
ABSTRAK
Menilai variasi dalam perilaku sosial hewan yang hidup berkelompok dapat memberikan wawasan tentang dampak stresor populasi. Dengan menggunakan citra satelit resolusi sangat tinggi (VHR) yang diperoleh selama 3 tahun, kami membandingkan ukuran dan komposisi kelompok sosial, serta kohesi spasial dan sosial dalam tiga populasi beluga ( Delphinapterus leucas ) yang mengalami tingkat paparan panen dan lalu lintas kapal yang berbeda: Cumberland Sound, Eastern High Arctic-Baffin Bay, dan Western Hudson Bay. Kami selanjutnya mengeksplorasi hubungan antara panen, aktivitas kapal, kepadatan beluga, dan konteks sosial dengan karakteristik pengelompokan beluga untuk memprediksi pendorong potensial dinamika kelompok sosial beluga. Ukuran kelompok rata-rata menurun dengan tingkat panen, mungkin mencerminkan efek penurunan populasi atau penghapusan individu sosial utama. Populasi yang terpapar peningkatan aktivitas kapal baru-baru ini dikaitkan dengan ukuran kelompok yang lebih besar dan kohesi spasial yang lebih besar, mungkin menunjukkan peningkatan kewaspadaan dalam menanggapi lalu lintas kapal. Kelompok campuran dewasa-remaja lebih besar dan memiliki jarak antarkelompok yang lebih kecil daripada kelompok yang hanya terdiri dari paus dewasa, yang mungkin mencerminkan ketergantungan paus yang lebih muda pada perawatan alloparental. Studi ini menyediakan aplikasi baru citra satelit VHR untuk pemantauan beluga dan menyoroti dampak potensial stresor antropogenik pada populasi beluga.
1 Pendahuluan
Penurunan luas dan durasi es laut Arktik Kanada menunjukkan bahwa aktivitas antropogenik akan meningkat di Arktik (Copland et al. 2021 ; Dawson et al. 2018 ; Nielsen 2009 ; Pizzolato et al. 2016 ; Stephenson et al. 2013 ) dengan dampak pada beberapa cetacea Arktik melalui peningkatan paparan kebisingan, risiko tumpahan beracun, dan tabrakan kapal (Halliday et al. 2017 ; Huntington et al. 2015 ; Reeves et al. 2014 ). Ada juga bukti bahwa pecahnya es laut lebih awal memungkinkan akses lebih awal ke tempat perburuan tradisional dan dengan demikian waktu yang lebih lama untuk memanen cetacea Arktik (Nielsen 2009 ). Meskipun paparan jangka pendek terhadap kapal dapat mengakibatkan perubahan perilaku (Martin et al. 2023 ), efek sub-mematikan kumulatif dari paparan kronis terhadap aktivitas panen dan kebisingan terkait kapal lebih sulit dideteksi dan saat ini belum dipahami dengan baik (Mesnick et al. 2023 ).
Aktivitas antropogenik dihipotesiskan mengubah dinamika kelompok sosial beberapa spesies yang hidup berkelompok di darat (Ausband et al. 2017 ; Bond et al. 2019 ; Gall et al. 2022 ; Manor and Saltz 2003 ; Meier et al. 2023 ), dan spesies cetacea sosial seperti paus pembunuh ( Orcinus orca ) (Busson et al. 2019 ), paus sperma ( Physeter macrocephalus ) (Whitehead et al. 2012 ), paus bungkuk ( Sousa plumbea ), dan lumba-lumba hidung botol biasa ( Tursiops truncatus ) (Koper et al. 2016 ; La Manna et al. 2023 ). Bahasa Indonesia: Menilai dinamika kelompok pada cetacea Arktik sosial, seperti beluga ( Delphinapterus leucas ), oleh karena itu dapat memberikan wawasan tentang dampak sub-mematikan dari paparan stresor antropogenik di Arktik. Pada prinsipnya, ukuran kelompok harus mencerminkan efek gabungan dari manfaat dan biaya pengelompokan, meskipun kelompok sosial sering kali melebihi ukuran optimalnya (Silk 2007 ). Beluga dianggap mengikuti sistem sosial fisi-fusi dan biasanya membentuk kelompok yang terdiri dari kurang dari 10 individu dalam 3 hingga 4 panjang tubuh (Boltunov dan Belikov 2002 ; O’Corry-Crowe et al. 2020 ) di mana individu dapat memperoleh manfaat dari perawatan alloparent, pembelajaran sosial dan budaya, atau kewaspadaan yang lebih tinggi terhadap ancaman (Aubin et al. 2021 , 2022 ; Krasnova et al. 2014 ). Metrik dinamika kelompok sosial mencakup ukuran kelompok dan ukuran kohesi kelompok, seperti jarak antar individu (IID) atau karakteristik berenang (misalnya, arah dan gerakan tersinkronisasi), yang mungkin menunjukkan investasi dalam perlindungan atau komunikasi, dan akhirnya kewaspadaan terhadap pemicu stres eksternal (Senigaglia et al. 2012 ; Warburton dan Lazarus 1991 ). Karakteristik tingkat kawanan seperti jarak antar kelompok (IGD) dapat menunjukkan investasi serupa dalam manfaat kehidupan kelompok di tingkat kawanan. Ukuran, IID, dan varians dalam arah berenang individu telah dijelaskan untuk kelompok beluga Laut Beaufort Timur (EBS) (Mayette et al. 2022 ). Membandingkan karakteristik kelompok di seluruh populasi yang berbeda dapat membantu mengidentifikasi pendorong antropogenik spesifik populasi dari karakteristik sosial ini pada beluga. Mengingat sistem sosial dapat memengaruhi reproduksi, populasi dan kesehatan individu, serta keberhasilan mencari makan pada hewan yang hidup berkelompok (La Manna et al. 2023 ), memahami dampak potensial dari stresor antropogenik pada dinamika sosial beluga dapat memberikan informasi yang relevan untuk pengelolaan beluga.
Selain stresor antropogenik, karakteristik kelompok beluga dapat mencerminkan kepadatan (yaitu, jumlah paus atau kelompok yang hadir di area tertentu) dan konteks sosial (yaitu, jumlah dan usia anggota kelompok). Misalnya, kepadatan paus yang lebih tinggi dalam bersuara dapat mengganggu komunikasi antara anggota kelompok, yang mungkin mengubah struktur kelompok (La Manna et al. 2023 ), sementara kepadatan paus yang lebih tinggi dapat membatasi pengaturan spasial kelompok sosial. Dari perspektif sosial, kelompok beluga yang lebih besar mungkin memerlukan lebih banyak ruang daripada kelompok yang lebih kecil atau mungkin kurang bergantung pada kewaspadaan tingkat kawanan terhadap ancaman karena kewaspadaan tingkat kelompok yang secara inheren lebih besar (Krause dan Ruxton 2002 ). Selain itu, odontocetes dewasa cenderung meningkatkan perilaku pengelompokan, misalnya, ukuran (La Manna et al. 2023 ) dan kohesi (Mayette et al. 2022 ), ketika paus muda hadir (Oremus et al. 2012 ). Penelitian mengenai efek gabungan konteks sosial dan kepadatan beserta faktor antropogenik belum dieksplorasi pada beluga sejauh pengetahuan kami.
Citra satelit resolusi sangat tinggi (VHR) muncul sebagai alat yang efektif untuk mengamati dan memantau cetacea (Belanger et al. 2024 ; Charry et al. 2021 ; Cubaynes et al. 2019 ; Fretwell et al. 2014 , 2023 ; Hodul et al. 2023 ; Höschle et al. 2021 ; Sherbo et al. 2024 ; Stewart et al. 2024 ; Watt et al. 2023 ). Ada beberapa keuntungan dalam memanfaatkan citra satelit VHR untuk menggambarkan karakteristik kelompok sosial beluga. Misalnya, citra VHR menangkap pandangan seketika dari area yang membentang ribuan km 2 (Fretwell et al. 2014 ; Cubaynes et al. 2019 ; Bamford et al., 2020), yang memungkinkan pengamatan simultan kelompok beluga dan distribusi spasialnya, yang sulit dinilai dengan citra udara yang menggunakan metode survei transek. Citra VHR juga non-invasif, yang memastikan pengamatan beluga tidak diubah oleh platform survei. Namun, citra VHR bisa mahal untuk diperoleh, waktu akuisisi citra tidak dapat diprediksi (Khan et al. 2023 ), dan citra ditangkap pada resolusi yang lebih rendah daripada sebagian besar platform survei udara. Meskipun ada keterbatasan ini, citra VHR telah terbukti menghasilkan estimasi kelimpahan permukaan yang tidak dikoreksi serupa dengan yang dihasilkan oleh drone (Stewart et al. 2024 ), dan oleh karena itu mungkin merupakan metode yang efektif untuk menggambarkan karakteristik kelompok beluga, meskipun sebagian besar masih belum teruji (lihat Rivas et al. 2024 ).
