian dan dipensiunkan lebih awal. Tiga burung tambahan dari Pseudo 2 gagal mencapai DR = 0,80 dalam 100 blok percobaan selama tahap diskriminasi awal dan kemudian dipensiunkan. Untuk tiga burung terakhir ini, tempat penampung 100 blok percobaan untuk awalnya mencapai DR = 0,80 dan 115 untuk menyelesaikan pelatihan diskriminasi masing-masing digunakan untuk tujuan analisis. Karena lima dari delapan burung Pseudo total gagal selama, atau segera setelah pelatihan diskriminasi awal, dan ini menunjukkan hubungan signifikan antara kondisi (Benar vs. Pseudo) dan hasil (ꭓ 2 (1) = 6,94, p = 0,008), kami memilih untuk mengecualikan kelompok Pseudo dari semua analisis berikutnya kecuali yang membandingkan jumlah blok percobaan yang diperlukan untuk mencapai DR = 0,80 dan lulus pelatihan diskriminasi awal (di mana kedelapan burung Pseudo dapat disertakan dalam analisis) di antara kelompok. Kami percaya bahwa pengecualian kelompok kontrol ini dapat dibenarkan karena kami dapat secara wajar berasumsi bahwa karena lima burung kelompok Pseudo gagal, sementara tidak ada burung kelompok Benar yang gagal, tugas menghafal kelompok Pseudo jauh lebih sulit daripada pembelajaran diskriminasi kelompok Benar. Namun, statistik ringkasan untuk burung-burung ini disertakan (lihat Tabel 1 ). Selain itu, selama pelatihan diskriminasi awal, satu burung dari Benar 2 membutuhkan waktu jauh lebih lama daripada burung lain di kedua kelompok Benar untuk mempelajari tugas tersebut. Kami menemukan bahwa ruang operan ini mengalami malfungsi audio selama beberapa uji coba. Karena malfungsi tersebut menyebabkan burung tersebut menjadi outlier yang ekstrem, burung ini juga dikeluarkan dari semua analisis.
| Pelatihan diskriminasi | |||
|---|---|---|---|
| N | M | SD | |
| Benar 1 | 6 | 0,65 | 0.11 |
| Benar 2 | 5 | 0.62 | 0,07 |
| Semu 1 | 4 | 0.51 | 0,01 |
| Semu 2 | 4 | 0,50 | 0,01 |
| Tes Transfer Pertama | |||
|---|---|---|---|
| Benar 1 | 6 | 0,93 | 0,03 |
| Benar 2 | 5 | 0,91 | 0,05 |
| Semu 1 | 4 | 0.51 | 0,03 |
| Semu 2 | 1 | 0.62 | Bahasa Indonesia |
| Tes Transfer Kedua | |||
|---|---|---|---|
| Benar 1 | 6 | 0.53 | 0.13 |
| Benar 2 | 5 | 0.48 | 0.22 |
| Semu 1 | 2 | 0.49 | 0,06 |
| Semu 2 | 1 | 0.49 | Bahasa Indonesia |
Catatan: Rasio diskriminasi menunjukkan proporsi respons pendekatan pengumpan (Go) yang benar dari semua respons Go. Ukuran sampel berkurang jika burung telah mencapai kriteria gagal pada tahap sebelumnya.
2 Analisis Statistik
Semua analisis statistik dilakukan menggunakan R v4.3.1 (Tim Inti R 2023 ). Kami melakukan Analisis Varians (ANOVA) model campuran menggunakan paket afex (Singmann et al. 2023 ) untuk membandingkan DR rata-rata untuk stimulus baru dan yang telah dilatih sebelumnya dalam enam blok percobaan pertama dari dua uji transfer dalam dan di antara kelompok. Kami juga menggunakan ANOVA model campuran untuk membandingkan proporsi respons Go terhadap stimulus S+, S−, P + dan P− di antara kelompok dan di seluruh uji probe. Paket afex secara otomatis menerapkan koreksi sferisitas Greenhouse–Geisser jika sferisitas dilanggar. Jika efek utama dan interaksi yang signifikan ditemukan, perbandingan post hoc Tukey dilakukan menggunakan paket emmeans (Lenth 2023 ). Normalitas dinilai menggunakan uji Shapiro–Wilk dan plot ggplot2 qq (Kassambara 2023 ; Wickham 2016 ). Jika normalitas menjadi perhatian (membandingkan jumlah blok uji coba dengan kriteria pada tahap diskriminasi awal), uji jumlah peringkat Kruskal-Wallis diikuti oleh uji jumlah peringkat wilcoxon post hoc dengan koreksi Benjamini-Hochberg digunakan sebagai pengganti ANOVA. Homogenitas varians dinilai menggunakan uji Levene. Dalam semua kasus, batas alfa p = 0,05 digunakan untuk menandai signifikansi statistik. Angka hasil dibuat dengan paket ggplot2 (Kassambara 2023 ).
