
Abstrak
Spesies Gracilariaceae termasuk rumput laut yang paling penting secara ekonomi, dengan berbagai macam aplikasi. Budidaya Gracilariopsis lemaneiformis skala besar telah ditetapkan di sepanjang pantai timur Cina. Namun, karena permintaan global untuk spesies Gracilariaceae meningkat, sangat penting bagi Cina untuk memperluas budidaya spesies ini dalam skala besar ke wilayah pesisir lainnya. Meskipun Laut Cina Selatan menyimpan sumber daya makroalga yang kaya, termasuk Gracilariaceae, budidaya spesies ini dalam skala besar belum dilaksanakan di wilayah tersebut. Untuk mengidentifikasi spesies Gracilariaceae yang toleran panas yang cocok untuk budidaya skala besar di Laut Cina Selatan, kami melakukan survei antara Juni dan Agustus dari tahun 2021 hingga 2024. Beberapa spesies yang toleran panas telah diidentifikasi. Identitas taksonomi dikonfirmasi melalui pemeriksaan morfologi terperinci dan analisis molekuler dari urutan gen rbc L dan cox1 . Kami mengidentifikasi delapan spesies Gracilariaceae, termasuk satu spesies Gracilariopsis ( Gp .) dan tujuh spesies Gracilaria ( G .). Secara morfologi, spesies tersebut dicirikan sebagai berikut: Gp. heteroclada (cabang sekunder panjang dengan cabang tersier pendek atau berduri), G. salicornia (segmen berbentuk gada), G. vermiculophylla dan G. tenuistipitata (bercabang ramping), G. edulis (tampilan seperti sapu dengan pangkal cabang berwarna merah), G. firma (pangkal cabang sangat menyempit), dan dua spesies yang tidak terklasifikasi, Gracilaria sp.1 dan Gracilaria sp.2. Studi ini memberikan kontribusi terhadap pengetahuan taksonomi Gracilariaceae dan memberikan informasi berharga untuk pengembangan spesies toleran panas yang cocok untuk budidaya skala besar di Laut Cina Selatan.
1. PENDAHULUAN
Famili Gracilariaceae, pertama kali dideskripsikan oleh Nägeli pada tahun 1847, termasuk dalam filum Rhodophyta (alga merah) dalam kelas Florideophyceae dan ordo Gracilariales (Guiry, 2024 ). Spesies ini sebagian besar menghuni lingkungan air hangat di seluruh laut tropis, subtropis, dan beriklim sedang, dan sering kali ditemukan di zona pasang surut dan subpasang surut di wilayah Atlantik barat dan Indo-Pasifik (Norris, 2014 ; Xia & Zhang, 1999 ). Gracilariaceae memiliki beragam aplikasi dalam makanan (Chan & Matanjun, 2017 ; Tabarsa et al., 2012 ), obat-obatan (Naveena & Prakash, 2013 ), akuakultur (Chaitanawisuti et al., 2011 ; Samocha et al., 2015 ), dan pertanian (Singh et al., 2016 ; Torres et al., 2018 ) karena profil nutrisinya yang kaya, termasuk kadar protein, mineral, vitamin, dan serat makanan yang tinggi. Secara historis di Tiongkok, alga ini tidak hanya dihargai sebagai makanan lezat tetapi juga digunakan sebagai bahan pengikat kapur untuk lukisan dinding tradisional karena aksesibilitas dan kemudahan pemrosesannya (Tabarsa et al., 2012 ). Yang terpenting, Gracilariaceae berfungsi sebagai bahan baku penting untuk memproduksi agar bermutu pangan, yang banyak digunakan sebagai bahan tambahan pangan, bahan alami berbasis bio, dan agen penstabil dalam perlengkapan medis (Armisen, 1995 ; Lee et al., 2022 ; Li & Liu, 2022 ; Sousa et al., 2010 ). Lebih jauh lagi, famili alga merah ini telah digunakan sebagai biostimulan dalam pertanian, sehingga meningkatkan hasil panen dan meningkatkan keuntungan (Bhushan et al., 2023 ). Singkatnya, karena aplikasinya yang luas, Gracilariaceae telah muncul sebagai salah satu alga yang paling signifikan secara ekonomi secara global.
Gracilaria (disebut sebagai G. dalam nama binomial) dan Gracilariopsis (disebut sebagai Gp . dalam nama binomial) adalah genera yang paling umum, signifikan secara ekonomi, dan digunakan secara luas dalam famili Gracilariaceae. Menurut literatur, ada 206 spesies Gracilaria dan 23 spesies Gracilariopsis , yang secara kolektif mencakup 94,24% spesies dalam famili Gracilariaceae (Guiry, 2024 ). Gp. lemaneiformis dibudidayakan secara luas di sepanjang pantai timur Tiongkok, menjadikannya spesies budidaya terbesar dari famili Gracilariaceae di wilayah tersebut. Pada tahun 2017, area budidaya untuk Gp. lemaneiformis mencakup lebih dari 8000 hektar, menghasilkan lebih dari 300.000 ton rumput laut kering (Jiang et al., 2019 ). Budidaya ekstensif ini menjadikan Tiongkok sebagai produsen agar terbesar di dunia (Porse & Rudolph, 2017 ). Meskipun Tiongkok telah mencapai keberhasilan penting dalam budidaya spesies Gracilariaceae, masih ada tantangan yang menghambat peningkatan lebih lanjut dalam produksinya. Misalnya, spesies Gracilariaceae yang dibudidayakan di Tiongkok kurang beragam. Gp. lemaneiformis sebagian besar dibudidayakan dalam skala besar, sedangkan G. vermiculophylla , G. firma , G. blodgettii , dan G. tenuistipitata dibudidayakan dalam skala kecil dan spesies Gracilariaceae lainnya hampir tidak dibudidayakan (Wang et al., 2020 ; Yang et al., 2015 ). Dalam akuakultur, budidaya satu spesies dapat dengan mudah menyebabkan ketidakseimbangan unsur dalam lingkungan, mengganggu ekosistem, dan memicu penyakit, yang pada gilirannya, mengurangi hasil keseluruhan (Dong & Gao, 2023 ; Laktuka et al., 2023 ). Selain itu, suhu pertumbuhan optimal untuk Gp. lemaneiformis berkisar antara 12 dan 26°C (Zhang & Fei, 2008 ), yang membuatnya tidak cocok untuk budidaya skala besar di perairan yang lebih hangat di wilayah pesisir selatan Tiongkok. Mengembangkan spesies yang tahan panas akan menjembatani kesenjangan dalam budidaya Gracilariaceae skala besar di wilayah ini dan selanjutnya meningkatkan produksi Gracilariaceae Tiongkok secara keseluruhan.
Laut Cina Selatan kaya akan sumber daya makroalga dan menjadi rumah bagi berbagai spesies Gracilariaceae (Li et al., 2020 ; Nguyen et al., 2023 ; Xia & Zhang, 1999 ). Akan tetapi, budidaya spesies Gracilariaceae dalam skala besar di wilayah ini masih belum berkembang. Kelompok penelitian kami telah melakukan beberapa survei terhadap sumber daya makroalga di wilayah pesisir ini dan menemukan bahwa spesies Gracilariaceae paling melimpah dari bulan November hingga bulan April berikutnya. Selama bulan-bulan musim panas (dari bulan Juni hingga bulan Agustus), suhu air bisa sangat tinggi, terkadang melebihi 35°C, sehingga menyulitkan kelangsungan hidup beberapa spesies, termasuk Gp. lemaneiformis . Beberapa spesies Gracilariaceae yang mampu bertahan hidup pada suhu tinggi telah diidentifikasi selama survei yang dilakukan antara Juni dan Agustus 2021 dan 2024. Untuk memastikan identitas taksonomi mereka, kami memeriksa dan mencatat secara menyeluruh karakteristik morfologi spesies ini dan memperoleh 26 subunit I sitokrom oksidase ( cox1 ) dan 30 ribulosa-1,5-bifosfat karboksilase oksigenase ( rbc L) dari sampel. Studi ini memberikan kontribusi terhadap pengetahuan taksonomi spesies Gracilariaceae di Laut Cina Selatan dan menjadi dasar bagi pengembangan spesies Gracilariaceae yang toleran terhadap panas yang cocok untuk budidaya skala besar di wilayah tersebut.