Di sini, kami menggunakan citra satelit VHR untuk mengeksplorasi dinamika kelompok sosial untuk beluga di Cumberland Sound (CS), Eastern High Arctic-Baffin Bay (EHA-BB), dan Western Hudson Bay (WHB). Populasi ini terpapar pada berbagai tingkat panen komersial masa lalu dan panen subsisten saat ini (Stewart 2018 ; Watt 2021 ) dan memiliki distribusi di dalam (EHA-BB dan WHB) dan di luar (CS) rute pengiriman utama. Secara khusus, kami membandingkan karakteristik kelompok beluga (ukuran kelompok, komposisi kelompok, jarak antar kelompok, jarak antar individu, dan varians dalam arah renang kelompok) di ketiga populasi ini sambil mengeksplorasi pengaruh stresor antropogenik (panen dan lalu lintas kapal), kepadatan, dan konteks sosial sebagai pendorong potensial dari karakteristik ini (Tabel 1 ). Hasil dari studi ini menyoroti potensi citra satelit VHR untuk memantau struktur sosial spesies cetacea dan untuk berkontribusi pada pemahaman dinamika kelompok sosial beluga dalam menanggapi stresor antropogenik di Arktik.
| Kategori variabel | Variabel | Karakteristik kelompok | Perubahan yang diprediksi dalam karakteristik kelompok | Mekanisme yang diprediksi |
|---|---|---|---|---|
| Stresor antropogenik | Panen jangka panjang atau aktivitas kapal | Ukuran kelompok | Meningkatkan | Beluga harus meningkatkan kewaspadaan atau menunjukkan peningkatan stres terhadap stresor kronis yang mengarah pada peningkatan perilaku pengelompokan. |
| Komposisi kelompok | Meningkatkan | |||
| Jarak antar kelompok | Mengurangi | |||
| Jarak antar individu | Mengurangi | |||
| Varians dalam orientasi renang | Mengurangi | |||
| Perubahan jangka pendek dalam panen atau aktivitas kapal | Ukuran kelompok | Meningkatkan | Beluga harus menunjukkan kewaspadaan yang lebih tinggi atau menunjukkan stres yang lebih tinggi terhadap peningkatan paparan stresor antropogenik yang mengarah pada peningkatan perilaku pengelompokan. | |
| Komposisi kelompok | Meningkatkan | |||
| Jarak antar kelompok | Mengurangi | |||
| Jarak antar individu | Mengurangi | |||
| Varians dalam orientasi renang | Mengurangi | |||
| Kepadatan | Jumlah kelompok per 1 km 2 | Jarak antar kelompok | Mengurangi | Kepadatan kelompok yang lebih tinggi akan menyebabkan ruang antar kelompok menjadi lebih kecil |
| Jumlah paus dalam radius 1 km | Ukuran kelompok | Mengurangi | Beluga harus membentuk kelompok yang lebih kecil dan lebih kohesif untuk meningkatkan komunikasi di lingkungan yang lebih bising, dan meningkatkan jumlah orang dewasa dalam kelompok ketika anak-anaknya berada di daerah dengan kepadatan lebih tinggi. | |
| Komposisi kelompok | Meningkatkan | |||
| Jarak antar individu | Mengurangi | |||
| Varians orientasi renang | Mengurangi | |||
| Konteks sosial | Tipe grup | Ukuran kelompok | Meningkatkan | Ukuran dan kekompakan kelompok akan meningkat ketika paus muda hadir |
| Jarak antar kelompok | Mengurangi | |||
| Jarak antar individu | Mengurangi | |||
| Varians dalam orientasi renang | Mengurangi | |||
| Ukuran kelompok | Jarak antar kelompok | Meningkatkan | Kelompok yang lebih besar memerlukan lebih banyak ruang atau kurang bergantung pada kohesi pada tingkat kawanan untuk kewaspadaan terhadap stresor antropogenik. |
2 Metode
2.1 Populasi Studi
Beluga dari populasi CS adalah penghuni Cumberland Sound sepanjang tahun, bermigrasi antara tempat musim dingin di muara Cumberland Sound dan tempat musim panas utama di Clearwater Fiord di ujung utara suara (Booy et al. 2021 ; COSEWIC 2020 ). Beluga CS saat ini ditetapkan sebagai spesies yang terancam punah oleh Committee on the Status of Endangered Wildlife in Canada (COSEWIC) dengan estimasi populasi saat ini sebesar 1.090 paus (95% CI = 617–1.864) (Watt et al. 2020 ). Beluga CS diasumsikan secara eksklusif diburu oleh pemburu dari Pangnirtung, Nunavut dengan kuota tangkapan 41 paus per tahun sejak 2002 (Stewart 2018 ). Baik tempat musim panas maupun musim dingin CS tidak terletak langsung di jalur rute pelayaran utama mana pun (Gambar 1 ; Tabel S1 ).

Beluga dari EHA-BB tersebar di sebagian besar Kepulauan Arktik Kanada selama musim panas dan di sepanjang pantai barat Greenland selama musim dingin (Heide-Jorgenssen et al. 1993 ; Richard et al. 1998 ). Beluga EHA-BB saat ini ditetapkan oleh COSEWIC sebagai perhatian khusus, dengan estimasi populasi lengkap terakhir sebesar 21.213 paus (95% CI = 10.985–32.619) pada tahun 1996 (Innes et al. 2002 ). Populasi ini saat ini dipanen tanpa kuota dari komunitas Kanada dan Greenland (Watt 2021 ) selama musim panas dan musim dingin, masing-masing (Heide-Jørgensen dan Rosing-Asvid 2002 ; Watt 2021 ) ( https://nammco.no/beluga ). Sebagian besar distribusi beluga EHA-BB berada di jalur pelayaran Northwest Passage (Gambar 1 ; Tabel S1 ).
Beluga dari populasi WHB bermigrasi dari Selat Hudson di musim dingin ke perairan pesisir di Teluk Hudson bagian barat di musim panas, dengan konsentrasi besar terbentuk di muara Sungai Churchill, Seal, dan Nelson. Beluga WHB saat ini terdaftar sebagai tidak berisiko oleh COSEWIC dengan estimasi populasi saat ini sebesar 54.473 paus (95% CI = 44.988–65.957) (Matthews et al. 2017 ). Beluga WHB dipanen dari komunitas Nunavut dan Nunavik di sepanjang jalur migrasi mereka, dan sebagian besar distribusinya berada di jalur rute pelayaran Jembatan Arktik (Gambar 1 ; Tabel S1 ).
2.2 Pengumpulan dan Analisis Citra
Kami menugaskan satelit VHR WorldView 3 (WV3; Maxar Technologies Inc., Colorado, AS) untuk memperoleh citra antara tahun 2020 dan 2022 yang merangkum sebagian dari area musim panas utama beluga CS, EHA-BB, dan WHB (Gambar 2 ). Semua citra diperoleh dengan resolusi spasial 0,31 m, meskipun semua citra berada di luar nadir yang mengurangi resolusi menjadi sekitar 0,32–0,39 m rata-rata di semua citra kami (Tabel S2 ). Citra dikumpulkan dalam kondisi cuaca yang sama dan memiliki status Laut Beaufort 3 atau kurang. Karena beberapa lokasi hanya memiliki citra pankromatik yang tersedia, kami melakukan analisis menggunakan pita pankromatik hanya untuk setiap citra. Ada tutupan awan minimal di area musim panas CS dan EHA-BB (0%–15%), dan sekitar 45% tutupan awan di area yang dapat disurvei (yaitu, area di atas air tempat paus diperkirakan ditemukan) di Sungai Churchill (Gambar S1 ).

Beluga tumpang tindih dengan beberapa spesies mamalia laut dalam distribusi CS, EHA-BB, dan WHB yang dapat ditangkap dalam citra kami, termasuk narwhal ( Monodon monoceros ), paus kepala busur ( Balaena mysticetus ), paus pembunuh, dan harpa ( Pagophilus groenlandicus ), berjanggut ( Erignathus barbatus ), bercincin ( Pusa hispida ), dan anjing laut pelabuhan ( Phoca vitulina ). Paus kepala busur, paus pembunuh, dan anjing laut harpa tidak mungkin disalahartikan sebagai beluga dewasa karena ukuran, warna, dan perilaku pengelompokannya yang berbeda (Jefferson et al. 2015 ; Sherbo et al. 2024 ); narwhal dewasa lebih sulit dibedakan dari beluga dewasa dalam citra VHR, tetapi mereka memiliki karakteristik yang berbeda (lihat Charry et al. 2021 ). Narwhal tidak berkumpul di Clearwater Fiord atau Sungai Churchill, tetapi narwhal dapat hadir di Teluk Creswell pada saat akuisisi gambar kami (Laidre et al. 2003 ). Namun, beluga diketahui berkumpul di Teluk Creswell dalam jumlah yang sangat tinggi (> 750 beluga dalam satu hari (Innes et al. 2002 )), sedangkan perkemahan lapangan di Teluk Creswell pada tahun 2022–2024 hanya sesekali melihat kelompok kecil narwhal (obs. pribadi J. Stewart). Warna putih beluga dewasa juga membuatnya lebih mudah dideteksi dalam citra satelit pankromatik dibandingkan dengan narwhal dewasa, meskipun beluga abu-abu gelap mirip dengan narwhal dan beberapa spesies anjing laut dalam hal warna. Anjing laut pelabuhan hadir di Sungai Churchill tetapi paling melimpah di lokasi-lokasi pelarian lebih jauh ke hulu dari tempat gambar kami diambil (Florko et al. 2018 ), dan anjing laut bercincin dan berjanggut tidak diamati dalam kepadatan besar di Teluk Creswell di perkemahan lapangan sebelumnya (obs. pribadi J. Stewart). Berdasarkan pengamatan sebelumnya, kami perkirakan sedikit narwhal dan anjing laut di wilayah survei EHA-BB dan WHB dibandingkan dengan beluga (Florko et al. 2018 ; Innes et al. 2002 ; Watt et al. 2023 ). Jadi, meskipun tidak mungkin untuk dikesampingkan, berdasarkan pengamatan sebelumnya, kami berasumsi hanya sebagian kecil dari beluga muda atau abu-abu yang teridentifikasi bisa jadi adalah narwhal atau anjing laut.
