
ABSTRAK
Model distribusi spesies (SDM) telah dikembangkan untuk beberapa spesies cetacea dalam Ekosistem Arus California (CCE) dari data survei di atas kapal yang dikumpulkan oleh Southwest Fisheries Science Center (SWFSC) pada musim panas dan gugur, sehingga membatasi kemampuan untuk menginformasikan keputusan pengelolaan di musim dingin ketika kelimpahan dan pola distribusi sangat berbeda. SDM musim dingin dan musim semi telah dikembangkan untuk beberapa spesies menggunakan data survei di atas kapal California Cooperative Oceanic Fisheries Investigations (CalCOFI), tetapi prediksi model terbatas pada perairan di lepas pantai California selatan dan tengah. Dalam studi ini, estimasi kepadatan musim dingin dan musim semi untuk seluruh area studi CCE dibuat dari prediksi model musim panas dan gugur SWFSC (ekstrapolasi temporal) dan prediksi model musim dingin dan musim semi CalCOFI (ekstrapolasi spasial) untuk lumba-lumba biasa berparuh pendek ( Delphinus delphis delphis ), lumba-lumba sisi putih Pasifik ( Lagenorhynchus obliquidens ), lumba-lumba Dall ( Phocoenoides dalli ), dan paus sirip ( Balaenoptera physalus ). Kinerja model dibandingkan berdasarkan estimasi kelimpahan yang tersedia dan pola distribusi yang terdokumentasi di musim dingin. Hasil mengungkapkan faktor ekologi spesifik spesies yang perlu dipertimbangkan saat mengekstrapolasi prediksi model secara temporal atau spasial, termasuk apakah area studi tertentu mencakup “habitat inti” spesies, dan apakah variabel statis harus disertakan saat spesies menunjukkan pergeseran distribusi temporal.
1 Pendahuluan
Pengelolaan dan langkah-langkah konservasi spesies laut yang efektif memerlukan prediksi kelimpahan dan distribusi spesies yang eksplisit secara spasial dan temporal. Model distribusi spesies (SDM) telah ditetapkan sebagai alat penting untuk konservasi dan pengelolaan laut karena dapat digunakan untuk memprediksi kelimpahan dan pola distribusi burung laut, ikan, penyu laut, cetacea, dan spesies lainnya (Abrahms et al. 2019 ; Becker et al. 2016 , 2022 ; Eguchi et al. 2017 ; Gilles et al. 2011 ; Hammond et al. 2013 ; Hazen et al. 2017 , 2018 ; Louzao et al. 2006 ; Oppel et al. 2012 ; Redfern et al. 2013 ; Torres et al. 2015 ; Welch et al. 2019 ). Model kepadatan berbasis habitat, yang mengintegrasikan data lingkungan ke dalam SDM, sangat berguna dalam lingkungan laut di mana banyak spesies menunjukkan perubahan dinamis dalam kelimpahan dan distribusi sebagai respons terhadap perubahan kondisi laut dalam suatu wilayah tertentu (Becker et al. 2014 , 2017 , 2018 ; Cañadas dan Hammond 2008 ; Gilles et al. 2016 ; Roberts et al. 2016 ).
Idealnya, SDM akan dikembangkan berdasarkan data yang spesifik untuk setiap area dan musim yang diminati; namun, kendala finansial dan logistik sering kali membatasi pengumpulan data ke sebagian wilayah atau periode waktu. Ekstrapolasi prediksi model, baik secara spasial maupun temporal, mungkin diperlukan untuk mendukung keputusan pengelolaan ketika tidak ada alternatif yang tersedia (Mannocci et al. 2015 ; Roberts et al. 2016 ; Wang et al. 2021 ). Prediksi yang diekstrapolasi tersebut, ketika diterapkan dengan hati-hati dari SDM yang dikembangkan dengan solid, sering kali dianggap lebih unggul daripada tidak memiliki informasi untuk mendukung keputusan konservasi atau pengelolaan (Bouchet et al. 2019 ; Forbes dan Calow 2002 ; Mannocci et al. 2017 ; Miller et al. 2004 ).