2 BAHAN DAN METODE
2.1 Koleksi sampel Gracilariaceae
Sampel spesies Gracilariaceae yang digunakan dalam penelitian ini dikumpulkan dari Beihai (BH, 21°27′33″ LU, 109°14′20″ BT) di Daerah Otonomi Guangxi Zhuang; Wushi (WS, 20°29′57″ LU, 109°50′3″ BT), Sanjiao (SJ, 20°33′51″ LU, 109°50′11″ BT), Haiwei (HW, 20°26′20″ LU, 109°57′10″ BT), Diaoluo (DL, 21°7′57″ LU, 110°21′58″ BT), Pulau Techeng (TCI, 21°9′9″ LU, 110°26′39″ BT), Pulau Donghai (DHI, 21°1′48″ LU, 110°32′57″ BT) di Provinsi Guangdong; Niuhe (NH, 19°24′14″ N, 110°41′5″ E) di Provinsi Hainan, Tiongkok (Gambar 1 ), antara Juli dan Agustus 2021 dan 2024. Sampel kemudian diangkut ke laboratorium dalam pendinginan pada suhu 4°C. Setelah itu, sampel dibersihkan dengan air laut yang disterilkan untuk menghilangkan kotoran permukaan dan dibudidayakan sementara di ruang kultur alga. Kondisi budidaya dipertahankan pada suhu 25°C dengan intensitas cahaya 70 μmol m −2 s −1 , salinitas 30, dan pH 8,0. Air laut diganti setiap minggu, dengan penambahan natrium nitrat (143 μmol/L) dan kalium dihidrogen fosfat (6,45 μmol/L) setelah setiap penggantian air.

2.2 Pengamatan morfologi
Sampel dengan perbedaan morfologi yang jelas dipilih dari ruang kultur alga untuk analisis morfologi yang terperinci. Fitur morfologi eksternal, termasuk warna talus, tekstur, bentuk keseluruhan, pola percabangan, dan ada atau tidaknya penyempitan pada pangkal cabang, difoto dan didokumentasikan menggunakan kamera digital (Canon EOS M6; Canon, Tokyo, Jepang). Selain itu, potongan cabang talus utama dibuat dengan tangan, dan struktur internal, seperti ukuran dan bentuk sel kortikal dan medula, difoto dan didokumentasikan menggunakan mikroskop cahaya (Olympus CX33; Olympus Imaging Corp., Tokyo, Jepang). Setiap organ reproduksi yang diamati juga dicatat.
2.3 Ekstraksi DNA dan amplifikasi PCR
Total DNA diekstraksi dari sampel menggunakan DNeasy Plant Mini Kit (Qiagen, Valencia, CA, AS). Konsentrasi cetakan DNA disesuaikan menjadi 50 ng/μL untuk amplifikasi PCR sekuens cox1 dan rbc L. Primer untuk amplifikasi sekuens cox1 dan rbc L tercantum dalam Tabel 1. Sistem reaksi PCR terdiri dari 20 μL, termasuk 1 μL cetakan DNA, masing-masing 1 μL primer positif dan negatif, 10 μL campuran PCR 2 × San Taq, dan 7 μL ddH 2 O. Prosedur amplifikasi PCR adalah sebagai berikut: denaturasi awal pada 94°C selama 5 menit; 28 siklus denaturasi pada 94°C selama 30 detik, annealing pada 58°C selama 30 detik, ekstensi pada 72°C selama 1 menit; dan perpanjangan akhir pada suhu 72°C selama 10 menit. Produk PCR disimpan pada suhu 4°C. Pengurutan produk yang diperkuat dilakukan oleh Shanghai Majorbio Bio-Pharm Technology Co., Ltd., dan urutan yang dirakit disediakan.
| Nama gen | Bahasa Inggris: Primer | Urutan | Referensi |
|---|---|---|---|
| cox1 | GWSFn | 5′ GTATACATATGATGGHGCTCAA 3′ | Saunders ( 2008 ) |
| Bahasa Indonesia: CoxIR1 | 5′ GTATACATATGATGGHGCTCAA 3′ | ||
| rbc L | 57-F | 5′ GTAATTCCATATGCTAAAATGGG 3′ | Air Tawar dan Rueness ( 1994 ) |
| 1381-R | 5′ ATCTTCCATAGATCTAAAGC 3′ |
2.4 Analisis sekuens cox1 dan rbc L
Urutan gen cox1 dan rbc L yang baru diperkuat (Tabel 2 ) dibandingkan dengan urutan terkait dalam GenBank (Tabel S1 dan S2 ) menggunakan alat BLAST di NCBI. Berdasarkan literatur yang ada, urutan keyakinan tinggi dipilih dan diunduh. Analisis fitur urutan dilakukan pada urutan yang diunduh dan yang baru diperkuat menggunakan MEGA v11.0.13 (Tamura et al., 2004 ). Model evolusi optimal untuk dataset molekuler dipilih menggunakan Modeltest v3.7 (Posada & Buckley, 2004 ). Sebanyak 55 urutan digunakan untuk membangun pohon filogenetik cox1 , dengan 29 urutan bersumber dari GenBank dan 26 urutan yang baru diperkuat. Untuk pohon filogenetik rbc L, total 58 urutan digunakan, dengan 28 urutan diperoleh dari GenBank dan 30 urutan yang baru diperkuat. Rhodymenia pseudopalmata digunakan sebagai outgroup untuk kedua pohon filogenetik. Pohon filogenetik dibangun menggunakan metode inferensi Bayesian (BI), kemungkinan maksimum (ML), dan neighbor-joining (NJ). MrBayes v3.1.2 (Ronquist & Huelsenbeck, 2003 ) digunakan untuk membangun pohon BI, dengan waktu proses 1.000.000 generasi, frekuensi pengambilan sampel 1000 generasi, dan membuang 25% pohon pertama. Pohon konsensus aturan mayoritas 50% dihitung dari pohon yang tersisa. MEGA v11.0.13 digunakan untuk membangun pohon ML dan NJ. Model GRT + G + I digunakan untuk pohon ML, dan model dua parameter Kimura diterapkan untuk pohon NJ dengan 1000 replikasi bootstrap. Jarak genetik antara sekuens dihitung menggunakan model dua parameter Kimura di MEGA v11.0.13.