3 Hasil
3.1 Pelatihan Diskriminasi
Selama pelatihan diskriminasi awal, burung kelompok True dilatih untuk membedakan di antara 44 stimulus berdasarkan diskriminasi individu, sementara burung kelompok Pseudo dilatih untuk membedakan di antara 44 stimulus menggunakan hafalan. Rata-rata, burung dalam True 1 mengambil M = 8,17 blok percobaan (SD = 4,75) untuk mencapai DR = 0,80; True 2 M = 7,6 blok percobaan (SD = 1,67); Pseudo 1 M = 36,8 blok percobaan (SD = 24,6); dan Pseudo 2 M = 87,5 blok percobaan (SD = 25; lihat Gambar 3 ). Ada perbedaan signifikan antar kelompok dalam jumlah blok percobaan yang diperlukan untuk mencapai DR = 0,80 (ꭓ 2 (3) = 13,627, p = 0,003). Perbandingan berpasangan post hoc menunjukkan bahwa burung dalam setiap kelompok True mengambil blok percobaan yang jauh lebih sedikit daripada kelompok Pseudo (semua p = 0,037). Tidak ada perbedaan signifikan antara kelompok True ( p = 0,713) atau antara kelompok Pseudo ( p = 0,065).
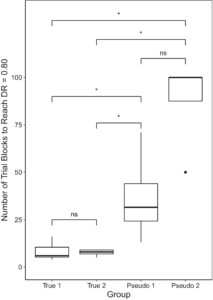
Burung dalam kelompok True juga menempuh lebih sedikit blok percobaan untuk mencapai kriteria lulus, dengan burung True 1 menempuh rata-rata 15,5 blok percobaan (SD = 9,29), burung True 2 M = 13,4 blok percobaan (SD = 1,34), burung Pseudo 1 M = 46,5 blok percobaan (SD = 29,6) dan burung Pseudo 2 M = 102 blok percobaan (SD = 26,5). Ada juga perbedaan signifikan antar kelompok dalam jumlah blok percobaan yang diperlukan untuk lulus tahap ini (ꭓ 2 (3) = 13,313, p = 0,004). Burung True 1 menempuh blok percobaan yang jauh lebih sedikit daripada burung Pseudo 2 ( p = 0,038), tetapi tidak dari burung Pseudo 1 ( p = 0,063). Burung True 2 menempuh blok percobaan yang jauh lebih sedikit daripada burung di kedua kelompok Pseudo ( ps = 0,038). Baik burung kelompok sejati ( p = 0,408) maupun burung kelompok semu ( p = 0,065) tidak berbeda secara signifikan satu sama lain.