Seiring dengan pertumbuhan populasi manusia dan dampaknya pada spesies laut, tuntutan konservasi kemungkinan akan meningkatkan minat dalam mengekstrapolasi SDM ke wilayah dan periode waktu baru; namun, transferabilitas model hanya dinilai untuk beberapa spesies laut (misalnya, Mannocci et al. 2017 ; Redfern et al. 2017 ; Torres et al. 2015 ). Torres et al. ( 2015 ) mengevaluasi prediksi ekstrapolasi penggunaan habitat burung petrel abu-abu ( Procellaria cinerea ) di seluruh Belahan Bumi Selatan dan menemukan bahwa dalam ekosistem baru, SDM dapat mengidentifikasi distribusi potensial (tempat suatu spesies dapat hidup) tetapi tidak dapat memprediksi distribusi yang terealisasi (tempat suatu spesies benar-benar berada relatif terhadap habitat yang tersedia). Redfern et al. ( 2017 ) mengevaluasi transferabilitas SDM paus biru ( Balaenoptera musculus ) dari dua ekosistem yang diambil sampelnya dengan baik ke ekosistem yang miskin data dan menemukan bahwa model khusus ekosistem tidak dapat ditransfer, sementara model yang dikembangkan menggunakan data dari dua ekosistem yang berbeda berkinerja lebih baik. Sebagian besar studi kelautan hingga saat ini berfokus pada ekstrapolasi dari wilayah yang disurvei dengan baik ke wilayah yang disurvei dengan buruk yang terpisah secara geografis satu sama lain, daripada ekstrapolasi temporal atau spasial dalam ekosistem tertentu (misalnya, Mannocci et al. 2015 ; Redfern et al. 2017 ). Dalam ekosistem yang menunjukkan variabilitas temporal dan spasial yang substansial dalam kondisi samudra, penting untuk menilai keakuratan ekstrapolasi temporal dan spasial, terutama jika ekstrapolasi diperlukan untuk memenuhi kebutuhan manajemen kritis.
Dalam Ekosistem Arus California (CCE), sejarah panjang penelitian telah mendokumentasikan perubahan dinamis dalam kelimpahan dan distribusi cetacea yang terkait dengan variabilitas musiman dan tahunan dalam kondisi lautan di seluruh wilayah (Becker et al. 2014 , 2017 , 2018 , 2020 ; Boyd et al. 2018 ; Campbell et al. 2015 ; Dohl et al. 1978 ; Douglas et al. 2014 ; Forney dan Barlow 1998 ; Hazen et al. 2017 ). SDM telah dikembangkan untuk banyak spesies cetacea di CCE menggunakan data survei sistematis yang dikumpulkan oleh Southwest Fisheries Science Center (SWFSC) di lepas Pantai Barat AS sejak 1991 (Barlow et al. 2009 ; Becker et al. 2010 , 2012 , 2016 , 2020 ; Forney 2000 ; Forney et al. 2012 ; Redfern et al. 2013 ). Mayoritas upaya survei SWFSC 1991–2018 (Gambar 1 ) telah dibatasi pada bulan-bulan musim panas dan gugur, karena kondisi cuaca selama musim dingin dan semi terlalu buruk untuk survei berbasis kapal. Prediksi kepadatan berdasarkan SDM yang dikembangkan dari data survei musim panas dan gugur telah digunakan untuk berbagai aplikasi manajemen, misalnya, untuk menilai risiko tabrakan kapal untuk paus besar (Redfern et al. 2013 , 2019 ; Rockwood et al. 2017 ); untuk mengevaluasi risiko kebisingan pengiriman kronis pada cetacea di California Selatan (Redfern et al. 2017 ); dan untuk mengevaluasi dampak potensial pada cetacea dari melakukan kegiatan pelatihan dan pengujian Angkatan Laut di laut (Becker et al. 2016 , 2018 , 2020 , 2022 ; Forney et al. 2012 ; Departemen Angkatan Laut AS 2015 , 2017 ). SDM ini memungkinkan penilaian aktivitas antropogenik selama musim panas dan gugur, tetapi dampak potensial selama musim dingin dan semi tidak ditangkap secara memadai. ( 2014 ) mengevaluasi ekstrapolasi musiman untuk tiga spesies cetacea dan menunjukkan bahwa ekstrapolasi tersebut dapat memberikan informasi untuk beberapa spesies, khususnya ketika rentang kovariat yang digunakan untuk membangun model tersebut serupa dengan yang diamati pada musim alternatif. Akan tetapi, penelitian tersebut juga menggambarkan risiko ekstrapolasi di luar ruang kovariat yang dimodelkan, yang menggarisbawahi perlunya penelitian lebih lanjut untuk menilai kapan dan bagaimana ekstrapolasi dapat dilakukan secara andal lintas ruang dan waktu.