| Kode regangan | Lokasi pengumpulan | Tanggal pengambilan | Nomor akses cox1 | Nomor akses rbc L |
|---|---|---|---|---|
| BH01 | Beihai, Guangxi, Tiongkok | 26-Agustus-2024 | PQ467722 | PQ438118 |
| BH02 | Beihai, Guangxi, Tiongkok | 26-Agustus-2024 | PQ467723 | / |
| DHI01 | Pulau Donghai, Guangdong, Cina | 15 Juli 2024 | / | PQ438119 |
| DHI02 | Pulau Donghai, Guangdong, Cina | 15 Juli 2024 | / | PQ438120 |
| Nomor DL01 | Diaoluo, Guangdong, Tiongkok | 17 Juli 2024 | / | PQ438121 |
| HW01 | Haiwei, Guangdong, Tiongkok | 10 Juli 2024 | / | PQ438122 |
| HW02 | Haiwei, Guangdong, Tiongkok | 10 Juli 2024 | PQ467721 | PQ438123 |
| HW03 | Haiwei, Guangdong, Tiongkok | 10 Juli 2024 | / | PQ438124 |
| HW04 | Haiwei, Guangdong, Tiongkok | 10 Juli 2024 | PQ467724 | PQ438125 |
| HW05 | Haiwei, Guangdong, Tiongkok | 10 Juli 2024 | PQ467725 | PQ438126 |
| HW06 | Haiwei, Guangdong, Tiongkok | 10 Juli 2024 | PQ467720 | PQ438127 |
| NH01 | Niuhe, Hainan, Tiongkok | 08 Juli 2021 | PQ467730 | PQ438128 |
| NH02 | Niuhe, Hainan, Tiongkok | 08 Juli 2021 | PQ467731 | PQ438129 |
| NH03 | Niuhe, Hainan, Tiongkok | 08 Juli 2021 | PQ467732 | PQ438130 |
| NH04 | Niuhe, Hainan, Tiongkok | 08 Juli 2021 | PQ467733 | PQ438131 |
| SJ01 | Sanjia, Guangdong, Tiongkok | 22 Juli 2021 | PQ467726 | PQ438132 |
| SJ02 | Sanjia, Guangdong, Tiongkok | 22 Juli 2021 | PQ467727 | / |
| SJ03 | Sanjia, Guangdong, Tiongkok | 22 Juli 2021 | PQ467728 | / |
| SJ04 | Sanjia, Guangdong, Tiongkok | 22 Juli 2021 | PQ467729 | / |
| SJ05 | Sanjia, Guangdong, Tiongkok | 22 Juli 2021 | / | PQ438133 |
| SJ06 | Sanjia, Guangdong, Tiongkok | 22 Juli 2021 | / | PQ438134 |
| SJ07 | Sanjia, Guangdong, Tiongkok | 18 Juli 2024 | / | PQ438135 |
| SJ08 | Sanjia, Guangdong, Tiongkok | 18 Juli 2024 | / | PQ438136 |
| Nomor WS01 | Wushi, Guangdong, Tiongkok | 20 Juli 2021 | PQ467734 | PQ438137 |
| Nomor WS02 | Wushi, Guangdong, Tiongkok | 20 Juli 2021 | PQ467735 | PQ438138 |
| Nomor WS03 | Wushi, Guangdong, Tiongkok | 20 Juli 2021 | PQ467736 | PQ438139 |
| Nomor WS04 | Wushi, Guangdong, Tiongkok | 20 Juli 2021 | / | PQ438140 |
| WS05 | Wushi, Guangdong, Tiongkok | 20 Juli 2021 | / | PQ438141 |
| Tanggal WS06 | Wushi, Guangdong, Tiongkok | 20 Juli 2021 | PQ467737 | PQ438142 |
| Tanggal WS07 | Wushi, Guangdong, Tiongkok | 20 Juli 2024 | PQ467738 | PQ438143 |
| Tanggal WS08 | Wushi, Guangdong, Tiongkok | 20 Agustus 2021 | PQ467739 | PQ438144 |
| Tanggal WS09 | Wushi, Guangdong, Tiongkok | 20 Agustus 2021 | PQ467740 | / |
| Bahasa Inggris WS10 | Wushi, Guangdong, Tiongkok | 20 Agustus 2021 | PQ467741 | / |
| Bahasa Inggris WS11 | Wushi, Guangdong, Tiongkok | 20 Agustus 2021 | PQ467742 | / |
| Bahasa Indonesia: WS12 | Wushi, Guangdong, Tiongkok | 20 Agustus 2021 | PQ467743 | / |
| Bahasa Indonesia: WS13 | Wushi, Guangdong, Tiongkok | 20 Agustus 2021 | PQ467744 | PQ438145 |
| Minggu ke 14 | Wushi, Guangdong, Tiongkok | 20 Agustus 2021 | / | PQ438146 |
| WS15 | Wushi, Guangdong, Tiongkok | 20 Agustus 2021 | PQ467745 | PQ438147 |
Catatan : “/” berarti data tidak tersedia.
3 HASIL
3.1 Morfologi Strain Gp. Heteroclada
Thalli bersifat soliter atau berkelompok, tegak, linier, silindris, dan terdapat dalam jumlah besar di kolam yang tenang atau saluran masuk yang bergerak lambat. Thalli tingginya 15–70 cm, dengan beberapa yang tingginya lebih dari 100 cm. Thalli berdaging, selalu berair, dan ditambatkan oleh holdfast berbentuk cakram, meskipun mudah patah (Gambar 2a ). Sumbu utama kadang-kadang diatapi, lebarnya berkisar antara 1 hingga 4 mm, dan memiliki satu hingga empat ordo percabangan, terutama lateral atau bergantian dengan cabang dikotomis sporadis. Cabang sekunder memanjang, sedangkan cabang tersier pendek, terkadang seperti duri, dan lebarnya berkisar antara 0,2 hingga 0,8 mm. Pangkal cabang sedikit menyempit, dengan ujung yang menyempit secara progresif (Gambar 2b ). Warna thalli sangat bervariasi tergantung pada lingkungannya: spesimen pasang surut berwarna hijau zamrud atau coklat muda; spesimen saluran masuk berwarna coklat kemerahan; dan spesimen kolam sebagian besar berwarna hijau tua. Spesimen herbarium melekat sepenuhnya pada kertas pemasangan.

Talus dibedakan menjadi medula dan korteks. Medula terdiri dari sel-sel besar, hampir bulat, berdinding tipis yang berdiameter 350–1150 μm dan tebal 5–10 μm. Lapisan gelatin, yang tebalnya sekitar 2–3 μm, menutupi permukaan talus. Korteks luar mengandung sel-sel kaya plastida, membentuk satu hingga dua lapisan yang berdiameter 6–10 μm, sedangkan lapisan korteks dalam memiliki dua hingga tiga sel yang berdiameter 15–50 μm. Batas antara korteks dan medula jelas (Gambar 2c,d ).
Sistokarp berbentuk subkonis, menonjol dengan sedikit penyempitan di pangkalnya, dan tidak berparuh, dengan tinggi 550–1100 μm dan lebar 900–2000 μm. Dinding sistokarp tipis (ketebalan 70–110 μm) dan terdiri dari 7–10 lapisan sel. Karposporangia di dalam sistokarp berdiameter 20–35 μm; tidak ada filamen transfer yang diamati antara sistokarp dan filamen sporofit (Gambar 2e ). Tetrasporangia, yang tersebar di permukaan talus, berwarna cokelat pucat, oval pada penampang melintang, dan berdiameter 15–25 μm (Gambar 2f ).
Nomor Sampel: HW01, dikumpulkan oleh Zhaojun Zeng pada 10 Juli 2024, dari Haiwei, Leizhou, Provinsi Guangdong. NH01-04, dikumpulkan oleh Jianjun Cui dan Enyi Xie pada 8 Juli 2021, dari Niuhe, Wenchang, Provinsi Hainan. SJ05, dikumpulkan oleh Rong Xin pada 22 Juli 2021, dari Sanjiao, Leizhou, Provinsi Guangdong. WS01-05, dikumpulkan oleh Youyou Guo dan Chunli Chen pada 20 Juli 2021, dari Wushi, Leizhou, Provinsi Guangdong. Spesimen diawetkan di Laboratorium Makroalga Universitas Kelautan Guangdong.