Citra satelit dianalisis dalam ArcGIS Pro 10.8.2 (ESRI Inc., Redlands, CA, AS). Seorang pengamat mengidentifikasi target beluga secara visual sebagai titik georeferensi, secara sistematis menggunakan kisi dengan sel individual berukuran 250 m × 250 m yang dihamparkan pada setiap citra satelit. Target beluga diklasifikasikan sebagai tidak pasti atau pasti berdasarkan fitur tubuh yang ada (misalnya, bulu ekor yang terlihat, kepala, atau sirip dada) atau konteks spasial (misalnya, kedekatan dengan pantai, atau target paus tertentu lainnya). Kami hanya menggunakan target dengan klasifikasi tertentu dalam analisis. Vektor panjang tubuh, yang dapat diukur langsung dari citra orthorektifikasi, ditambahkan dari perkiraan awal dan akhir setiap target beluga, dengan titik ditambahkan di awal, tengah, dan akhir vektor menggunakan alat ‘Buat Titik Sepanjang Garis’ dan digunakan untuk menganalisis parameter kelompok.
2.3 Karakteristik Kelompok
Kami menggunakan aturan rantai 12 m (3–4 panjang tubuh) sebagai jarak batas kelompok kami berdasarkan kurva fungsi kerapatan jarak beluga ke tetangga terdekatnya (Syme et al. 2022 ) (Gambar S2 ). Ukuran kelompok condong positif bahkan ketika ditransformasikan log dan oleh karena itu diubah menjadi variabel kategoris dengan tiga tingkat yang relevan secara biologis: paus tunggal, pasangan paus, dan kelompok paus (3+), ketika digunakan sebagai variabel penjelas. Kami memperkirakan usia target beluga dari ukuran relatifnya. Target yang panjangnya tiga meter atau lebih diklasifikasikan sebagai dewasa atau subdewasa, dengan ini disebut sebagai dewasa; target yang panjangnya kurang dari tiga meter diklasifikasikan sebagai remaja atau anak sapi, dengan ini disebut sebagai remaja (Luque dan Ferguson 2006 ; Mayette et al. 2022 ). Jika panjang target tidak dapat ditentukan dengan yakin dari vektor panjang tubuh, maka target tersebut diklasifikasikan sebagai tidak diketahui. Kelompok dengan satu atau lebih paus beluga yang tidak diketahui usianya diklasifikasikan sebagai “Tidak Diketahui”. Kami memperoleh pengukuran jarak antarkelompok (IGD) geodesik minimum dari lokasi rata-rata koordinat titik tengah setiap paus dalam satu kelompok ke kelompok paus terdekat atau paus soliter menggunakan paket R “geosphere” (Hijmans 2022 ).
Kami memperoleh mean IID dan orientasi tubuh semua individu dalam kelompok yang menampilkan dua atau lebih paus sebagai proksi untuk kohesi sosial. Kami menggunakan paket “geosphere” untuk menghitung mean IID berdasarkan jarak geodesik antara koordinat titik tengah setiap paus individu dalam suatu kelompok. Kami menentukan orientasi setiap beluga dalam suatu kelompok menggunakan dua titik akhir dari setiap vektor panjang tubuh (Utara = 0°, Barat = −90°, Selatan = −180°/180°, Timur = 90°) yang darinya kami menghitung mean resultan varians panjang untuk setiap kelompok menggunakan paket R “circular” (Agostinelli dan Lund 2023 ; Cremers dan Klugkist 2018 ). Varians panjang rata-rata yang dihasilkan adalah ukuran penyebaran orientasi yang direpresentasikan oleh nilai antara nol dan satu, di mana satu menunjukkan penyebaran orientasi yang besar dan nol menunjukkan semua paus dalam suatu kelompok memiliki orientasi yang sama (Cremers dan Klugkist 2018 ). Kami menyebut varians panjang rata-rata yang dihasilkan sebagai varians orientasi dari sini ke depan.
2.4 Variabel Gangguan Antropogenik
Kami mengkaji pengaruh lalu lintas kapal pada karakteristik kelompok sosial beluga menggunakan data lalu lintas dari Sistem Informasi Manajemen Lalu Lintas Kapal Kanada (VTMIS; https://www.innav.gc.ca ) yang dapat diakses secara bebas. Di Zona Lalu Lintas Arktik (NORDREG), VTMIS terutama mencakup informasi tentang kapal apa pun dengan tonase kotor 300 atau lebih atau panjang 20 m atau lebih (lihat Lampiran S1 untuk detail lebih lanjut) (Menteri Kehakiman 2010 ). Oleh karena itu, data ini tidak mencakup kapal pesiar kecil yang mungkin sering mengunjungi area studi kami dan dapat mengubah perilaku beluga. Beluga di Sungai Churchill, tempat kapal pesiar kecil paling banyak ditemukan di lokasi studi kami (Malcolm dan Penner 2011 ; Westdal et al. 2023 ), tidak menunjukkan perubahan perilaku terhadap kapal pesiar kecil (Ausen et al. 2022 ). Namun, apakah struktur kelompok beluga diubah oleh keberadaan kapal pesiar kecil belum diperiksa.
Kami menggunakan jumlah waktu kapal melaporkan ke VTMIS di yurisdiksi pelabuhan yang terletak di jalur distribusi atau migrasi populasi beluga yang sesuai sebagai proksi untuk aktivitas kapal (Gambar 1 ; Lampiran S1 ). Untuk menyederhanakan, kami mengekstrak aktivitas kapal dari semua pelabuhan untuk semua bulan kecuali Juli hingga Oktober untuk pelabuhan Greenland, dan Juli hingga Agustus untuk pelabuhan Hudson Bay timur, karena beluga EHA-BB dan WHB, masing-masing, dianggap tidak ada di area tersebut selama periode waktu ini (de March et al. 1992 ; de March dan Postma 2003 ; Richard et al. 1998 , 2001 ). Karena data yang tidak konsisten pada tahun 2006 dan sebelumnya, kami hanya memasukkan tahun dari 2007 dan seterusnya dalam analisis. Entri duplikat atau entri dengan data yang tidak mencukupi (misalnya, pengidentifikasi kapal atau lokasi yang hilang) dihapus dari kumpulan data. Data aktivitas kapal terakhir yang digunakan dalam analisis kami terdiri dari jumlah rata-rata transit kapal per tahun selama 13 tahun hingga tahun sebelum pengambilan citra satelit terkait. Selain itu, dalam upaya untuk menangkap pengaruh tren terkini dalam gangguan kapal terhadap karakteristik kelompok sosial beluga, kami membuat variabel yang mewakili penyimpangan jangka pendek dari tingkat lalu lintas rata-rata. Ini dihitung sebagai perbedaan antara rata-rata transit kapal per tahun selama lima tahun terakhir sebelum citra satelit terkait (tahun 1–5) dan rata-rata tahunan 8 tahun sebelumnya (tahun 6–13), data tepercaya tertua yang tersedia:
![]()
Kami menggunakan data panen beluga yang dilaporkan yang diproduksi oleh Fisheries and Oceans Canada Resource Management untuk menyelidiki pengaruh potensial dari stresor terkait panen, seperti kebisingan, yang dikejar, dan hilangnya anggota kelompok, pada dinamika kelompok sosial beluga (Lampiran S1 ). Data ini tidak termasuk paus yang tertabrak dan hilang karena tidak ada cukup data untuk ketiga populasi. Untuk memperhitungkan efek yang tidak proporsional dari panen tahunan yang mungkin terjadi pada suatu populasi (misalnya, panen 40 paus adalah ~4% dari populasi CS, dibandingkan dengan 0,07% dari populasi WHB), total panen tahunan diwakili dengan membagi panen tahunan rata-rata dengan kelimpahan populasi yang diperkirakan saat ini. Kami menggunakan tingkat panen per kapita tahunan rata-rata selama 15 tahun hingga tahun sebelum penangkapan citra satelit yang sesuai untuk secara kasar mencocokkan kerangka waktu yang sama dari data aktivitas kapal. Penyimpangan jangka pendek dari tingkat panen per kapita rata-rata direpresentasikan sebagai perbedaan antara rata-rata panen per kapita tahunan selama lima tahun terakhir sebelum citra satelit terkait (tahun 1–5) dan rata-rata tahunan 10 tahun sebelumnya (tahun 6–15):
![]()
Namun, perubahan panen jangka pendek tidak berbeda secara signifikan antar populasi (Gambar S3 ) dan tidak digunakan dalam analisis kami.