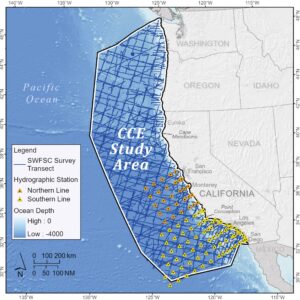
Dalam studi ini, kami membandingkan kinerja prediksi SDM cetacea yang diekstrapolasi secara temporal (dari musim panas dan gugur ke musim dingin dan semi) dengan yang diekstrapolasi secara spasial dalam satu musim dari satu bagian area studi CCE ke seluruh kawasan CCE. Untuk menyederhanakannya, “musim hangat” mengacu pada musim panas dan gugur sementara “musim dingin” mengacu pada musim dingin dan semi (Departemen Angkatan Laut AS 1977 ). Perbandingan kami menggunakan dua set survei cetacea: (1) survei musim panas dan gugur di seluruh CCE 1991–2018 yang dilakukan oleh SWFSC yang telah menjadi dasar untuk SDM yang dijelaskan dalam Becker et al. ( 2012 , 2014 , 2016 , 2020 ), dan (2) survei mamalia laut 2005–2020 yang dilakukan selama pelayaran triwulanan California Cooperative Oceanic Fisheries Investigations (CalCOFI) di lepas pantai California selatan dan tengah (Campbell et al. 2015 ; Douglas et al. 2014 ). Sebagian kecil data terakhir (2005 hingga 2015) sebelumnya digunakan untuk mengembangkan SDM musim dingin dan musim semi untuk lumba-lumba biasa berparuh pendek ( Delphinus delphis delphis ), lumba-lumba Dall ( Phocoenoides dalli ), dan paus bungkuk ( Megaptera novaeangliae ) (Becker et al. 2017 ). Hasil tersebut memberikan prediksi kepadatan eksplisit spasial pertama untuk spesies ini di lepas pantai California tengah dan selatan selama musim dingin dan mengonfirmasi bahwa pola kelimpahan dan distribusi sangat berbeda dari yang didokumentasikan untuk musim panas dan gugur. Namun, karena wilayah studi CalCOFI hanya mencakup sebagian wilayah studi CCE (Gambar 1 ), variabilitas musiman dalam kelimpahan spesies dan pola distribusi di utara 38° LU tidak tertangkap.
Analisis yang disajikan di bawah ini menguji apakah ekstrapolasi temporal atau spasial dapat digunakan untuk menyediakan SDM yang diselesaikan secara musiman untuk seluruh wilayah CCE, mengisi kesenjangan data yang semakin penting untuk pengelolaan aktivitas antropogenik di sepanjang Pantai Barat AS. Ukuran sampel cukup untuk membandingkan hasil model yang diekstrapolasi untuk empat spesies yang beragam secara taksonomi dan ekologis: lumba-lumba biasa berparuh pendek, lumba-lumba sisi putih Pasifik ( Lagenorhynchus obliquidens ), lumba-lumba Dall, dan paus sirip ( Balaenoptera physalus ).