3.2 Morfologi strain G. salicornia
Talus tegak, baik soliter atau berkelompok, silindris, dan umumnya berukuran tinggi 5–20 cm, dengan spesimen tumbuh di kolam yang tingginya melebihi 25 cm. Talus ditambatkan oleh holdfast berbentuk cakram. Talus tersusun dari ruas berbentuk tongkat, meruncing dari bagian atas yang lebih tebal ke bagian bawah yang lebih tipis, dengan diameter berkisar antara 1,5 hingga 3,5 mm. Bagian atas talus sering kali memiliki beberapa cabang kecil yang tersusun dalam pola seperti payung (Gambar 3a ). Talus berwarna coklat kekuningan atau kuning gading, berdaging dan segar, dengan talus segar sangat rapuh dan sulit untuk diawetkan secara utuh. Spesimen herbarium yang telah disiapkan tidak sepenuhnya menempel pada kertas pemasangan.

Medula terdiri dari sel-sel besar, bersudut, berdinding tipis dengan diameter 350–550 μm dan ketebalan 5 μm. Sel-sel kortikal hampir berbentuk bulat dengan diameter berkisar antara 6 hingga 50 μm, termasuk 1–2 lapisan sel kortikal luar berpigmen dan 1–3 lapisan sel kortikal dalam. Lapisan gelatin yang tebalnya sekitar 5–7 μm menutupi permukaan talus (Gambar 3b,c ).
Cystocarpus hampir berbentuk bulat dengan diameter 800–1400 μm, menonjol dengan paruh, dan pangkalnya sedikit menyempit atau tidak menyempit. Carposporangia hampir berbentuk bulat dengan diameter 25–30 μm. Pericarpus tebal (180–250 μm) dan tersusun atas 12–18 lapisan sel (Gambar 3d ). Spermatangia menghuni sel-sel kortikal dengan rapat dan rongga berbentuk oval dengan diameter rongga 30–40 μm, yang diklasifikasikan sebagai “tipe V” (Gambar 3e ).
Nomor Sampel: HW06, dikumpulkan oleh Zhaojun Zeng pada 10 Juli 2024, dari Haiwei, Leizhou, Provinsi Guangdong. SJ07, dikumpulkan oleh Zhaojun Zeng pada 18 Juli 2024, dari Sanjiao, Leizhou, Provinsi Guangdong. WS13-15, dikumpulkan oleh Kun Lin dan Rong Xin pada 20 Juli 2021, dari Wushi, Leizhou, Provinsi Guangdong. Spesimen diawetkan di Laboratorium Makroalga Universitas Kelautan Guangdong.
3.3 Morfologi dua spesies Gracilaria dengan cabang ramping
3.3.1 Galur G. vermiculophylla
Talus tegak, soliter atau berkelompok, linier, dan silindris, memiliki tinggi berkisar antara 20 hingga 35 cm, dan holdfast berbentuk cakram kecil di pangkalnya (Gambar 4a ). Sumbu utama kadang-kadang diatapi, dan lebarnya berkisar antara 0,8 hingga 2,0 mm. Cabang-cabang sekunder relatif panjang, menghasilkan cabang-cabang pendek yang tidak teratur bergantian, lateral, atau dikotomi dengan penyempitan yang terlihat di pangkal cabang (Gambar 4b ). Talus berwarna coklat, teksturnya lembut, dan spesimen herbarium yang disiapkan menempel pada kertas pemasangan.

Medula terdiri dari sel-sel besar, berbentuk tidak teratur, dan bulat, berdinding tipis dengan diameter 160–300 μm dan ketebalan 8–10 μm. Korteks terdiri dari 3–6 lapisan sel yang ukurannya bertambah dari lapisan luar ke lapisan dalam, dengan 1–2 lapisan terluar mengandung plastida. Batas antara korteks dan medula tidak jelas dan lapisan gelatin yang menutupi permukaan talus setebal 6–10 μm (Gambar 4c,d ).
Cystocarpus hampir berbentuk bulat, menonjol dengan jelas, tidak berparuh dengan pangkal yang sedikit menyempit, dan memiliki diameter 500–700 μm. Karposporangia berbentuk bulat dan memiliki diameter 15–20 μm. Perikarpnya tipis dan terdiri dari 6–12 lapisan sel dengan ketebalan 100–200 μm; tidak ada filamen transfer yang diamati (Gambar 4e ). Tetrasporangia berwarna cokelat pucat, menunjukkan pembelahan berbentuk salib, dan tersebar di antara sel-sel kortikal, yang tampak berbentuk kerucut atau oval memanjang pada penampang melintang, dengan tinggi 25–40 μm dan lebar 15–25 μm (Gambar 4f ).
Nomor Sampel: DL01-03, dikumpulkan oleh Zhaojun Zeng pada tanggal 17 Juli 2024, dari Diaoluo, Zhanjiang, Provinsi Guangdong. Spesimen tersebut diawetkan di Laboratorium Makroalga Universitas Kelautan Guangdong.
3.3.2 strain G. tenuistipitata
Talus ramping, soliter atau berkelompok, linier, dan silindris, dengan tinggi berkisar antara 15 hingga 35 cm, dan tidak ada holdfast yang diamati (Gambar 4g ). Sumbu utama, yang memiliki diameter 0,2–0,5 mm, tidak mencolok, dengan banyak cabang halus dan tersusun rapat yang tidak meruncing di pangkalnya. Banyak cabang kecil dan rapuh terdapat pada cabang sekunder (Gambar 4h ). Talus berwarna cokelat muda dan bertekstur rapuh. Spesimen herbarium yang telah disiapkan menempel pada kertas pemasangan. Medula terdiri dari sel-sel yang hampir bulat dengan diameter 150–450 μm dan ketebalan 5–10 μm. Sel-sel kortikal luar mengandung plastida, membentuk 1–2 lapisan, hampir bulat, dan memiliki diameter 5–10 μm. Sel-sel kortikal bagian dalam terdiri dari 2–3 lapisan dengan diameter berkisar antara 20 hingga 70 μm. Lapisan gelatin yang menutupi permukaan talus memiliki ketebalan 5–8 μm (Gambar 4i,j ).
Nomor Sampel: BH01-02, dikumpulkan oleh Zhaojun Zeng pada tanggal 26 Agustus 2024, dari Beihai, Daerah Otonomi Guangxi Zhuang. HW04-05, dikumpulkan oleh Zhaojun Zeng pada tanggal 10 Juli 2024, dari Haiwei, Leizhou, Provinsi Guangdong. Spesimen tersebut diawetkan di Laboratorium Makroalga Universitas Kelautan Guangdong.
3.4 Morfologi dua spesies Gracilaria dengan penampakan corymbose
3.4.1 Galur G. edulis
Talus tegak dan berkelompok, silindris, memiliki stolon menjalar dan holdfast berbentuk cakram di pangkalnya, dan tingginya berkisar antara 10 hingga 20 cm. Sumbu utama hanya terlihat jelas di bagian bawah dan memiliki diameter 1,5–2,0 mm. Percabangan terjadi lima hingga enam kali dan sebagian besar bergantian atau dikotomi, dengan lebih sedikit contoh percabangan lateral atau trikotomi yang diamati. Percabangan biasanya dimulai di bagian tengah talus, sehingga membuat seluruh struktur tampak seperti sapu atau umbellate (Gambar 5a ). Pangkal cabang tidak meruncing atau sedikit meruncing dan menunjukkan warna merah yang jelas; cabang-cabang kecil memiliki diameter 0,5–0,8 mm dengan ujung yang tajam, sering kali bercabang (Gambar 5b ). Talus berwarna coklat teh, bertulang rawan, dan spesimen herbarium yang disiapkan tidak sepenuhnya menempel pada kertas pemasangan.