2.5 Variabel Kepadatan
Kami menilai pengaruh kepadatan beluga pada tingkat kelompok dan individu terhadap karakteristik kelompok. Kami memperkirakan kepadatan kelompok sebagai jumlah kelompok dibagi dengan luas perairan yang dapat diamati dalam setiap citra satelit:
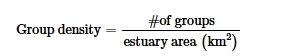
Kepadatan kelompok hanya digunakan dalam analisis IGD. Kami menghitung proksi untuk kepadatan individu dan dengan demikian, potensi efek kebisingan pada karakteristik kelompok (Tabel 1 ), sebagai jumlah beluga dalam radius 1 km dari setiap kelompok. Kami memilih radius 1 km karena ini adalah jarak di mana sebagian besar pita oktaf kemungkinan besar akan tetap terdengar oleh beluga lainnya (Vergara et al. 2021 ), dan ini memberikan keseimbangan yang wajar antara menangkap representasi bentang suara lokal untuk setiap kelompok, yang kemungkinan besar paling berpengaruh pada kohesi sosial beluga, dan ukuran setiap area musim panas. Kepadatan individu digunakan dalam analisis ukuran kelompok, IID, dan varians orientasi saja.
2.6 Analisis Statistik
2.6.1 Perbandingan Antar Populasi
Semua analisis statistik dilakukan dalam R versi 2023.06.0 (Tim Inti R 2023 ). Data untuk semua karakteristik kelompok dan parameter antropogenik (Tabel 1 ) tidak memenuhi asumsi normalitas (Shapiro–Wilk: p < 0,05). Oleh karena itu, kami menggunakan uji Kruskal–Wallis diikuti oleh uji post hoc Conover–Iman (Conover dan Iman 1979 ; Dinno 2017 ) untuk menentukan apakah distribusi salah satu parameter berbeda di antara ketiga populasi. Kami juga membandingkan lima karakteristik pengelompokan di masing-masing dari tiga populasi terhadap pengamatan yang mencakup deteksi target yang tidak pasti untuk memperkirakan efek penghapusan deteksi yang tidak pasti dalam analisis kami (Lampiran S1 ).
2.6.2 Membangun Set Model
Kami melengkapi rangkaian kandidat model efek campuran linier umum (GLMM) untuk menyelidiki hubungan antara karakteristik kelompok di ketiga populasi yang digabungkan dan variabel prediktor kami (Tabel 1 ). Kami memisahkan observasi ke dalam tiga set data: (1) semua observasi kelompok tanpa memperhatikan komposisi kelompok, (2) hanya kelompok di mana kelas usia semua paus dalam kelompok tersebut diperkirakan, dan (3) hanya kelompok dari set data dua yang memiliki setidaknya satu juvenil yang hadir. Informasi set model terperinci dapat ditemukan di Lampiran S1 . Singkatnya, set data pertama digunakan untuk melengkapi model untuk memprediksi ukuran kelompok, IGD, IID, dan varians orientasi yang membahas hipotesis spesifik kami, di mana setiap model hanya berisi satu variabel yang berlaku dari setiap kategori variabel (Tabel 1 ). Set data kedua digunakan dengan cara yang sama tetapi menyertakan variabel kategoris yang menggambarkan tipe kelompok sebagai istilah aditif atau interaktif (hanya dewasa atau campuran). Karena kami tertarik pada potensi beluga dewasa untuk membentuk kelompok yang lebih besar di hadapan anak-anaknya, ukuran kelompok dengan kumpulan data ini dihitung sebagai jumlah total individu dikurangi jumlah anak-anak yang ada. Kumpulan data ketiga hanya digunakan untuk menyaingi model untuk memprediksi proporsi orang dewasa yang hadir dalam kelompok dengan setidaknya satu anak-anaknya menggunakan kombinasi variabel yang sama seperti kumpulan model pertama (yaitu, tanpa jenis kelompok sebagai istilah aditif atau interaktif). Dalam semua model, kami menambahkan variabel acak yang mewakili citra satelit tempat pengamatan dilakukan ( n = 5). Kami menguji autokorelasi spasial yang dikelompokkan berdasarkan efek acak gambar di semua model kandidat menggunakan paket DHARMa (Hartig 2022 ). Jika autokorelasi spasial terdeteksi dalam model kandidat mana pun, kami menyesuaikan kembali semua model dengan efek acak pengelompokan kelompok yang bersarang dalam setiap gambar untuk memperhitungkan pola spasial yang diamati dalam setiap gambar. Pengelompokan ditentukan oleh pengelompokan K-means menggunakan jumlah pusat terkecil yang secara wajar sesuai dengan pola pengelompokan di setiap gambar, sementara juga cukup besar untuk memperhitungkan autokorelasi spasial. Model nol (hanya intersep dan efek acak) disertakan dalam semua set model, dan semua model kandidat memiliki faktor inflasi varians < 2 (Zuur et al. 2010 ). Spesifikasi dan set model lengkap dapat ditemukan di Lampiran S1 (Tabel S3 ). Kami memodelkan model menggunakan kriteria informasi Akaike (AIC) untuk set model dengan ukuran sampel > 200, dan AIC yang dikoreksi untuk ukuran sampel kecil (AICc) untuk set model dengan ukuran sampel < 200. Kami melakukan perata-rataan model pada model apa pun dengan ΔAIC ≤ 2 menggunakan “model.avg” dalam paket MuMIn (Barton 2023 ) dan menginterpretasikan koefisien model rata-rata bersyarat . Jika model nol adalah model teratas atau berada dalam dua ΔAIC dari model teratas, kami menginterpretasikan variabel yang kami selidiki sebagai tidak dapat menggambarkan variabel respons yang diberikan secara memadai. Eksplorasi data dilakukan sebelum pemasangan model mengikuti Zuur et al . Kami memeriksa distribusi residual model teratas dan homogenitas varians model menggunakan paket DHARMa (Hartig 2022 ) (Gambar S4 ).
3 Hasil
Sebanyak 2483 target beluga terdeteksi di antara tiga populasi (Tabel 2 ). Dari 2051 deteksi tertentu, 245 dilakukan di Clearwater Fiord, 245 selama 2 hari pada tahun 2020, dengan 96 pada tahun 2021. 1333 paus lainnya terdeteksi di Teluk Creswell, dengan 377 paus di Sungai Churchill. Mengecualikan deteksi yang tidak pasti tidak memiliki dampak signifikan secara statistik pada ukuran kelompok rata-rata, IGD, dan IID untuk semua populasi. Namun, varians orientasi secara signifikan lebih besar pada beluga CS ketika target yang tidak pasti disertakan (Tabel S4 ). Namun, besarnya perbedaan ini kecil dan kemungkinan merupakan hasil dari ukuran sampel yang besar, yang menunjukkan relevansi fungsional yang terbatas. Autokorelasi spasial terdeteksi di semua set model kecuali model yang memprediksi IID ketika kelompok dengan komposisi yang diketahui dipertimbangkan (Tabel S5 ).
| Populasi | Bahasa Inggris | EHA-BB | WHB |
|---|---|---|---|
| Jumlah total paus | 341 | tahun 1333 | 377 |
| Jumlah paus yang menyendiri (% kelompok) | 147 (65.9) | 367 (52.4) | 129 (62.0) |
| # pasangan paus (% kelompok) | 49 (22.0) | 184 (26.2) | 47 (22.6) |
| # grup yang beranggotakan 3+ (% grup) | 27 (12.1) | 150 (21.4) | 32 (15.4) |
| # paus dewasa/sub-dewasa (% paus) | 196 (57.5) | 856 (64.2) | 166 (44.0) |
| # paus muda (% paus) | 14 (4.1) | 53 (4.0) | 36 (9.5) |
| # paus yang tidak diketahui (% paus) | 131 (38.4) | 424 (31.8) | 175 (46.4) |
3.1 Ukuran Kelompok
Distribusi ukuran kelompok berbeda di antara populasi ( χ2 = 17,36, df = 2, p < 0,001), meskipun ukuran kelompok rata-rata < 2 untuk ketiga populasi. Kelompok dalam populasi EHA-BB lebih besar rata-rata daripada populasi CS ( p < 0,001), sementara kelompok dalam populasi WHB (1,81 ± 0,10) serupa dalam ukuran rata-rata dengan populasi EHA-BB ( p = 0,06) dan CS ( p = 0,66). Ukuran kelompok dengan setidaknya satu juvenil yang dikonfirmasi adalah sekitar 3 paus rata-rata di setiap populasi dan kurang dari 1,5 paus untuk kelompok dewasa saja (Tabel 3 ).