1.1 Tinjauan Umum Spesies
Di bawah ini adalah ringkasan singkat informasi tentang apa yang saat ini diketahui tentang distribusi, kelimpahan, dan tren keempat spesies yang diperiksa, untuk memberikan konteks ekologi untuk analisis ini.
1.1.1 Lumba-lumba Biasa Berparuh Pendek
Selama akhir tahun 1970-an dan awal tahun 1980-an di sepanjang Pantai Barat AS, lumba-lumba biasa berparuh pendek terlihat terutama di selatan Point Conception (Dohl et al. 1983 ), tetapi sejak awal tahun 2000-an mereka umumnya ditemui sejauh utara 42° LU (Hamilton et al. 2009 ), dan kadang-kadang sejauh utara 48° LU (Forney 2007 ). Pergeseran distribusi musiman diucapkan, dengan pergeseran selatan yang signifikan ke selatan Point Arguello di musim dingin (Becker et al. 2014 ; Campbell et al. 2015 ; Forney dan Barlow 1998 ). Lumba-lumba biasa berparuh pendek adalah spesies beriklim hangat hingga tropis, dan berdasarkan model habitat yang dikembangkan menggunakan data survei garis transek di lepas Pantai Barat AS dari tahun 1991 hingga 2018, kepadatan paling besar terjadi saat perairan paling hangat (Barlow et al. 2009 ; Becker et al. 2010 , 2016 , 2014 , 2018 , 2020 ; Forney dan Barlow 1998 ). Barlow ( 2016 ) mencatat peningkatan yang hampir monoton dalam kelimpahan lumba-lumba biasa berparuh pendek dari tahun 1991 hingga 2014 di lepas Pantai Barat AS pada musim panas dan gugur. Prediksi dari model kepadatan berbasis habitat menunjukkan peningkatan serupa dalam kelimpahan lumba-lumba paruh pendek di musim panas dan gugur di lepas Pantai Barat AS dari tahun 1996 hingga 2018, dengan estimasi kelimpahan terbaru (2018) sebesar 1.056.308 (CV = 0,207) (Barlow 2016 ; Becker et al. 2020 ; Carretta et al. 2023 ). Namun, peningkatan kelimpahan lumba-lumba biasa berparuh pendek dapat disebabkan oleh pergerakan hewan ke utara dari perairan lepas pantai Meksiko, dan analisis tambahan diperlukan untuk menilai tren populasi potensial dengan lebih baik (Barlow 2016 ; Becker et al. 2022 ).
1.1.2 Lumba-lumba Sisi Putih Pasifik
Lumba-lumba sisi putih Pasifik ditemukan di perairan beriklim dingin di sepanjang tepi utara Samudra Pasifik sejauh utara Laut Bering selatan dan sejauh selatan Teluk California di lepas pantai Meksiko (Jefferson et al. 2015 ). Forney dan Barlow ( 1998 ) menemukan pergeseran utara/selatan yang signifikan dalam distribusi musiman lumba-lumba sisi putih Pasifik di lepas pantai California, dengan hewan bergerak ke utara ke perairan Oregon dan Washington selama musim panas dan menunjukkan peningkatan kelimpahan di Teluk California Selatan selama musim dingin. Selama kondisi air hangat yang tidak biasa terjadi pada tahun 2014, ada beberapa penampakan lumba-lumba sisi putih Pasifik di lepas pantai California tengah dan selatan (Barlow 2016 ). Ada variabilitas musiman dan tahunan yang cukup besar dalam kelimpahan dan distribusi lumba-lumba sisi putih Pasifik di lepas Pantai Barat AS, dan tidak ada tren populasi jangka panjang yang telah diidentifikasi (Carretta et al. 2023 ). Berdasarkan data yang dikumpulkan pada musim panas dan gugur, estimasi kelimpahan terbaru (2018) untuk lumba-lumba sisi putih Pasifik di lepas pantai California, Oregon, dan Washington adalah 34.999 (CV = 0,222) (Becker et al. 2020 ; Carretta et al. 2023 ).