3.2 Tahap Pengujian
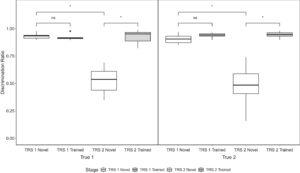
Selama uji coba, kami membandingkan proporsi rata-rata respons Go burung terhadap stimulus S+, S−, P+, dan P−. Jika burung dalam kelompok True telah mempelajari tugas diskriminasi individual, kami akan memperkirakan mereka akan mendekati tempat makan dengan tingkat yang lebih tinggi setelah presentasi P + daripada setelah presentasi P−. Rata-rata, selama uji coba pertama, burung True 1 merespons dengan memasuki tempat makan (Go) pada 81,2% percobaan S+, 9,1% percobaan S−, 74% percobaan P +, dan 8,2% percobaan P−. Burung True 2 menunjukkan respons Go terhadap 91,8% percobaan S+, 27,5% percobaan S−, 95,9% percobaan P +, dan 30,5% percobaan P−. Selama uji coba kedua, burung True 1 merespons dengan memasuki tempat makan (Go) pada 84,3% percobaan S+, 7,6% percobaan S−, 28,2% percobaan P +, dan 54% percobaan P−. Burung True 2 menunjukkan respons Go pada 88,2% percobaan S+, 5,1% percobaan S−, 10,9% percobaan P +, dan 15% percobaan P−. Terdapat pengaruh utama yang signifikan dari uji probe (pertama atau kedua; F (1,9) = 13,534, p = 0,005, η 2 G = 0,228) dan jenis stimulus (S+, S−, P + atau P−; F (2,56,23) = 147,952, p < 0,001, η 2 G = 0,792), dan interaksi yang signifikan dari faktor-faktor ini ( F (1,81,16,33) = 44,240, p < 0,001, η 2 G = 0,523) pada proporsi respons Go. Tidak terdapat pengaruh signifikan dari keanggotaan kelompok ( F (1,9) = 0,147, p = 0,710, η 2 G = 0,006). Pada pemeriksaan pertama, burung True 1 menunjukkan respons Go yang signifikan lebih banyak terhadap percobaan S+ dibandingkan dengan percobaan S− ( t (9) = 7.371, p < 0.001) dan terhadap percobaan P + dibandingkan dengan percobaan P− ( t (9) = 8.205, p < 0.001). Demikian pula, selama uji pemeriksaan pertama, burung True 2 memberikan respons lebih banyak terhadap percobaan S+ dibandingkan dengan percobaan S− ( t (9) = 5.931, p = 0.003) dan terhadap percobaan P + dibandingkan dengan percobaan P− ( t (9) = 7.475, p < 0.001). Namun, selama uji coba kedua, burung True 1 hanya merespons secara signifikan lebih banyak pada percobaan S+ daripada S− ( t (9) = 20.203, p < 0,001), dan benar-benar merespons secara signifikan lebih banyak pada percobaan P− daripada pada percobaan P + ( t (9) = 4.067, p = 0,037). Pada uji coba kedua, burung True 2 juga merespons secara signifikan lebih banyak pada percobaan S+ daripada pada percobaan S− ( t(9) = 19.966, p < 0,001), namun tidak memberikan respon yang signifikan terhadap percobaan P + atau P− ( t (9) = 0,589, p = 0,998).
Selama pengujian transfer penuh, kami membandingkan DR rata-rata burung untuk stimulus baru dan stimulus terlatih dalam enam blok percobaan pertama dari setiap pengujian transfer (lihat Gambar 4 ). Jika burung Sejati mempelajari tugas diskriminasi individu, mereka harus mampu menggeneralisasi pembelajaran ini dalam pengujian transfer pertama ke stimulus baru, sebagaimana dibuktikan oleh DR yang tinggi dan kira-kira sama untuk stimulus baru seperti untuk stimulus terlatih. Dalam pengujian transfer pertama, burung Sejati 1 memiliki DR rata-rata 0,921 (SD = 0,029) untuk stimulus terlatih dan 0,931 (SD = 0,028) untuk stimulus baru. Burung Sejati 2 memiliki DR rata-rata 0,937 (SD = 0,026) untuk stimulus terlatih dan 0,905 (SD = 0,046) untuk stimulus baru dalam pengujian transfer pertama. Jika burung True mampu menggeneralisasi diskriminasi individu yang dipelajari di seluruh jenis vokalisasi, maka kita akan mengharapkan burung menunjukkan DR yang tinggi dan kira-kira sama untuk stimulus baru seperti untuk stimulus yang dilatih dalam uji transfer kedua. Selama uji transfer