| Ukuran | IGD | IDAI | Varians orientasi | Proporsi orang dewasa yang hadir | |||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Bahasa Inggris | EHA-BB | WHB | Bahasa Inggris | EHA-BB | WHB | Bahasa Inggris | EHA-BB | WHB | Bahasa Inggris | EHA-BB | WHB | Bahasa Inggris | EHA-BB | WHB | |
| Semua kelompok | |||||||||||||||
| Jangkauan | 1–11 | 1–16 | 1–11 | 17–1530 | 14–8434 | 14–616 | 0,9–18,6 | 0,4–17,9 | 0,6–16,4 | 0–0,28 | 0–0,51 | 0–0,39 | 0,50–1,00 | 0,33–1,00 | 0,25–1,00 |
| # kelompok | 223 | 701 | 208 | 223 | 701 | 208 | 76 | 334 | 79 | 76 | 334 | 79 | 29 | 133 | 24 |
| Rata-rata | 1.00 | 1.00 | 1.00 | 101 | 43 | 83 | 4.9 | 4.8 | 3.4 | 0,017 tahun | 0,013 | 0,007 tahun | 1.00 | 1.00 | 1.00 |
| Rata-rata ± SE | 1,53 ± 0,07 | 1,90b ± 0,05 | 1,81 inci ± 0,11 | 202 tahun ± 18 | 149b ± 26 | 113 tahun ± 8 | 5,7 inci ± 0,4 | 5,9 ± 0,2 | 4,9b ± 0,5 | 0,031 pon ± 0,006 | 0,058 ± 0,005 | 0,026b ± 0,007 | 0,918 ± 0,032 * | 0,902 ± 0,017 * | 0,839 ± 0,048 * |
| Kelompok dengan komposisi yang diketahui | |||||||||||||||
| # kelompok usia campuran | 6 | 29 | 6 | 6 | 29 | 6 | 6 | 29 | 6 | 6 | 29 | 6 | 6 | 29 | 6 |
| Rata-rata ± SE | 2,83 ± 0,48 | 3,07 ± 0,26 | 3,00 ± 0,37 | 84 ± 24 | 126 ± 30 | 95,7 ± 20 | 3,3 ± 0,7 | 4,7 ± 0,6 | 4,0 ± 0,9 | 0,030 ± 0,015 | 0,032 ± 0,012 | 0,020 ± 0,009 | 0,606 ± 0,051 | 0,551 ± 0,028 | 0,570 ± 0,056 |
| # kelompok dewasa | 111 | 383 | 80 | 111 | 383 | 80 | 23 | 104 | 15 | 23 | 104 | 15 | |||
| Rata-rata ± SE | 1,29 ± 0,06 | 1,44 ± 0,05 | 1,34 ± 0,01 | 198 ± 26 | 137 ± 33 | 103 ± 10 | 6,5 ± 0,8 | 6,5 ± 0,3 | 6,6 ± 1,2 | 0,036 ± 0,013 | 0,033 ± 0,006 | 0,041 ± 0,025 | |||
Catatan: Huruf subskrip yang berbeda menunjukkan perbedaan yang signifikan secara statistik antara populasi untuk data yang mencakup semua kelompok. Jarak antarkelompok dilaporkan sebagai data yang tidak ditransformasikan. * Proporsi remaja untuk setiap populasi adalah 0,082 ± 0,043, 0,098 ± 0,017, dan 0,161 ± 0,048 untuk populasi CS, EHA-BB, dan WHB.
Panen tahunan rata-rata, perubahan jangka pendek dalam lalu lintas kapal, dan jumlah paus dalam jarak 1 km dari kelompok muncul dalam model teratas untuk memprediksi ukuran kelompok ketika semua kelompok dipertimbangkan (Tabel S6 dan S7 ). Aktivitas kapal jangka panjang juga muncul dalam dua ΔAIC dari model peringkat teratas sebagai model terbaik keempat, tetapi dalam versi yang lebih kompleks dari model peringkat ketiga dengan sedikit daya prediksi tambahan. Oleh karena itu, kami menginterpretasikan model ini memiliki interpretabilitas biologis yang terbatas (Arnold 2010 ), dan dengan demikian aktivitas kapal tahunan jangka panjang bukan merupakan prediktor penting dari ukuran kelompok. Ukuran kelompok menurun dengan panen jangka panjang (Gambar 3b ) tetapi meningkat dengan meningkatnya tingkat aktivitas kapal jangka pendek dan dengan jumlah paus dalam radius 1 km (Gambar 3c,d ). Kombinasi aditif dari jumlah paus dalam jarak 1 km dan jenis kelompok adalah model teratas untuk memprediksi ukuran kelompok ketika kelompok dengan komposisi yang dikonfirmasi dipertimbangkan (Tabel S6 ). Ukuran kelompok meningkat seiring dengan jumlah paus dalam jarak 1 km dan lebih besar untuk kelompok dengan setidaknya satu paus muda (Gambar 4b,c ). Kombinasi interaktif jumlah paus dalam jarak 1 km dan jenis kelompok, serta aktivitas penangkapan ikan dan kapal jangka panjang juga muncul dalam model dalam jarak dua ΔAICc dari model peringkat teratas tetapi merupakan versi yang lebih kompleks dari model teratas dengan sedikit daya prediksi tambahan.

3.2 Komposisi Kelompok
Kelompok yang hanya menampilkan paus yang berhasil diklasifikasikan ke dalam kelas usia memiliki komposisi yang serupa di ketiga populasi, dengan 84%–92% merupakan paus dewasa ( χ 2 = 2,74, df = 2, p = 0,25). Kelompok dengan setidaknya satu paus muda juga memiliki komposisi yang serupa di antara ketiga populasi, dengan paus dewasa mewakili rata-rata 55%–61% dari suatu kelompok (Tabel 3 ). Model nol adalah model dengan peringkat teratas untuk memprediksi proporsi beluga dewasa dalam kelompok dengan setidaknya satu paus muda (Tabel S6 ).
3.3 Kohesi Spasial
Distribusi IGD minimum berbeda di antara tiga populasi ( χ2 = 83,12, df = 2, p < 0,001) yang tampaknya lebih mencerminkan perbedaan dalam pengukuran IGD median daripada pengukuran rata-rata. Kelompok dalam populasi CS lebih kohesif secara spasial daripada populasi EHA-BB ( p < 0,001), sementara kelompok dalam WHB tidak menunjukkan perbedaan dalam kohesi spasial dengan populasi CS ( p = 0,38) dan kurang kohesif secara spasial daripada populasi EHA-BB ( p < 0,001). Kelompok yang mencakup setidaknya satu remaja umumnya memiliki IGD yang lebih pendek daripada kelompok dengan orang dewasa hanya dalam populasi CS dan EHA-BB tetapi umumnya lebih besar di WHB (Tabel 3 ).
Perubahan jangka pendek dalam lalu lintas kapal, ukuran kelompok, dan interaksi antara kedua variabel muncul dalam model peringkat teratas untuk memprediksi IGD kelompok minimum beluga menggunakan kedua set data (Tabel S6 dan S7 ). Kepadatan kelompok juga muncul dalam < 2 ΔAICc dari model peringkat teratas, tetapi dalam model yang merupakan versi yang lebih kompleks dari model teratas dengan sedikit daya prediksi tambahan. Mengingat ukuran sampel yang lebih besar dari set data yang mempertimbangkan semua kelompok, kami menginterpretasikan hasil model menggunakan set data ini untuk memprediksi IGD minimum. IGD minimum menurun dengan perubahan jangka pendek dalam aktivitas kapal, sementara ada beberapa ketidakpastian seputar perbedaan dalam IGD minimum untuk kelompok dengan ukuran yang berbeda (Gambar 5b,c ). IGD minimum menurun dengan perubahan jangka pendek dalam aktivitas kapal untuk ketiga kelompok; namun, hubungannya secara signifikan kurang negatif pada kelompok tiga atau lebih beluga dibandingkan dengan paus tunggal (Gambar 5a ).

3.4 Kohesi Kelompok Sosial
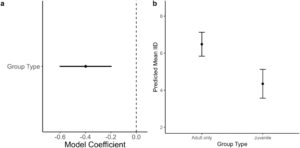
Rata-rata IID berbeda di antara ketiga populasi ( χ2 = 8,37, df = 2, p = 0,02 ) . Beluga dari populasi EHA-BB membentuk unit yang kurang kohesif secara spasial daripada populasi WHB ( p = 0,01), sementara populasi CS tidak menunjukkan perbedaan dengan populasi EHA-BB ( p = 0,99) dan WHB ( p = 0,21) (Tabel 3 ). Model nol termasuk di antara model teratas untuk memprediksi rata-rata IID ketika semua kelompok dipertimbangkan. Ketika hanya mempertimbangkan kelompok dengan komposisi yang dikonfirmasi, tipe kelompok muncul dalam model teratas untuk memprediksi rata-rata IID, dengan kelompok yang lebih kohesif secara spasial diamati dalam kelompok dengan setidaknya satu juvenil (Gambar 6 ; Tabel S6 dan S7 ). Kombinasi aditif dari tipe kelompok, panen per kapita, dan perubahan jangka pendek dalam lalu lintas kapal semuanya muncul dalam dua ΔAICc dari model peringkat teratas tetapi merupakan versi yang lebih kompleks dari model teratas dengan sedikit daya prediksi tambahan.
Varians orientasi kelompok sosial berbeda di antara ketiga populasi ( χ2 = 8,44, df = 2, p = 0,02). Varians orientasi kelompok rendah di ketiga populasi secara rata-rata, dengan hanya WHB (0,026 ± 0,007) yang memiliki varians kelompok lebih kecil daripada populasi EHA-BB ( p = 0,02), sementara varians orientasi kelompok dalam populasi CS mirip dengan populasi EHA-BB ( p = 0,28) dan WHB ( p = 0,99). Varians orientasi kelompok juga rendah untuk kelompok yang hanya mencakup sedikitnya satu remaja dan dewasa (Tabel 3 ). Model nol termasuk di antara model teratas untuk memprediksi varians orientasi renang kelompok beluga untuk semua kelompok dan untuk kelompok dengan komposisi kelompok yang dikonfirmasi (Tabel S6 ).