Medula tersusun atas sel-sel besar, hampir bulat, berdinding tipis dengan diameter 180–320 μm dan tebal 5–6 μm. Sel-sel kortikal berpigmen membentuk 2–3 lapisan dan berdiameter 3,0–4,5 μm. Batas antara korteks dan medula jelas, dan lapisan gelatin yang menutupi permukaan talus setebal 2–3 μm (Gambar 5c,d ). Sistokarp memiliki tinggi 700–1200 μm dan lebar 12–1800 μm, hampir bulat, menonjol, sedikit berparuh dengan pangkal sedikit menyempit, dan sebagian besar terdistribusi di bagian atas talus; karposporangia berbentuk bulat dengan diameter 20–30 μm. Perikarp tersusun atas 9–13 lapisan sel dan tebalnya 200–300 μm; filamen transfer terlihat di dasar sistokarp (Gambar 5e ).
Nomor Sampel: DHI01-02, dikumpulkan oleh Zhaojun Zeng pada tanggal 15 Juli 2024, dari Pulau Donghai, Zhanjiang, Provinsi Guangdong. SJ08, dikumpulkan oleh Zhaojun Zeng pada tanggal 18 Juli 2024, dari Sanjiao, Leizhou, Provinsi Guangdong. Spesimen tersebut diawetkan di Laboratorium Makroalga Universitas Kelautan Guangdong.
3.4.2 Strain Gracilaria sp.1
Thalli tegak, silindris, baik soliter atau berkelompok dengan tinggi berkisar antara 10 hingga 18 cm, dan ditambatkan oleh holdfast berbentuk cakram. Sumbu utama hanya menonjol di bagian bawah dan memiliki diameter 1–2 mm. Percabangan sebagian besar bergantian tidak teratur, dengan pangkal cabang sedikit meruncing atau tidak meruncing. Setengah bagian atas talus dicirikan oleh banyak cabang padat dengan ujung pendek dan tajam, yang memberikan kontur umbellate secara keseluruhan (Gambar 5f ). Warna thalli bervariasi dari coklat tua dan coklat muda hingga hijau zamrud, dengan spesimen herbarium yang disiapkan menempel pada kertas pemasangan.
Medula terdiri dari sel-sel besar, hampir bulat, berdinding tipis dengan diameter 160–500 μm dan ketebalan 3,5–6,0 μm. Lapisan kortikal luar mengandung sel-sel kaya plastida yang membentuk 2–3 lapisan dan memiliki diameter 5–9 μm; lapisan kortikal dalam memiliki 1–2 lapisan dan diameter 10–50 μm. Batas antara korteks dan medula jelas, dengan lapisan gelatin setebal 4–7 μm yang menutupi permukaan talus (Gambar 5g,h ).
Cystocarpus hampir bulat, menonjol, dengan tinggi 700–1200 μm dan lebar 1300–1800 μm, tanpa paruh, dan sedikit menyempit di pangkalnya. Karposporangia berbentuk bulat atau oval, dengan diameter 20–30 μm. Perikarp terdiri dari 7–10 lapisan sel dan tebalnya 120–180 μm; tidak ada filamen transfer yang diamati antara perikarp dan filamen sporofit (Gambar 5i ). Tetrasporangia berwarna cokelat, menunjukkan pembelahan berbentuk salib, berbentuk oval pada penampang melintang, dengan tinggi 25–30 μm dan lebar 9–15 μm, dan sel-sel kortikal di sekitarnya dimodifikasi (Gambar 5j ).
Nomor Sampel: SJ01-04, dikumpulkan oleh Chunli Chen pada tanggal 22 Juli 2021, dari Sanjiao, Leizhou, Provinsi Guangdong. Spesimen tersebut diawetkan di Laboratorium Makroalga Universitas Kelautan Guangdong.
3.5 Morfologi dua spesies Gracilaria dengan pangkal cabang yang sangat menyempit
3.5.1 Strain G. firma
Thalli tegak, silindris, tebal, baik soliter atau berkelompok, dengan tinggi berkisar antara 10 hingga 25 cm. Sumbu utamanya tidak mencolok, memiliki lebar 1,5–3,0 mm, dan pangkalnya ditambatkan oleh holdfast berbentuk cakram (Gambar 6a ). Percabangan sebagian besar bergantian, dengan pangkal cabang sering kali sangat menyempit, sehingga membentuk tangkai kecil (Gambar 6b ). Ujung cabang secara bertahap meruncing. Talus berwarna hijau tua atau hijau zamrud dengan tekstur keras, dan spesimen herbarium yang disiapkan tidak sepenuhnya menempel pada kertas pemasangan.
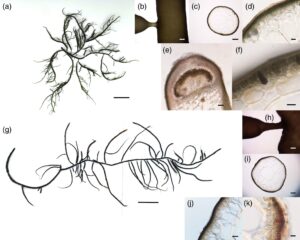
Medula terdiri dari sel-sel besar berbentuk bulat tidak beraturan dengan diameter 300–600 μm dan ketebalan 10–15 μm. Korteks luar terdiri dari 1–3 lapisan sel yang mengandung plastida dan berdiameter 3–6 μm, sedangkan korteks dalam terdiri dari 3–5 lapisan sel dan berdiameter 20–40 μm. Batas antara korteks dan medula jelas, dan lapisan gelatin yang menutupi permukaan talus setebal 5–7 μm (Gambar 6c,d ).
Cystocarpus berbentuk setengah bola atau kerucut, menonjol, dengan pangkal yang tidak menyempit atau sedikit menyempit dan tidak berparuh, dan memiliki tinggi 500–1000 μm dan lebar 600–1500 μm. Karposporangia berbentuk bulat dengan diameter 20–30 μm. Perikarpus tersusun atas 9–13 lapisan sel dengan ketebalan 100–180 μm; tidak ada filamen transfer yang diamati (Gambar 6e ). Tetrasporangia berwarna cokelat, tersebar di antara sel-sel kortikal, tampak oval atau kerucut pada penampang melintang, memiliki tinggi 20–40 μm dan lebar 10–30 μm, dan sel-sel kortikal di sekitarnya termodifikasi (Gambar 6f ).
Nomor Sampel: HW02-03, dikumpulkan oleh Zhaojun Zeng dan Wencheng Yang pada tanggal 10 Juli 2024, dari Haiwei, Leizhou, Provinsi Guangdong. SJ06, dikumpulkan oleh Rong Xin pada tanggal 22 Juli 2021, dari Sanjiao, Leizhou, Provinsi Guangdong. WS06-12, dikumpulkan oleh Kun Lin pada tanggal 20 Juli 2021, dari Wushi, Leizhou, Provinsi Guangdong. Spesimen diawetkan di Laboratorium Makroalga Universitas Kelautan Guangdong.
3.5.2 Strain Gracilaria sp.2
Thalli tegak, silindris, baik soliter atau berkelompok, dan relatif besar, biasanya berkisar antara 20 hingga 50 cm tingginya, dengan spesimen terpanjang melebihi 60 cm. Sumbu utama tidak mencolok, memiliki lebar 1-3 mm, dan holdfast berbentuk cakram di pangkalnya (Gambar 6g ). Cabang-cabangnya panjang, dengan cabang terpanjang berukuran 51 cm, dan tersusun tidak teratur, bergantian, atau lateral dengan percabangan dikotomis sesekali. Pangkal cabang seringkali sangat menyempit, sehingga membentuk tangkai kecil (Gambar 6h ) dan ujung cabang secara bertahap meruncing, dengan sebagian besar patah menjadi bentuk terpotong tidak teratur, di mana cabang-cabang kecil dapat muncul dari tepi yang patah. Talus berwarna merah tua atau coklat kemerahan, berdaging, dan menjadi lunak setelah dikeringkan. Spesimen herbarium yang telah disiapkan menempel pada kertas pemasangan.