1.1.3 Lumba-lumba Dall
Lumba-lumba Dall adalah salah satu spesies odontocete yang paling umum di perairan Pasifik Utara, yang terutama terdapat di antara 30° LU dan 62° LU, meskipun selama periode air yang luar biasa dingin mereka terdapat di selatan sejauh 28° LU (Jefferson et al. 2015 ). Distribusi lumba-lumba Dall di lepas Pantai Barat AS sangat bervariasi antar tahun, kemungkinan besar karena perubahan kondisi oseanografi (Barlow et al. 2009 ; Becker et al. 2016 , 2018 , 2020 ; Forney 2000 ; Forney et al. 2012 ). Pergerakan utara-selatan di California, Oregon, dan Washington telah diamati, dengan lumba-lumba Dall menggeser distribusinya ke selatan selama periode air yang lebih dingin pada skala waktu antartahunan dan musiman (Becker et al. 2014 , 2018 ; Boyd et al. 2018 ; Forney dan Barlow 1998 ). Berdasarkan model habitat yang dikembangkan menggunakan data survei 1991–2018 yang dikumpulkan di perairan lepas Pantai Barat AS selama musim panas dan gugur, kepadatan lumba-lumba Dall paling besar di perairan landas dan lereng, dan menurun secara substansial di perairan yang lebih hangat dari sekitar 17°C (Barlow et al. 2009 ; Becker et al. 2020 , 2016 , 2020 ; Forney et al. 2012 ). Selama survei kapal yang dilakukan setiap triwulan di lepas pantai California selatan dari tahun 2004 hingga 2008, lumba-lumba Dall ditemui sepanjang tahun, dengan pertemuan tertinggi selama bulan-bulan air dingin (Douglas et al. 2014 ). Mengingat variabilitas substansial dalam kelimpahan dan distribusi lumba-lumba Dall di lepas Pantai Barat AS, tidak ada tren populasi jangka panjang yang telah diidentifikasi (Carretta et al. 2023 ). Berdasarkan data yang dikumpulkan pada musim panas dan gugur, estimasi kelimpahan terbaru (2018) untuk lumba-lumba Dall di lepas pantai California, Oregon, dan Washington adalah 16.498 hewan (CV = 0,608) (Becker et al. 2020 ; Carretta et al. 2023 ).
1.1.4 Paus Sirip
Paus sirip menunjukkan pola pergerakan yang kompleks dalam Ekosistem Arus Kalifornia, dan tampaknya tidak mengikuti model migrasi paus balin yang umum. Pelacakan satelit menunjukkan bahwa pergerakan paus sirip di lepas Pantai Barat AS sangat bervariasi, menunjukkan pergerakan jarak jauh di sepanjang pantai serta perjalanan musiman pendek di musim semi dan gugur (Falcone dan Schorr 2014 ; Mate et al. 2015 ; Scales et al. 2017 ). Bahasa Indonesia: Di musim panas dan gugur, paus sirip terdistribusi secara luas dalam kepadatan yang relatif tinggi di lepas Pantai Barat AS, dengan agregasi paus sirip hadir sepanjang tahun di California selatan dan tengah (Barlow et al. 2009 ; Becker et al. 2016 , 2018 , 2020 ; Calambokidis et al. 2024 ; Campbell et al. 2015 ; Dohl et al. 1983 ; Douglas et al. 2014 ; Forney dan Barlow 1998 ; Scales et al. 2017 ). Penampakan dari survei sepanjang tahun di lepas pantai California selatan dari tahun 2004 hingga 2013 menunjukkan paus sirip lebih jauh ke lepas pantai di musim panas dan gugur dan lebih dekat ke pantai di musim dingin dan semi (Campbell et al. 2015 ; Douglas et al. 2014 ). Analisis data identifikasi foto jangka panjang menunjukkan bahwa dua populasi paus sirip yang tumpang tindih terjadi di lepas Pantai Barat AS, populasi sementara dengan pergerakan musiman yang luas yang berkisar dari Teluk California Selatan ke utara hingga perairan di lepas pantai Washington dan populasi penduduk sepanjang tahun di dalam Teluk California Selatan yang menunjukkan pergeseran musiman di dekat pantai dan lepas pantai dalam distribusi (Falcone et al. 2022 ). Analisis data survei garis-transek yang dikumpulkan selama musim panas dan gugur antara tahun 1991 dan 2018, termasuk estimasi berbasis desain, model kepadatan berbasis habitat, dan analisis tren Bayesian, menunjukkan bahwa kelimpahan paus sirip di Arus California meningkat selama periode ini (Becker et al. 2020 ; Moore dan Barlow 2011 ; Nadeem et al. 2016 ). Berdasarkan data yang dikumpulkan pada musim panas dan gugur, estimasi kelimpahan terbaru (2018) untuk paus sirip di lepas pantai California, Oregon, dan Washington adalah 11.065 (CV = 0,405) (Becker et al. 2020 ; Carretta et al. 2023 ).