4 Diskusi
4.1 Karakteristik Kelompok
4.1.1 Ukuran Kelompok
Mirip dengan sebagian besar hewan yang hidup berkelompok (Reiczigel et al. 2008 ), beluga cenderung menyendiri atau dalam kelompok kecil di ketiga populasi. Ukuran kelompok rata-rata yang serupa dilaporkan untuk beluga EHA-BB dari survei udara visual (Innes et al. 2002 ), tetapi ukuran kelompok yang kami amati untuk beluga di Sungai Churchill lebih kecil daripada yang dilaporkan sebelumnya dalam survei udara fotografi di mana paus terlihat dalam kelompok yang rata-rata terdiri dari empat ekor (Matthews et al. 2017 ). Namun, pengamatan ini dilakukan selama beberapa hari tanpa definisi ukuran kelompok yang dilaporkan. Ryan et al. ( 2022 ) juga melaporkan ukuran kelompok rata-rata yang lebih besar pada beluga CS yang dikumpulkan dari drone dengan definisi ukuran kelompok yang serupa; namun, pengamatan bersifat oportunistik dan survei diterbangkan dari perahu di dekatnya. Sementara perbedaan ukuran kelompok ini dari platform fotografi yang berbeda dapat menunjukkan pergeseran temporal dalam perilaku pengelompokan beluga, hal itu lebih mungkin berasal dari perbedaan dalam metode survei atau definisi kelompok. Beluga terdeteksi lebih dalam di kolom air dalam citra drone (resolusi sekitar 3 cm) dibandingkan dalam citra satelit VHR 0,31 m (Stewart et al. 2024 ) dan dapat menyebabkan ukuran kelompok yang lebih kecil terdeteksi dalam citra VHR. Dengan citra satelit VHR, kami dapat mengamati sejumlah besar kelompok secara non-invasif secara bersamaan, yang menghilangkan faktor pengganggu potensial seperti gangguan perahu atau perubahan perilaku pengelompokan di berbagai hari dan waktu. Namun, pengamatan kami terbatas pada satu konteks, yang membatasi kemampuan kami untuk menafsirkan struktur sosial ini, yang juga dapat menjadi masalah dalam survei udara.
4.1.2 Komposisi Kelompok
Kelompok sebagian besar terdiri dari paus dewasa dengan sedikit juvenil di ketiga populasi (8%–16% juvenil; Tabel 3 ). Proporsi juvenil yang dikonfirmasi dalam setiap area musim panas adalah antara 5% dan 18%, yang lebih rendah dari proporsi paus muda yang disarankan dalam populasi beluga yang stabil (Taylor et al. 2007 ). Sementara hasil ini konsisten dengan penurunan yang dilaporkan untuk populasi CS (COSEWIC 2020 ), populasi EHA-BB dan WHB dianggap stabil (COSEWIC 2020 ). Oleh karena itu, hasil ini kemungkinan besar disebabkan oleh keterbatasan resolusi yang memengaruhi kemampuan untuk mengidentifikasi beluga juvenil dengan yakin, yang akan menunjukkan resolusi citra VHR saat ini mungkin terlalu rendah untuk menggambarkan komposisi kelompok beluga dengan yakin. Rata-rata, beluga dewasa terdiri sekitar 50%–60% dari anggota kelompok untuk kelompok yang menampilkan setidaknya satu juvenil di ketiga populasi (Tabel 3 ). Hal ini menunjukkan bahwa untuk kelompok yang kami amati, seekor beluga remaja pada umumnya ditemani oleh satu atau lebih beluga dewasa dan menunjukkan mungkin ada perbedaan minimal dalam prevalensi alokasi di antara populasi penelitian kami.
4.1.3 Jarak Antar Kelompok
IGD mengukur derajat kohesi spasial antara kelompok sosial beluga dan dapat menunjukkan variasi dalam tingkat perilaku fisi-fusi di seluruh populasi. Dinamika fisi-fusi pada odontocetes dianggap spesifik konteks, dengan ukuran dan komposisi kelompok berubah tergantung pada tuntutan perilaku langsung, seperti bersosialisasi atau mencari makan (Lunardi dan Ferreira 2014 ; O’Corry-Crowe et al. 2020 ). Menjaga jarak yang lebih pendek antara kelompok sosial dapat memfasilitasi penggabungan kelompok dan pemeliharaan hubungan sosial dengan individu yang terkait dan tidak terkait dalam kelompok tetangga (O’Corry-Crowe et al. 2020 ). Oleh karena itu, hasil kami menunjukkan bahwa beluga EHA-BB mungkin paling terlibat dalam pemisahan dan pembentukan kelompok dan mengadopsi struktur sosial yang berpotensi berbeda dibandingkan dengan beluga CS dan WHB. Ketiga area studi kami merupakan habitat penting bagi pemeliharaan anak beluga, sosialisasi, dan pada tingkat yang lebih rendah, mencari makan (Kilabuk 1998 ; Koski et al. 2002 ; Smith et al. 2017 ), sehingga kecil kemungkinan bahwa perbedaan dalam IGD rata-rata berasal dari variasi perilaku sosial atau mencari makan. Namun, perbedaan ini dapat disebabkan oleh variasi stresor lingkungan, yang kami bahas di bawah ini (lihat Bagian 4.2.1 ).
4.1.4 Jarak Antar Individu dan Varian Orientasi
Rata-rata IID kelompok untuk ketiga populasi melebihi panjang rata-rata beluga di setiap populasi (Luque dan Ferguson 2006 ). Ini menunjukkan jarak ideal antara beluga dalam kelompok mungkin lebih besar daripada yang secara hipotetis diperlukan berdasarkan ukurannya, mungkin memungkinkan pergerakan yang lebih mudah sambil mempertahankan kohesi kelompok. Kelompok WHB lebih kohesif secara sosial daripada kelompok EHA-BB, yang menunjukkan perbedaan potensial dalam organisasi spasial kelompok dalam dua populasi. Atau, variasi dalam kohesi sosial dapat mencerminkan perilaku rata-rata beluga di kedua populasi. Lumba-lumba Guyana ( Sotalia guianensis ), misalnya, memiliki IID yang lebih kecil saat beristirahat, bersosialisasi, dan bepergian di daerah dengan risiko predasi yang lebih besar, dan memiliki IID yang lebih besar saat menggiling atau makan (Lunardi dan Ferreira 2014 ). Jika hubungan serupa antara kohesi sosial dan perilaku ada pada beluga, hasil kami menunjukkan bahwa air payau yang keruh di Sungai Churchill memfasilitasi kepadatan yang lebih tinggi dari beluga yang beristirahat dan bersosialisasi. Beluga WHB juga mungkin lebih berisiko dimangsa paus pembunuh karena pecahnya es lebih awal di Selat Hudson memungkinkan akses lebih lama ke beluga WHB dibandingkan dengan beluga EHA-BB, yang dapat menjelaskan IID mereka yang lebih kecil (Ferguson et al. 2010 ). Sebaliknya, Teluk Creswell mungkin menawarkan peluang mencari makan yang lebih baik bagi beluga dibandingkan dengan Sungai Churchill, meskipun makan tidak dianggap menonjol di Teluk Creswell (Koski et al. 2002 ). Teluk Creswell jauh lebih besar daripada area survei Sungai Churchill (sekitar 1100 km 2 vs. 30 km 2 ) dan kemungkinan memfasilitasi rentang perilaku dan ukuran IID yang lebih besar, yang juga dapat menjelaskan variasi ini. Rata-rata, varians orientasi dalam kelompok di ketiga populasi itu rendah, yang menunjukkan anggota kelompok umumnya memiliki orientasi yang sama dan tingkat kohesi renang yang tinggi.
4.2 Gangguan Antropogenik
4.2.1 Aktivitas Kapal
Peningkatan ukuran kelompok dan penurunan IGD minimum yang diamati dengan perubahan jangka pendek dalam aktivitas kapal tahunan selaras dengan prediksi kami dan menunjukkan bahwa beluga yang mengalami peningkatan tingkat aktivitas lalu lintas kapal jangka pendek mungkin cenderung membentuk kelompok yang lebih besar yang menjaga jarak yang lebih dekat dengan kelompok tetangga. Dari perspektif adaptif, ini dapat menunjukkan bahwa kelompok yang lebih besar yang memiliki kohesi spasial yang lebih besar mungkin bermanfaat untuk deteksi dini gangguan kapal. Atau, ini dapat menunjukkan respons untuk mempertahankan komunikasi antarkelompok dalam lanskap suara yang lebih bising yang disebabkan oleh peningkatan aktivitas kapal. Paparan kapal dapat menjadi pemicu stres utama bagi beberapa spesies laut dalam konteks perilaku dan spasial tertentu (Lemos et al. 2022 ; Rolland et al. 2012 ; Watt et al. 2021 ; Wright, Soto, et al. 2007 ). Dengan demikian, hasil yang diamati dapat mengindikasikan peningkatan stres pada populasi yang terkait dengan peningkatan cepat lalu lintas kapal di seluruh distribusinya, seperti EHA-BB dan WHB, karena hewan sosial dapat meningkatkan ukuran kelompok atau kohesi sebagai respons stres (Culbert et al. 2019 ; DeVries et al. 2003 ). Tabrakan dengan kapal yang lebih besar diperkirakan tidak menjadi perhatian utama bagi paus yang lebih kecil dan lebih lincah seperti beluga (Lesage 2021 ; Martins et al. 2022 ), tetapi tabrakan mungkin menjadi perhatian bagi kapal pesiar yang lebih kecil yang tidak termasuk dalam penelitian ini (Huddart dan Stott 2020 ). Polusi suara kapal berpotensi mengganggu interaksi sosial beluga dan komunikasi topeng (Vergara et al. 2021 ), dengan beluga berpotensi dapat mendeteksi frekuensi kebisingan kapal hingga ~80 km jauhnya (Martin et al. 2022 ). Sementara gangguan yang disebabkan oleh kapal pada interaksi sosial beluga dapat berlangsung singkat (misalnya, Nowacek et al. 2007 ; Thompson et al. 2013 ), ada bukti perubahan jangka panjang pada perilaku cetacea dan ukuran kelompok sosial sehubungan dengan gangguan kapal (Bejder et al. 2006 ; Koper et al. 2016 ; Lusseau 2005 ; Parks et al. 2007 ). Saat ini tidak ada pengukuran langsung dari respons stres beluga terhadap lalu lintas kapal; namun, ada bukti stres yang terkait dengan kapal pada cetacea lain (Rolland et al. 2012 ), termasuk narwhal, yang ditemukan memiliki kadar kortisol yang lebih tinggi selama periode lalu lintas kapal yang tinggi dibandingkan dengan tahun-tahun sebelum gangguan (Watt et al. 2021 ).