kedua ini, burung True 1 memiliki DR rata-rata 0,925 (SD = 0,065) untuk stimulus yang dilatih dan 0,525 (SD = 0,129) untuk stimulus baru. Burung True 2 memiliki DR rata-rata 0,942 (SD = 0,031) untuk stimulus yang dilatih dan 0,475 (SD = 0,217) untuk stimulus baru. Secara keseluruhan, terdapat efek signifikan dari fase uji (pertama atau kedua; F (1,9) = 69,796, p < 0,001, η 2 G = 0,595), dan jenis stimulus (baru atau terlatih; F (1,9) = 55,286, p < 0,001, η 2 G = 0,629), dan interaksi signifikan dari faktor-faktor ini ( F (1,9) = 40,342, p < 0,001, η 2 G = 0,606) pada DR rata-rata. Tidak terdapat efek signifikan keanggotaan kelompok pada DR rata-rata ( F (1,9) = 0,174, p = 0,686, η 2 G = 0,004). Dalam uji transfer pertama, burung True 1 tidak secara signifikan lebih baik dalam membedakan stimulus terlatih daripada stimulus baru ( t (9) = 1,106, p = 0,695). Hal yang sama ditemukan untuk burung True 2 ( t (9) = 3,030, p = 0,057). Namun, dalam uji transfer kedua, baik burung True 1 ( t (9) = 4,718, p = 0,005) dan True 2 ( t (9) = 5,032, p = 0,003) menunjukkan diskriminasi stimulus baru yang secara signifikan lebih buruk daripada stimulus terlatih. Baik burung True 1 ( t (9) = 5,289, p = 0,002) dan burung True 2 ( t (9) = 5,113, p = 0,003) secara signifikan lebih buruk dalam membedakan stimulus baru dalam uji transfer kedua daripada yang pertama.
4 Diskusi
Studi kami bertujuan untuk mengonfirmasi apakah burung chickadee bertopi hitam dapat menggunakan panggilan chick-a-dee untuk diskriminasi individual dan menjawab apakah identifikasi yang dipelajari dapat digeneralisasi dari panggilan chick-a-dee ke lagu fee-bee dan sebaliknya. Kami melatih dua kelompok burung chickadee untuk membedakan di antara vokalisasi empat individu, menggunakan panggilan atau lagu sebagai stimulus diskriminatif (kelompok diskriminasi Sejati), dan membandingkan perolehan pembelajaran dalam kelompok Sejati dengan dua kelompok kontrol di mana burung dilatih menggunakan hafalan dari stimulus yang sama (kelompok diskriminasi Semu). Selanjutnya kami menguji generalisasi di seluruh jenis vokalisasi dengan menghadirkan burung chickadee dengan stimulus yang direkam dari burung yang sama tetapi kali ini dari jenis vokalisasi yang berlawanan dengan yang digunakan selama pelatihan diskriminasi awal. Burung chickadee dalam kelompok Sejati mempelajari tugas diskriminasi secara signifikan lebih cepat daripada mereka yang berada dalam kelompok kontrol Semu, yang menunjukkan aturan diskriminasi yang digunakan burung kelompok Sejati memberi burung-burung ini keuntungan substansial dibandingkan dengan pembelajaran melalui hafalan. Faktanya, ID memberikan keuntungan seperti itu sehingga sementara total lima burung Pseudo gagal dalam penelitian, tiga di antaranya gagal selama tahap diskriminasi awal, tidak ada burung Sejati (bahkan satu burung yang pembelajarannya terganggu oleh kegagalan peralatan) gagal menyelesaikan seluruh penelitian. Burung dalam Pseudo 2 tampaknya sangat dirugikan, khususnya selama tahap diskriminasi awal. Ini mungkin mencerminkan kesulitan yang lebih besar dalam membedakan antara iterasi lagu fee-bee yang lebih stereotip (Ficken et al. 1978 ) daripada iterasi panggilan chick-a-dee yang lebih bervariasi yang disajikan pada burung Pseudo 1. Kami mengakui bahwa ukuran sampel kami terbatas. Namun, ketika perbedaan yang signifikan secara statistik terdeteksi, ukuran efek yang menyertainya secara konsisten besar. Sementara penelitian masa depan dengan ukuran sampel yang lebih besar akan diterima, temuan yang telah kami laporkan di sini didukung oleh ukuran efeknya yang besar.