Medula tersusun atas sel-sel besar, berbentuk tidak teratur, berdinding tipis dengan diameter 350–700 μm dan ketebalan 3,5–5,5 μm. Medula di bagian bawah talus sering pecah dan berongga. Korteks terdiri atas 2–4 lapisan sel dan berdiameter 5–9 μm, dengan 1–2 lapisan terluar mengandung plastida. Batas antara korteks dan medula jelas, dan lapisan gelatin yang menutupi permukaan talus setebal 4,5–6,5 μm. Permukaan talus memiliki berbagai kotoran dan alga epifit (Gambar 6i,j ). Tetrasporangia mengalami pembelahan krusiatum, berwarna merah lembayung, dan terdistribusi padat di dalam sel-sel kortikal talus. Tetrasporangia tampak berbentuk oval memanjang pada penampang melintang, dengan tinggi 20–30 μm dan lebar 13–18 μm, dan sel-sel kortikal di sekitarnya mengalami modifikasi (Gambar 6k ).
Nomor Sampel: TCI01-03, dikumpulkan oleh Zhaojun Zeng pada tanggal 5 Agustus 2024, dari Pulau Techeng, Zhanjiang, Provinsi Guangdong. Spesimen tersebut diawetkan di Laboratorium Makroalga Universitas Kelautan Guangdong.
3.6 Penilaian molekuler cox1
Panjang sekuens cox1 terakhir adalah 446 bp, dengan 275 situs yang dilestarikan, 171 situs variabel, dan 144 situs yang informatif tentang parsimoni. Pohon filogenetik dibangun menggunakan metode BI, ML, dan NJ. Struktur topologi yang diperoleh dari ketiga metode tersebut sebagian besar konsisten; oleh karena itu, pohon filogenetik yang dibangun menggunakan metode ML dipilih untuk dipresentasikan (Gambar 7 ).
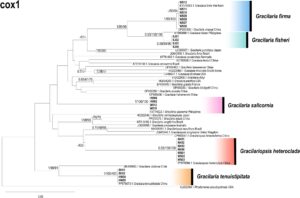
Menurut pohon filogenetik cox1 , 26 sekuens yang diperkuat dalam penelitian ini membentuk lima klade. Klade pertama terdiri dari sampel WS06–12 dan G. firma dari Vietnam (KY315253.1), yang dikelompokkan bersama, tanpa perbedaan sekuens basa dan jarak genetik 0 yang diamati. Sekuens ini juga dikelompokkan dengan G. changii dari Tiongkok (OP669398.1) dengan dukungan bootstrap tinggi (1/99/100) dan jarak genetik di antara keduanya adalah 0,225%. Klade kedua terdiri dari sampel SJ01–04, yang dikelompokkan bersama tanpa perbedaan sekuens basa dan jarak genetik 0 yang diamati. Sekuens ini juga dikelompokkan dengan G. fisheri dari Filipina (KY995690.1) dengan dukungan bootstrap tinggi (0,93/100/100) dan jarak genetik di antara keduanya adalah 1,358%. Klade ketiga terdiri dari sampel HW02, HW06, WS13, WS15, dan G. salicornia dari Filipina (KY774622.1), yang dikelompokkan bersama, dengan dukungan bootstrap tinggi (1/100/100), tidak ada perbedaan urutan basa, dan jarak genetik 0 yang diamati. Klade keempat terdiri dari sampel WS01–03, NH01–04, dan Gp. heteroclada dari Tiongkok (PP579596.1), yang dikelompokkan bersama, dengan dukungan bootstrap tinggi (0,92/100/100), tidak ada perbedaan urutan basa, dan jarak genetik 0 yang diamati. Klade kelima terdiri dari sampel BH01–02, HW04–05, dan G. tenuistipitata dari Cina (PP579573.1), yang dikelompokkan bersama, dengan dukungan bootstrap yang tinggi (1/100/100), tidak ada perbedaan urutan basa, dan jarak genetik 0 yang diamati.
3.7 Penilaian molekular rbc L
Panjang sekuens rbc L akhir adalah 1132 bp, dengan 730 situs yang dilestarikan, 401 situs variabel, dan 328 situs yang informatif tentang parsimoni. Pohon filogenetik dibangun menggunakan metode BI, ML, dan NJ. Struktur topologi yang diperoleh dari ketiga metode tersebut sebagian besar konsisten; oleh karena itu, pohon filogenetik yang dibangun menggunakan metode ML dipilih untuk dipresentasikan (Gambar 8 ).
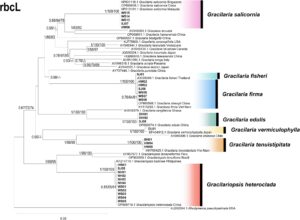
Menurut pohon filogenetik rbc L, 30 sekuens sampel yang dianalisis dalam penelitian ini membentuk tujuh klade. Klade pertama terdiri dari sampel HW06, SJ07, WS13–15, dan sekuens G. salicornia dari Singapura (KF831118.1), Tiongkok (OP669616.1), dan Malaysia (KP010101.1), yang dikelompokkan bersama dengan dukungan bootstrap tinggi (1/100/100). Tidak ada perbedaan sekuens basa yang diamati di antara mereka, dan jarak genetiknya adalah 0. Klade kedua terdiri dari sampel SJ01 dan G. fisheri dari Thailand (JQ026049.1), yang juga dikelompokkan bersama tanpa perbedaan sekuens dan dengan jarak genetik 0 yang diamati. Klade ketiga terdiri dari sampel HW02–03, SJ06, WS06–08, G. firma dari Thailand (KY315293.1), dan G. changii dari Tiongkok (OP669568.1), yang membentuk klaster, tanpa perbedaan urutan basa yang diamati, dan jarak genetiknya adalah 0, yang menunjukkan bahwa G. firma dan G. changii adalah spesies yang sama. Khususnya, klade kedua dan ketiga membentuk kelompok monofiletik dengan dukungan bootstrap yang tinggi (1/100/100) dan jarak genetik di antara keduanya hanya 0,088%. Klade keempat terdiri dari sampel DHI01–02, SJ08, dan G. edulis dari Tiongkok (OP669574.1), yang dikelompokkan bersama, dengan dukungan bootstrap yang tinggi (1/100/100), tidak ada perbedaan urutan basa, dan jarak genetik 0 yang diamati. Klade kelima terdiri dari sampel DL01 dan G. vermiculophylla dari Jepang, yang dikelompokkan bersama dengan dukungan bootstrap tinggi (1/100/100) dan jarak genetik 0,265%. Klade keenam terdiri dari sampel BH01, HW04–05, dan G. tenuistipitata dari Vietnam (MH760425.1), yang dikelompokkan bersama, dengan dukungan bootstrap tinggi (1/100/100), tidak ada perbedaan urutan basa, dan jarak genetik 0 yang diamati. Klade ketujuh terdiri dari sampel HW01, SJ05, NH01–04, WS01–05, dan Gp. heteroclada dari Cina (OP669716.1), yang dikelompokkan bersama, tanpa perbedaan urutan basa dan jarak genetik 0 yang diamati. Klade ketujuh, bersama dengan G. tenuifrons dan Gp. lemaneiformis , membentuk kelompok monofiletik yang berbeda dari spesies Gracilaria lainnya .