2 Bahan dan Metode
2.1 Wilayah Penelitian dan Metode Lapangan
Area studi Ekosistem Arus California (CCE) mencakup perairan di lepas Pantai Barat AS hingga sekitar 555 km lepas pantai, sementara area studi CalCOFI kira-kira sepertiga ukuran area studi CCE, meliputi 11 garis transek yang membentang kira-kira tegak lurus ke pantai antara stasiun pengambilan sampel di lepas pantai selatan dan tengah California (Gambar 1 ). Data penampakan cetacea yang dikumpulkan oleh Southwest Fisheries Science Center (SWFSC) selama survei kapal sistematis di perairan CCE dari tahun 1991 hingga 2018 (Tabel 1 ) digunakan untuk mengembangkan SDM musim panas dan gugur, mengikuti metode Becker et al. ( 2020 ), tetapi dengan serangkaian kovariat yang berbeda, seperti dijelaskan di bawah ini. SDM musim dingin dan semi dikembangkan menggunakan data penampakan cetacea yang dikumpulkan selama pelayaran CalCOFI triwulanan yang dilakukan dari tahun 2005 hingga 2020 (Tabel 2 ), mengikuti metode Becker et al. ( 2017 ), lagi-lagi dengan serangkaian kovariat yang berbeda. Sementara ada beberapa variasi temporal dalam pengaturan waktu pelayaran, survei musim dingin umumnya dilakukan pada bulan Januari dan Februari, dan survei musim semi pada bulan Maret dan April. Pelayaran CalCOFI dilakukan di sepanjang enam garis transek utama yang terletak di lepas pantai California Selatan, dengan garis yang bertambah panjang dari utara ke selatan (470–700 km; Gambar 1 ). Ada lima garis transek tambahan yang terletak di lepas pantai California tengah yang disurvei lebih jarang (Gambar 1 , Tabel 2 ). Data penampakan cetacea yang dikumpulkan pada semua garis disertakan dalam studi ini untuk menangkap rentang tipe habitat seluas mungkin dalam model. Baik survei musim panas dan gugur SWFSC maupun survei musim dingin dan semi CalCOFI dilakukan dengan menggunakan metode garis-transek (Buckland et al. 2001 ), meskipun protokolnya agak bervariasi seperti yang dirangkum di bawah ini.