Chickadee dalam kelompok True mampu menggeneralisasi pembelajaran mereka ke stimulus baru dengan tipe vokalisasi yang sama, tanpa pengurangan signifikan dalam DR ke stimulus baru vs. stimulus terlatih. Dengan demikian, kami mampu menunjukkan bahwa chickadee dapat menggunakan ID menggunakan panggilan chick-a-dee dalam paradigma pengkondisian operan, dan juga mampu mereplikasi temuan Montenegro et al. ( 2020 ) tentang diskriminasi individu menggunakan lagu fee-bee . Namun, ketika disajikan dengan stimulus dari tipe vokalisasi yang berlawanan (misalnya, lagu fee-bee untuk panggilan chick-a-dee ), chickadee tidak dapat menggeneralisasi pembelajaran mereka. Ada penurunan signifikan dalam DR untuk stimulus baru ini dibandingkan dengan stimulus yang dilatih sebelumnya. Kedua kelompok True merespons stimulus ini pada peluang yang hampir sama (DR = 0,50), yang menunjukkan tidak ada keuntungan bagi burung True berdasarkan pembelajaran sebelumnya untuk membedakan stimulus baru ini. Meskipun kami tidak menyertakan burung kelompok Pseudo dalam analisis fase pengujian generalisasi apa pun karena tingkat kegagalannya sangat tinggi, burung yang menyelesaikan fase pengujian kedua juga, seperti yang dapat diduga, merespons rangsangan baru ini secara kebetulan.
Menariknya, meskipun chickadee dalam kelompok True tidak dapat menggeneralisasi ID yang dipelajari dengan sukses di seluruh jenis vokalisasi, burung True 1 sekitar 2,5 kali lebih mungkin merespons nyanyian betina baru selama uji coba kedua daripada nyanyian jantan baru. Karena jenis kelamin diseimbangkan antara kategori stimulus yang diberi hadiah dan tidak diberi hadiah (satu jantan dan satu betina masing-masing diberi hadiah dan dihukum), dan setiap bias yang sudah ada sebelumnya dikendalikan selama pra-pelatihan, kami tidak akan mengantisipasi perbedaan substansial dalam menanggapi satu jenis kelamin daripada yang lain. Perbedaan seperti itu juga tidak diamati pada burung True 2, yang menanggapi panggilan chick-a-dee jantan dan betina baru secara setara. Kami pertama-tama berasumsi bahwa perilaku yang diamati dalam burung True 1 ini mungkin merupakan fungsi dari jantan (yang merupakan empat dari enam burung di True 1) yang menanggapi pasangan pengembangbiakan potensial. Namun, keempat jantan itu sendiri hanya sekitar dua kali lebih mungkin menanggapi betina daripada jantan, sementara dua betina sedikit lebih dari tiga kali lebih mungkin menanggapi betina daripada jantan. Mengingat preferensi yang tinggi untuk lagu betina dibanding lagu jantan, kami malah mengandaikan bahwa bias ini mencerminkan peningkatan kemauan untuk mendekat saat penyanyi cenderung menjadi individu yang kurang dominan, dan kurang mengancam, karena burung chickadee bertopi hitam betina cenderung tunduk pada jantan (Desrochers 1989 ; Odum 1941 ), dan jantan cenderung memainkan peran yang lebih aktif dalam pertahanan teritorial, sering kali didahului dengan bernyanyi (Baker et al. 2012 ; Stefanski 1967 ). Penulis studi masa depan yang menerapkan lagu jantan dan betina sebagai stimulus harus berhati-hati saat menafsirkan bias yang dimediasi jenis kelamin, terutama yang menyangkut tugas kognitif di mana kinerja yang berubah dapat dikaitkan dengan perolehan pembelajaran yang terhambat untuk satu jenis kelamin dibandingkan yang lain.