4 DISKUSI
Tata nama Gracilariopsis heteroclada J.-F. Zhang & B.-M. Xia agak membingungkan, karena namanya berasal dari cabang-cabangnya yang panjang dan pendek, seperti duri, dan pertama kali diberi nama Gracilaria heteroclada JF Zhang & BM Xia oleh Junfu dan Bangmei ( 1988 ). Selain G. heteroclada , ia memiliki dua sinonim heterotipe: Gracilariopsis bailiniae J. Zhang & BM Xia dan Gracilaria bailiniae (JF Zhang & BM Xia) JF Zhang & BM Xia. Selama beberapa dekade terakhir, kontroversi internasional atas keberadaan genus Gracilariopsis telah menyebabkan banyak perubahan dalam status taksonomi Gp. heteroclada , pergeseran antara Gracilaria dan Gracilariopsis (Abbott et al., 1991 ; Gurgel et al., 2018 ; Junfu & Bangmei, 1991 ; Xia & Zhang, 1999 ). Gracilariopsis dipisahkan dari Gracilaria pada tahun 1949, yang terutama dibedakan oleh dua karakteristik cystocarp (Dawson, 1949 ). Namun, beberapa sarjana percaya bahwa mengandalkan hanya pada karakteristik morfologi yang terbatas untuk klasifikasi tingkat genus tidaklah memadai (Abbott et al., 1991 ; Tseng & Xia, 1999 ; Xia & Zhang, 1999 ; Yamamoto, 1978 ). Dengan diperkenalkannya teknik biologi molekuler, para peneliti seperti Junfu dan Bangmei ( 1991 ), Bird ( 1995 ), Bellorin et al. ( 2002 ), Muangmai et al. ( 2014 ), dan Lyra et al. ( 2021 ) telah menegaskan validitas Gracilariopsis berdasarkan analisis filogenetik morfologi dan molekuler. Menurut pohon filogenetik, banyak spesies dalam Gracilariopsis jelas berbeda dari spesies dalam Gracilaria , yang membentuk klade independen, dan pohon filogenetik yang dibangun dalam penelitian ini menghasilkan hasil yang serupa. Sampel yang dikumpulkan dalam penelitian ini, termasuk HW01, WS01–05, SJ05, dan NH01–04, menunjukkan berbagai warna talus, dari hijau zamrud hingga coklat kemerahan, cabang sekunder yang memanjang, dan cabang tersier yang pendek dan kadang-kadang berduri (Gambar 2a ). Karakteristik morfologi ini sebagian besar selaras dengan deskripsi oleh Xia dan Zhang ( 1999 ) dan Yang et al. ( 2012 ). Lebih jauh lagi, dalam cox1 dan rbcPohon filogenetik L, sampel-sampel ini dikelompokkan bersama dengan Gp. heteroclada yang teridentifikasi , dengan nilai bootstrap tinggi yang diamati, dengan demikian mengonfirmasi identitas taksonomi mereka sebagai Gp. heteroclada . Khususnya, Gp. heteroclada ditemukan di zona pasang surut, kolam air laut, dan teluk bagian dalam yang berarus lambat. Berdasarkan pengamatan lapangan visual, kepadatan populasinya tampak jauh lebih tinggi daripada spesies Gracilariaceae lainnya, karena sering ditemukan dalam jumlah yang lebih besar per satuan luas di beberapa lokasi survei. Individu yang menghuni teluk bagian dalam yang berarus lambat berada dalam kondisi terbaik dan paling melimpah, dengan jenis habitat ini mirip dengan lingkungan yang digunakan untuk budidaya kolam aliran-melalui. Kami percaya bahwa spesies ini memiliki potensi untuk budidaya skala besar di Laut Cina Selatan.
Gracilaria salicornia (C. Agardh) EY Dawson pertama kali dilaporkan oleh Agardh ( 1820 ). Spesies ini tersebar luas di Asia Timur (Phang et al., 2016 ; Xia & Zhang, 1999 ; Yang et al., 2015 ), Asia Tenggara (Beringuela et al., 2020 ; Tseng & Xia, 1999 ; Yang et al., 2015 ), Timur Tengah (Einav et al., 2021 ; Wynne, 2018 ), Afrika (Vieira et al., 2021 ), di antara lokasi lainnya (Sherwood & Guiry, 2023 ; Yadav & Palanisamy, 2020 ). Perbedaan morfologi G. salicornia dari berbagai wilayah secara historis telah menyebabkan kesalahan identifikasi sebagai spesies jamak, yang pada gilirannya mengakibatkan banyak kasus sinonim. Saat ini, G. salicornia memiliki dua sinonim homotipe: Sphaerococcus salicornia C. Agardh dan Corallopsis salicornia (C. Agardh) Greville, dan lima sinonim heterotipe: Corallopsis dichotoma Ruprecht, Corallopsis cacalia J. Agardh, Corallopsis opuntia J. Agardh, Corallopsis concrescens Reinbold, dan Gracilaria cacalia (J. Agardh) EY Dawson. G. salicornia memiliki ruas berbentuk gada, yang berbeda dari ruas spesies Gracilaria lainnya , dan karakteristik morfologi ini memudahkan untuk mengidentifikasinya di alam liar. Sampel yang dikumpulkan dalam penelitian ini, termasuk HW06, SJ07, dan WS13–15, juga menunjukkan karakteristik ini (Gambar 3a ), yang konsisten dengan deskripsi Xia dan Zhang ( 1999 ) dan Yang et al. ( 2012 ). Berdasarkan pohon filogenetik cox1 dan rbc L, HW06, SJ07, dan WS13–15 dikelompokkan bersama dengan G. salicornia yang teridentifikasi , dengan nilai bootstrap tinggi yang diamati, sehingga mengonfirmasi identitas taksonomi mereka sebagai G. salicornia . Temuan Phoprong et al. ( 2007 ) menunjukkan bahwa G. salicornia beradaptasi dengan berbagai suhu dan tingkat salinitas, dengan populasi dari Pulau Phuket mencapai laju fotosintesis maksimum pada suhu 35°C. Selama survei lapangan kami pada bulan Juli dan Agustus, kami mengamati bahwa jumlah G. salicornia jauh lebih tinggi daripada spesies Gracilaria lainnya , dan talusnya berada dalam kondisi yang lebih baik. Sebagai spesies invasif, G. salicorniatelah berkembang biak di Hawaii (Smith et al., 2004 ) dan kemampuan adaptasinya yang kuat terhadap suhu tinggi merupakan salah satu alasan mengapa penyebarannya begitu luas.
Gracilaria vermiculophylla (Ohmi) Papenfus dan Gracilaria tenuistipitata CF Chang & B.-M. Xia memiliki karakteristik morfologi yang sama dengan thalli yang sangat halus (Gambar 4a,g ). Khususnya, mirip dengan G. salicornia , G. vermiculophylla sering dilaporkan sebagai spesies invasif dalam artikel penelitian (Krueger-Hadfield et al., 2021 ; Mendoza-Segura et al., 2023 ; Sfriso et al., 2010 ). Pada tahun 1991, spesies G. vermiculophylla hanya didokumentasikan di Jepang, Vietnam, dan Cina (Xia & Zhang, 1999 ). Namun, spesies ini saat ini ditemukan hingga ke timur Korea (Yang et al., 2015 ), barat di berbagai negara Eropa (Burel et al., 2019 ; Sfriso et al., 2023 ; Thomsen et al., 2007 ) dan Amerika Utara (Pedroche & Sentíes, 2020 ), dan Rusia di utara (Rueness, 2010 ), yang meliputi hampir seluruh Belahan Bumi Utara. Perluasan populasi G. vermiculophylla yang cepat dikaitkan dengan kemampuan adaptasi ekologisnya yang kuat (Roleda et al., 2012 ). G. tenuistipitata umumnya ditemukan di sepanjang pantai Tiongkok selatan dan Asia Tenggara (Belous et al., 2021 ; Xia & Zhang, 1999 ; Yang et al., 2015 ). Karena tantangan yang terkait dengan budidayanya, G. tenuistipitata menjadi spesies Gracilaria utama untuk budidaya kolam di Tiongkok selatan pada tahun 1980-an (Wang et al., 2020 ). Namun, beberapa penelitian telah menunjukkan bahwa G. tenuistipitata memiliki kandungan agar yang jauh lebih rendah daripada Gp. lemaneiformis (Craigie et al., 1984 ), yang menarik minat para ahli fikologi Tiongkok dalam budidaya Gp. lemaneiformis skala besar sejak akhir abad ke-20. Akibatnya, area budidaya G. tenuistipitata belum berkembang lebih jauh.