4.2.2 Panen
Ukuran kelompok memiliki hubungan negatif dengan panen jangka panjang, yang menunjukkan bahwa populasi beluga yang mengalami tingkat panen yang lebih besar, seperti populasi CS, mungkin ada dalam kelompok sosial yang lebih kecil secara rata-rata atau memiliki proporsi paus tunggal yang lebih besar. Hasil ini tampaknya bertentangan dengan apa yang telah ditemukan pada paus pembunuh dan paus sperma, di mana terdapat bukti peningkatan perilaku pengelompokan setelah panen kumulatif untuk mempertahankan layanan sosial, seperti mempertahankan ukuran kelompok yang cukup untuk membesarkan anak-anak atau perlindungan dari predator (Busson et al. 2019 ; Whitehead et al. 2012 ). Dinamika kelompok pada spesies ini dianggap lebih didorong oleh matrilineal daripada pada beluga (O’Corry-Crowe et al. 2020 ), dan mengingat dinamika sosial fisi-fusi mereka, layanan sosial beluga mungkin sebaliknya dipertahankan melalui pembentukan beberapa pengelompokan seiring waktu daripada ukuran satu pengelompokan pada saat tertentu. Hasil ini tidak menyarankan respons perilaku beluga untuk meningkatkan kewaspadaan terhadap potensi ancaman panen, meskipun, pembentukan kelompok yang lebih kecil mungkin bermanfaat bagi beluga jika pemburu secara aktif berusaha untuk memanen kelompok yang lebih besar. Misalnya, beluga dipanen dari ukuran kelompok rata-rata empat dan lima dari pemburu di Inuvialuit Settlement, wilayah NU pada tahun 2014 dan 2015 (Ostertag et al. 2019 ). Namun, pemburu juga dapat memanen secara oportunistik tanpa preferensi untuk ukuran kelompok (Ostertag et al. 2018 ). Atau, hasil ini bisa jadi merupakan produk dari tekanan panen. Misalnya, mengingat bahwa panen beluga secara langsung memengaruhi jumlah individu dalam populasi, ukuran kelompok yang lebih kecil dapat menjadi hasil dari penurunan populasi beluga terkait panen (Bearzi et al. 2005 ), atau penghilangan individu sosial utama (Haber 1996 ; Lusseau dan Newman 2004 ; Williams dan Lusseau 2006 ). Populasi yang menurun biasanya memiliki komposisi populasi yang “sangat padat” dengan proporsi individu dewasa yang lebih besar daripada yang terlihat pada populasi yang stabil atau berkembang (Taylor et al. 2007 ). Mayoritas (67%) paus tunggal yang kami amati diklasifikasikan sebagai paus dewasa dan kemungkinan besar adalah paus jantan, karena setelah mandiri, paus beluga jantan dewasa dapat meninggalkan kelompok yang usianya bercampur dan tetap menyendiri atau bergaul dengan paus jantan lain (Colbeck et al. 2013 ; Mayette et al. 2023 ; O’Corry-Crowe et al. 2020). Namun, beluga tunggal juga bisa menjadi jantan muda yang lebih tua. Oleh karena itu, populasi beluga yang menurun cenderung menampilkan proporsi paus tunggal yang lebih besar daripada populasi yang stabil. Ini dapat menjelaskan ukuran kelompok rata-rata yang lebih kecil dan proporsi paus tunggal yang lebih besar di Cumberland Sound, karena beluga CS adalah satu-satunya populasi studi kami yang menghadapi penurunan kelimpahan yang terutama dikaitkan dengan pemanenan komersial yang berlebihan (COSEWIC 2020 ; Watt et al. 2020 ). Pemanenan juga secara instan mengubah komposisi kelompok dan jaringan sosial di mana penghilangan individu sosial utama, yang dapat bertindak sebagai penghubung dalam atau antara kelompok sosial, dapat menyebabkan kelompok yang terfragmentasi dan lebih kecil secara keseluruhan (Haber 1996 ; Lusseau dan Newman 2004 ; Williams dan Lusseau 2006 ). Efek ini kemungkinan lebih menonjol pada populasi yang lebih kecil, yang sekali lagi dapat menjelaskan ukuran kelompok yang lebih kecil yang kami amati pada populasi CS yang merupakan populasi studi kami yang terkecil (COSEWIC 2020 ).
4.3 Kepadatan
Bertentangan dengan prediksi kami, ukuran kelompok meningkat dengan jumlah paus dalam radius 1 km. Mengingat suara umumnya merambat lebih sedikit di air dangkal dibandingkan dengan air yang lebih dalam (Liang et al. 2014 ), kebisingan dari paus sejenis mungkin tidak memengaruhi ukuran kelompok beluga sejauh yang kami prediksi sebelumnya di habitat musim panas yang relatif dangkal tempat pengamatan kami dikumpulkan. Sebaliknya, ada kemungkinan bahwa area dengan kepadatan tinggi merupakan indikasi habitat yang lebih kondusif untuk peningkatan aktivitas sosial, seperti di air dangkal yang lebih dekat ke pantai tempat beluga lebih terlindungi dari ancaman pemangsaan musim panas dari paus pembunuh (Frost dan Lowry 1990 ; Westdal et al. 2016 ). Misalnya, kelompok beluga EBS muda dan campuran usia lebih mungkin ditemukan di habitat perairan dangkal dibandingkan dengan perairan yang lebih dalam (Loseto et al. 2006 ; Mayette et al. 2023 ), meskipun segregasi usia dalam kaitannya dengan habitat tidak ditemukan menonjol di beluga WHB (Westdal et al. 2022 ). Namun, hasil ini tampaknya tidak sejalan dengan perilaku mencari makan khas hewan yang hidup berkelompok, yang umumnya membentuk kelompok yang lebih kecil di daerah dengan kepadatan tinggi untuk memaksimalkan asupan energi (Hintz dan Lonzarich 2018 ; Webber dan Vander Wal 2021 ). Kelompok yang lebih besar mungkin bermanfaat untuk mencari makan secara kooperatif atau menemukan mangsa jika spesies mangsa langka (Thompson et al. 1974 ); sayangnya, data kelimpahan mangsa terbatas untuk area studi kami. Karakteristik kelompok sosial beluga dalam kaitannya dengan fungsi fitur habitat sebagai area dengan aktivitas sosial yang meningkat, tempat perlindungan predator, atau peluang mencari makan memerlukan pemeriksaan lebih lanjut.
4.4 Konteks Sosial
4.4.1 Ukuran Kelompok
Dibandingkan dengan beluga tunggal, IGD minimum kelompok dengan tiga atau lebih memiliki hubungan negatif yang signifikan dengan perubahan jangka pendek dalam aktivitas kapal. Ini sejalan dengan prediksi kami dan menunjukkan bahwa manfaat kohesi spasial kelompok mungkin tidak sepenting bagi kelompok beluga yang lebih besar daripada bagi paus tunggal. Seperti yang disebutkan sebelumnya, IGD minimum yang lebih pendek dapat meningkatkan kewaspadaan kawanan dan bermanfaat untuk deteksi dini gangguan kapal atau peningkatan komunikasi antarkelompok di lanskap suara yang lebih bising. Faktor-faktor ini mungkin sangat bermanfaat bagi paus tunggal yang mungkin memiliki kewaspadaan tingkat kelompok yang lebih rendah terhadap pemicu stres potensial (Krause dan Ruxton 2002 ).