Kurangnya generalisasi lintas-vokalisasi yang diamati dalam penelitian kami meninggalkan kita dengan dua pertanyaan utama. Pertama, mengapa burung chickadee tidak dapat menggeneralisasi ID yang dipelajari di antara tipe vokalisasi? Di permukaannya, kurangnya generalisasi pengenalan individu di antara tipe vokalisasi tampaknya merupakan sistem yang tidak efisien, meskipun ini bukan temuan yang unik. Elie dan Theunissen ( 2018 ) menemukan bahwa burung pipit zebra ( Taeniopygia guttata ) dapat membedakan antara individu menggunakan setiap tipe panggilan dalam repertoar mereka, tetapi burung pipit zebra kemungkinan menghafal penanda tanda tangan individu dari setiap tipe panggilan secara independen. Demikian pula, burung pipit lagu yang dikondisikan secara operan untuk membedakan antara individu tidak dapat menggeneralisasi pembelajaran mereka di antara tipe lagu (Beecher et al. 1994 ). Elie dan Theunissen ( 2018 ) mengusulkan bahwa sementara menghafal tanda tangan vokal untuk setiap tipe vokalisasi secara independen adalah sistem yang kurang efisien, itu kemungkinan merupakan hasil dari penyaringan frekuensi pasif yang terbatas. Dengan kata lain, anatomi burung kecil seperti zebra finch, dan mungkin chickadee, sangat terbatas dalam kapasitasnya untuk variasi antar individu sehingga hanya ada sedikit peluang bagi frekuensi resonansi untuk digunakan secara andal sebagai isyarat identitas lintas-vokalisasi. Untuk mendukung teori Elie dan Theunissen ( 2018 ) mengenai penyaringan frekuensi pasif terbatas pada spesies burung kecil, bukti terkini untuk generalisasi identitas lintas-vokalisasi, atau pemodelan generalisasi tersebut, mudah ditemukan pada mamalia (Cheney dan Seyfarth 1988 ; Pisanski et al. 2020 ; Reby et al. 2006 ), tetapi tidak pada burung. Hafalan tanda tangan individu spesifik vokalisasi akan menjelaskan ketidakmampuan chickadee dalam penelitian kami untuk menggeneralisasi ID yang dipelajari di seluruh panggilan dan lagu. Akan tetapi, hal ini membawa kita kepada pertanyaan kedua: jika burung chickadee menghafal tanda vokal individu secara independen untuk setiap jenis vokalisasi, lalu bagaimana burung chickadee memahami bahwa tanda vokalisasi spesifik ini berasal dari produsen yang sama?
Bisa jadi burung chickadee yang terbang bebas mendekati seekor individu setiap kali jenis vokalisasi baru dihasilkan, mengidentifikasi individu tersebut secara visual sebelum menambahkan tanda vokal baru yang dihafal ini ke “kotak peralatan IVR” mereka. Namun, strategi ini bisa memakan waktu, dan akan menghadirkan risiko yang lebih besar daripada mengidentifikasi potensi ancaman dari jauh. Hipotesis lain adalah bahwa burung chickadee mengandalkan penggunaan isyarat eksternal seperti kedekatan spasial dan temporal, atau penanda identitas vokal lainnya, seperti jenis kelamin atau dominasi, untuk menghubungkan vokalisasi yang berbeda bersama-sama. Logika teori ini adalah bahwa ketika dua vokalisasi dari jenis yang berbeda berasal dari tempat yang sama pada waktu yang sama dan dari individu yang sama-sama dominan dengan jenis kelamin yang sama, mereka dapat secara wajar dikaitkan dengan individu yang sama. Atau, individu dapat mengandalkan informasi kontekstual yang dikodekan untuk secara bersamaan memperoleh makna dan identitas dari sinyal akustik.