Gracilaria edulis (SG Gmelin) PC Silva merupakan spesies rumput laut penting secara ekonomi yang tersebar luas di perairan tropis dan subtropis (Bhushan et al., 2023 ). Dalam penelitian ini, sampel DHI01–02 dan SJ08 memiliki thalli seperti sapu dengan pangkal kemerahan pada cabang-cabangnya (Gambar 5a,b ), yang konsisten dengan deskripsi G. edulis oleh Xia dan Zhang ( 1999 ) dan Yang et al. ( 2012 ). Menurut pohon filogenetik, DHI01–02 dan SJ08 dikelompokkan bersama dengan G. edulis , dengan jarak genetik 0 yang diamati. G. edulis merupakan spesies yang tahan panas dan bahan baku penting untuk produksi agar di India (Bhushan et al., 2023 ). Namun, metode utama untuk memperoleh spesies ini secara lokal adalah melalui koleksi liar, dengan budidayanya yang terbatas (Ganesan et al., 2011 ; Sakthivel & Devi, 2015 ). Dalam studi ini, sampel G. edulis yang dikumpulkan dari kolam aliran-melalui selama musim panas mempertahankan kondisi yang baik, menunjukkan bahwa spesies ini memiliki potensi untuk budidaya skala besar di Cina. Sampel SJ01–04 terjadi di zona pasang surut yang sama dengan G. edulis . Pengamatan morfologi menunjukkan bahwa, selain sedikit lebih besar, mereka sangat mirip dengan Gracilaria coronopifolia J. Agardh seperti yang dijelaskan oleh Xia dan Zhang ( 1999 ). Namun, urutan gen cox1 yang diperoleh dari sampel-sampel ini menunjukkan bahwa mereka dikelompokkan bersama dengan Gracilaria fisheri (BM Xia & IA Abbott) IA Abbott, J. Zhang & BM Xia (Gambar 7 ). Selain itu, hasil sequencing gen rbc L menempatkan mereka dalam klade yang sama dengan G. firma , dengan jarak genetik yang hanya 0,088% yang diamati (Gambar 8 ), yang konsisten dengan hasil pohon filogenetik yang dibangun oleh Ng et al. ( 2017 ). Menurut Kim et al. ( 2010 ), Saunders ( 2009 ), dan Le Gall dan Saunders ( 2010 ), jarak genetik intraspesifik yang lebih besar dari 2% harus dianggap sebagai spesies yang berbeda. Dalam penelitian ini, G. fisheri dan G. firma diidentifikasi sebagai spesies yang berbeda berdasarkan pohon filogenetik cox1 , sedangkan pohon filogenetik rbc L menunjukkan bahwa kedua spesies tersebut adalah sama. Meskipun demikian, hubungan taksonomi antara G. fisheri dan G. firma memerlukan penelitian lebih lanjut.G. fisheri hanya tersebar di beberapa negara Asia Tenggara (Beringuela et al., 2020 ; Yang et al., 2015 ) dan belum dilaporkan di Tiongkok. Karena kurangnya bukti morfologi yang memadai (misalnya, spermatangia dan filamen transfer) dan perbedaan tertentu dalam penampilan talus dari deskripsi G. fisheri oleh Abbott et al. ( 1991 ), kami secara tentatif menyebut sampel WS1–10 yang dikumpulkan dalam penelitian ini sebagai Gracilaria sp. 1.
Gracilaria firma CF Chang & B.-M. Xia pertama kali dilaporkan oleh Chang ( 1976 ) di Xindi, Guangdong, Tiongkok, dan Gracilaria changii (BM Xia & IA Abbott) IA Abbott, J. Zhang & BM Xia pertama kali dilaporkan oleh Abbott et al. ( 1991 ) di Malaysia. Kedua spesies ini secara morfologis mirip dan hanya dapat dibedakan berdasarkan jenis spermatangia. Dengan kemajuan penelitian molekuler, Ng et al. ( 2017 ) menganalisis karakteristik morfologi, serta sekuens gen cox1 dan rbc L dari G. firma dan G. changii , dan hasilnya menunjukkan bahwa keduanya adalah spesies yang sama, yang konsisten dengan temuan Li et al. ( 2024 ). Namun, karena kurangnya perbandingan dengan sekuens gen spesimen tipe, pandangan ini masih kontroversial (Wang et al., 2023 ). Sampai saat ini, G. changii tidak dianggap sinonim dari G. firma pada basis data AlgaeBase ( http://www.algaebase.org ). Sampel HW02–03, SJ06, dan WS06–12 yang dikumpulkan dalam studi ini berwarna hijau, panjang 10–25 cm, dengan pangkal cabang yang sangat menyempit, dan tidak ditemukan filamen penyerap di dalam sistokarp (Gambar 6a–f ). Karakteristik ini konsisten dengan deskripsi G. firma oleh Chang ( 1976 ) dan Xia dan Zhang ( 1999 ). Dalam pohon filogenetik rbc L, spesimen yang dikumpulkan dalam studi ini dikelompokkan bersama dengan spesimen “ G. firma ” yang dikumpulkan oleh Ng et al. ( 2017 ) dari Vietnam dan spesimen “ G. changii ” yang dikumpulkan oleh Wang et al. ( 2023 ) dari Tiongkok, dengan jarak genetik 0 yang diamati. Lebih jauh lagi, lokasi pengumpulan sampel HW02–03 dalam studi ini dekat dengan lokasi tipe G. firma , dan jarak garis lurus kurang dari 50 km, yang meningkatkan keandalan identifikasi sampel ini sebagai G. firma . Khususnya, Wang et al. ( 2023 ) mengumpulkan spesimen (ZJ5-1) dari lokasi tipe G. firma , yang dikelompokkan bersama dengan Gracilaria hainanensis CF Chang & BM Xia. Spesimen ini diidentifikasi sebagai G. firma , dan kami percaya ini adalah kasus dua spesies yang hidup bersamaan, dengan kesalahan identifikasi yang diakibatkan oleh kurangnya analisis morfologi terperinci dari spesimen yang dikumpulkan. Sebagai kesimpulan, studi ini mendukung penanganan G. changii dan G. firmasebagai spesies yang sama. Kelompok penelitian kami juga mengumpulkan beberapa sampel (TCI01–03) dengan pangkal cabang yang sangat menyempit (Gambar 6h ) dari hutan bakau di Pulau Techeng. Akan tetapi, talusnya lebih besar dan lebih panjang daripada talus G. firma yang dikumpulkan dari zona pasang surut dengan substrat berpasir-lumpur, dan warnanya merah tua. Kami telah berulang kali mencoba mengekstraksi sekuens DNA dari sampel-sampel ini tetapi tidak berhasil, dan memperoleh data morfologi yang lengkap merupakan tantangan karena terbatasnya jumlah sampel yang dapat kami kumpulkan. Oleh karena itu, kami sementara menyebut sampel TCI01–03 sebagai Gracilaria sp.2.
5 KESIMPULAN
Survei yang dilakukan dari tahun 2021 hingga 2024 mengidentifikasi spesies Gracilariaceae yang toleran terhadap panas dan cocok untuk budidaya skala besar di Laut Cina Selatan. Kami mengidentifikasi enam spesies yang toleran terhadap panas, termasuk satu Gracilariopsis dan lima spesies Gracilaria : Gp. heteroclada , G. salicornia , G. vermiculophylla , G. tenuistipitata , G. edulis , G. firma , dan dua spesies yang memerlukan studi lebih lanjut ( Gracilaria sp.1 dan Gracilaria sp.2). Penelitian ini meningkatkan pemahaman kita tentang keragaman taksonomi spesies Gracilariaceae yang toleran terhadap panas dan meletakkan dasar bagi pengembangan spesies yang cocok untuk budidaya skala besar di wilayah tersebut.