4.4.2 Tipe Kelompok
Bahasa Indonesia: Allocare oleh kerabat dan non-kerabat konspesifik disarankan menjadi salah satu manfaat utama dari sosialitas pada beluga (Aubin et al. 2021 ), dan merupakan penjelasan yang mungkin untuk peningkatan ukuran kelompok yang diamati ketika ada juvenil. Selain itu, ketika beluga jantan dewasa menjadi lebih mandiri, mereka mungkin sering diamati sendirian (Colbeck et al. 2013 ; Mayette et al. 2023 ; O’Corry-Crowe et al. 2020 ), yang selanjutnya berkontribusi pada ukuran kelompok dewasa yang lebih kecil dibandingkan dengan kelompok usia campuran. Meningkatnya perilaku pengelompokan di hadapan juvenil bukanlah hal yang tidak biasa pada cetacea (Oremus et al. 2012 ), dan mungkin memberikan perlindungan beluga muda, pembelajaran sosial, dan peluang perawatan (Brodie 1971 ; Colbeck et al. 2013 ; Krasnova et al. 2009 ; Whitehead 2003 ). Akan tetapi, proporsi paus dewasa di hadapan paus-paus muda tidak diprediksi secara kuat oleh stresor antropogenik, yang menunjukkan bahwa perlindungan mungkin bukan pendorong yang kuat terhadap alokasi paus beluga muda.
Kelompok beluga campuran usia umumnya lebih kohesif secara sosial daripada kelompok yang hanya berisi beluga dewasa. Hasil serupa diamati pada beluga EBS (Mayette et al. 2022 ), dan lumba-lumba (Lunardi dan Ferreira 2014 ; Scott dan Perryman 1999 ), dan kemungkinan mencerminkan hubungan dekat antara beluga betina dewasa dan beluga muda (Brodie 1971 ; Colbeck et al. 2013 ). Beluga dewasa yang lebih besar mungkin juga memerlukan jarak yang lebih jauh antara anggota kelompok daripada beluga muda untuk memfasilitasi gerakan berenang, sekaligus menjaga kekompakan kelompok.
4.5 Keterbatasan
Citra satelit menguntungkan untuk pemantauan beluga karena menawarkan snapshot non-invasif dari satu contoh waktu dari mana kita dapat membuat pengamatan spesies yang sangat mobile yang bergerak dalam tiga dimensi. Namun, ada kemungkinan bahwa karakteristik kelompok beluga berubah sepanjang hari, bulan, dan tahun yang tidak ditangkap, dan mirip dengan metode udara lainnya, ada potensi bagi individu yang menyelam dalam kelompok untuk terlewatkan. Semua pengamatan kami dilakukan dari populasi beluga pada saat-saat tunggal dalam habitat musim panas mereka pada waktu yang hampir sama dalam sehari. Oleh karena itu, kami berasumsi bahwa perbandingan antara populasi relevan. Individu yang menyelam dapat dikoreksi untuk kemungkinan mendapatkan ukuran kelompok yang lebih akurat (Stewart et al. 2024 ), meskipun, individu yang menyelam cukup dalam di kolom air untuk menghindari deteksi pengamat dapat dianggap eksternal terhadap kelompok sosial. Selain itu, sifat statis citra membatasi interpretasi fungsi karakteristik kelompok sosial. Misalnya, perilaku kelompok beluga dengan orientasi kelompok yang bervariasi tidak sepenuhnya jelas. Analisis di masa mendatang akan lebih baik dari pengamatan perilaku yang tidak mencolok, misalnya, dari darat atau dari survei video drone di ketinggian tinggi, untuk membantu interpretasi kelompok sosial.
Resolusi citra satelit VHR kami berarti kami hanya dapat memperkirakan kelas usia paus individu berdasarkan ukuran. Contoh perkiraan panjang yang meyakinkan kemungkinan lebih rendah di WHB (54% vs. 62%–68% untuk dua populasi lainnya) karena sebagian besar beluga diamati di perairan keruh di dalam Sungai Churchill daripada di aliran sungai keruh di dalam Clearwater Fiord dan Teluk Creswell (Gambar 2 ). Jumlah paus yang dapat diklasifikasikan berdasarkan usia lebih rendah di ketiga populasi studi kami dibandingkan dengan klasifikasi usia beluga EBS dari survei foto udara di mana kelas usia yang sama dapat diperkirakan untuk minimal 85% beluga (Mayette et al. 2022 ), dan kemungkinan merupakan hasil dari resolusi yang lebih rendah dalam citra satelit VHR. Hal ini membuat kami memiliki sejumlah kelompok yang terbatas untuk mengeksplorasi bagaimana karakteristik kelompok berbeda dengan keberadaan paus muda. Lebih jauh, resolusi citra yang lebih rendah dapat membuat pembedaan antara beluga dan narwhal dan anjing laut yang berukuran dan berwarna serupa menjadi tantangan. Meskipun tidak diharapkan berdampak signifikan pada studi ini, kehati-hatian diperlukan untuk mendeteksi beluga dalam citra VHR di area tempat spesies ini tumpang tindih. Meskipun demikian, studi ini memberikan aplikasi baru citra satelit VHR untuk pemantauan beluga dan estimasi komposisi kelompok. Kemampuan satelit VHR diproyeksikan akan terus meningkat dengan resolusi serendah 10 cm yang diharapkan dalam beberapa tahun mendatang (Khan et al. 2023 ), yang akan memungkinkan pengukuran yang lebih efektif untuk semua beluga di masa mendatang.
4.6 Kesimpulan
Kami menemukan perbedaan dalam karakteristik kelompok antara tiga populasi beluga yang berbeda dan hubungan antara karakteristik ini dengan panen tahunan jangka panjang dan perubahan jangka pendek dalam aktivitas kapal. Meskipun penting untuk dicatat bahwa hasil kami tidak menyiratkan sebab akibat, hubungan ini mungkin memiliki beberapa implikasi untuk populasi beluga. Populasi beluga yang telah mengalami panen per kapita tahunan lebih banyak memiliki kelompok yang lebih kecil secara rata-rata. Pembentukan kelompok yang lebih kecil secara terus-menerus dapat mengakibatkan berkurangnya alokasi dan perlindungan anak sapi, dan lebih sedikit peluang untuk transfer budaya atau pengetahuan, yang dianggap bermanfaat bagi paus untuk mengeksploitasi sumber daya mangsa atau menghindari pemicu stres seperti jebakan es, terdampar di air surut, atau pemangsaan (Wade et al. 2012 ). CS memiliki tingkat panen per kapita rata-rata jangka panjang tertinggi dan ukuran kelompok rata-rata terkecil dan merupakan satu-satunya populasi studi kami yang terdaftar sebagai terancam punah (COSEWIC 2020 ). Status ini terutama dikaitkan dengan pemanenan berlebihan secara historis dan kurangnya pemulihan populasi (COSEWIC 2020 ); Namun, alasan kurangnya pemulihan sulit untuk dipastikan. Penjelasan yang mungkin adalah tingginya tingkat sosialitas pada populasi beluga dan degradasi layanan sosial yang disebabkan oleh perburuan besar-besaran dan pengurangan ukuran kelompok sosial berikutnya (Wade et al. 2012 ). Pertimbangan tentang pengaruh perburuan pada dinamika sosial beluga mungkin penting untuk menerapkan strategi pengelolaan di masa mendatang.
Ukuran kelompok dan kohesi spasial terbesar pada beluga EHA-BB dan terkait dengan peningkatan aktivitas kapal baru-baru ini, yang semuanya merupakan fitur yang serupa dengan yang sebelumnya dikaitkan dengan penggambaran stres yang meningkat pada hewan yang hidup berkelompok (Culbert et al. 2019 ; DeVries et al. 2003 ). Namun, ini spekulatif, karena saat ini tidak ada bukti langsung stres yang disebabkan oleh kapal pada beluga. Stres kronis dapat memiliki banyak implikasi bagi beluga, terutama pada parameter riwayat hidup seperti tingkat reproduksi dan kerentanan penyakit (Wright, Deak, et al. 2007 ). Kapal pesiar kecil juga dianggap memengaruhi beluga secara negatif pada beberapa populasi (Blane dan Jaakson 1994 ), tetapi kami tidak dapat memasukkan aktivitas kapal pesiar kecil dalam studi ini. Lalu lintas kapal diproyeksikan meningkat di Arktik (Stephenson et al. 2013 ) dan mungkin menjadi perhatian khusus bagi beluga dalam populasi EHA-BB, yang mengalami peningkatan terbesar dalam aktivitas kapal baru-baru ini. Menggabungkan dampak perahu yang lebih kecil ke dalam studi masa depan dan terus memantau hubungan antara stres beluga dan aktivitas kapal besar, mungkin melalui perubahan perilaku pengelompokan, memerlukan penelitian lebih lanjut.
Kami menggunakan citra satelit VHR untuk menyoroti kompleksitas kelompok sosial beluga dan menunjukkan bahwa kombinasi gangguan antropogenik, kepadatan, dan konteks sosial membentuk karakteristik kelompok pada beluga. Kami memberikan wawasan pertama tentang karakteristik kelompok populasi beluga CS, EHA-BB, dan WHB sehubungan dengan stresor antropogenik; namun, penelitian lanjutan untuk lebih memahami pengaruh stresor ini pada dinamika kelompok sosial beluga diperlukan. Mengingat pentingnya sosialitas bagi beluga, upaya pengelolaan di masa mendatang dapat memperoleh manfaat dari penilaian berkelanjutan terhadap dinamika kelompok sosial beluga di samping upaya pemantauan tradisional, terutama mengingat perluasan stresor antropogenik di Arktik.