Cheney dan Seyfarth ( 1988 ) menemukan bahwa monyet vervet ( Chlorocebus pygerythrus ) mampu menggeneralisasikan habituasi yang dipelajari untuk memutar ulang panggilan Wrr individu tertentu ke panggilan Chutter individu yang sama (dua panggilan yang menyampaikan informasi kontekstual yang sama dan memunculkan respons perilaku yang sama), tetapi tidak menggeneralisasikan habituasi individu yang dipelajari ketika panggilan yang terbiasa dan panggilan baru menyampaikan informasi kontekstual yang berbeda (seperti panggilan alarm macan tutul dan panggilan alarm elang yang memunculkan respons perilaku yang sangat berbeda). Cheney dan Seyfarth ( 1988 ) juga dapat menunjukkan bahwa informasi identitas individu diperhatikan selama paradigma ini, karena generalisasi tidak terjadi ketika panggilan yang terbiasa diikuti oleh stimulus yang dihasilkan oleh individu baru, terlepas dari apakah kedua jenis panggilan tersebut berbagi atau tidak berbagi makna kontekstual. Tampaknya, monyet vervet dapat menggunakan generalisasi lintas-vokalisasi dari pengenalan individu, tetapi kemampuan ini mungkin terbatas pada vokalisasi yang menyampaikan informasi kontekstual yang sama. ( 2006 ) menemukan bahwa auman umum rusa merah ( Cervus elaphus ) dapat digunakan untuk melatih model Markov tersembunyi untuk mengkategorikan auman berdasarkan individu. Ketika model tersebut kemudian diuji dengan auman keras , gonggongan kejaran , dan gonggongan (vokalisasi dengan penggunaan kontekstual yang tumpang tindih) yang direkam dari individu yang sama, model tersebut mampu mengidentifikasi produser dengan benar sebanyak 63,4% dari waktu tersebut. Auman umum rusa merah sebagian besar digunakan selama penggembalaan betina dan kontes jantan sejenis, auman keras pada akhir kontes auman atau selama penggembalaan, gonggongan kejaran dalam konteks perilaku agonistik dan berkembang biak/menggembalakan sesama spesies, dan gonggongan dapat mendahului auman, dapat menandakan ancaman di dekatnya, atau dapat digunakan dalam konteks teritorial atau perkawinan (Reby dan McComb 2003 ). Demikian pula, analisis akustik Pisanski et al. ( 2020 ) terhadap rekaman manusia menemukan bahwa frekuensi fundamental dapat digunakan untuk memprediksi identitas dalam vokalisasi non-verbal individu manusia. Kekuatan korelasi ini paling besar antara vokalisasi dengan penggunaan kontekstual yang serupa, seperti ucapan dan teriakan yang menakutkan, atau ucapan dan raungan agresif.
Bahasa Indonesia : Dalam mempertimbangkan Reby dan McComb ( 2003 ) dan Pisanski et al. ( 2020 ), sementara berbagai vokalisasi yang dijelaskan mungkin memiliki tujuan yang sedikit berbeda, mereka dapat terjadi secara bersamaan dan berbagi tumpang tindih kontekstual yang substansial. Dalam kasus ini, bagaimanapun, vokalisasi juga lebih mirip secara akustik satu sama lain, yang mungkin lebih mudah mengakomodasi tanda vokal bersama daripada yang diberikan oleh vokalisasi yang digunakan dalam penelitian kami. Kami menyajikan chickadee dengan panggilan chick-a-dee dan lagu fee-bee , dua vokalisasi yang, meskipun terkadang diproduksi dalam suksesi pendek, berfungsi untuk menyampaikan informasi kontekstual yang sangat berbeda dan sangat berbeda secara akustik (Ficken et al. 1978 ; Smith 1991 , 57–74). Jika kami menggunakan dua vokalisasi yang lebih mirip secara kontekstual, seperti panggilan gargle , yang digunakan dalam konteks teritorial dan agonistik (Smith 1991 , 57), dan lagu fee-bee , kami mungkin telah melihat generalisasi yang lebih besar. Jika burung chickadee benar-benar memperhatikan makna kontekstual dari sinyal akustik seperti halnya mereka memperhatikan identitas penghasil sinyal, seperti yang dijelaskan pada monyet vervet oleh Cheney dan Seyfarth ( 1988 ), mungkin informasi kontekstual ini membantu memfasilitasi asimilasi tanda vokal khusus vokalisasi oleh penerima sinyal. Hipotesis semacam itu memerlukan penyelidikan lebih lanjut untuk lebih memahami cara pengenalan vokal individu diperoleh, disimpan, dimodifikasi, dan diambil pada burung chickadee dan burung penyanyi lainnya.