| Survei | Tanggal | Upaya (km) | Daerah | Sumber |
|---|---|---|---|---|
| Kamera91 | Juli–November 1991 | 10.353,60 | Kalifornia | Hill dan Barlow ( 1992 ) |
| POD93 | Juli–November 1993 | 6437.40 | Kalifornia/Baja | Mangels dan Gerrodette ( 1994 ) |
| ORCAWAL96 | Juli–November 1996 | 15.530,80 | WA, ATAU, CA | von Saunder dan Barlow ( 1999 ) |
| ORCAWAL01 | Juli–Desember 2001 | 10.343,40 | WA, ATAU, CA | Appler dan kawan-kawan ( 2004 ) |
| CSCAP05 | Juli–Desember 2005 | 10.221,90 | WA, ATAU, CA | Forney ( 2007 ) |
| ORCAWAL08 | Juli–November 2008 | 12.242,30 | WA, ATAU, CA | Barlow dan kawan-kawan ( 2010 ) |
| DELPHINUS09 | September–Desember 2009 | 4389.10 | CenCA, SoCA, Baja | Chivers dan kawan-kawan ( 2010 ) |
| KalkulusCEAS14 | Agustus–Desember 2014 | 10.205,40 | WA, ATAU, CA | Barlow ( 2016 ) |
| CCES18 | Juni–Desember 2018 | 9554.70 | Kanada. WA, OR, CA, Baja | Henry dan kawan-kawan ( 2020 ) |
| Total | 89.278,60 |
Catatan: Wilayah yang tercakup dalam area studi: WA, Washington; OR, Oregon; CA, California; CenCA, California Tengah; SoCA, California Selatan; Baja, Baja California, Meksiko.
| Tahun | Tanggal | Wilayah |
|---|---|---|
| tahun 2005 | 4–19 Januari | Selatan |
| 15–30 April | ||
| tahun 2006 | 4–25 Februari | Penuh |
| 1–17 April | ||
| Tahun 2007 | 12 Januari−2 Februari | Penuh |
| 28 Maret−16 April | ||
| Tahun 2008 | 8–24 Januari | Selatan |
| 25 Maret−6 April | ||
| Tahun 2009 | 8–22 Januari | Selatan |
| 7–21 Maret | ||
| Tahun 2010 | 13 Januari−4 Februari | Penuh |
| Tahun 2011 | 12 Januari−6 Februari | Penuh |
| Tahun 2012 | 27 Januari−3 Februari | Selatan |
| 26 Maret−3 April | ||
| Tahun 2013 | 11–31 Januari | Penuh |
| 6–29 April | ||
| Tahun 2014 | 29 Januari−4 Februari | Selatan |
| 28 Maret−17 April | ||
| Tahun 2015 | 15 Januari−7 Februari | Penuh |
| 4–19 April | ||
| Tahun 2016 | 7–24 Januari | Penuh |
| 1–21 April | ||
| Tahun 2017 | 5–19 Januari | Penuh |
| 28 Maret−19 April | ||
| Tahun 2018 | 1–9 Februari | Penuh |
| 5–26 April | ||
| Tahun 2019 | 6–12 Februari | Selatan |
| 2–17 April | ||
| Tahun 2020 | 4–21 Januari | Selatan |
Catatan: Entri di bawah “Wilayah” menunjukkan apakah ada upaya hanya pada enam garis transek selatan CalCOFI utama (“Selatan”) atau pada semua 11 garis transek, termasuk lima garis transek utara CalCOFI (“Penuh”; lihat Gambar 1 ).
2.1.1 Survei Musim Panas dan Musim Gugur SWFSC
Setiap survei menggunakan kapal penelitian NOAA dan tim yang terdiri dari enam pengamat visual yang berpengalaman. Untuk setiap rotasi, tiga pengamat yang ditempatkan di anjungan terbang kapal secara visual mencari dan mencatat penampakan cetacea antara 0° dan 90° ke kiri dan kanan menggunakan protokol garis transek standar. Ketinggian anjungan terbang bervariasi antara keempat kapal yang digunakan pada survei ini, tetapi ini ditemukan sebagai faktor yang paling tidak penting yang mempengaruhi jarak penampakan tegak lurus pada survei garis transek kapal SWFSC (Barlow et al. 2001 ). Pengamat di kiri dan kanan mencari dengan teropong 25 × 150 yang dipasang pada alas, dan pengamat ketiga yang ditempatkan di tengah mencari dengan mata atau dengan teropong genggam 7 × 50. Penampakan dicatat bersama dengan jarak dan arah dari kapal, dari mana jarak penampakan tegak lurus dihitung. Untuk penampakan yang berada dalam jarak tiga mil laut (5,6 km) tegak lurus dari transek, kapal biasanya akan mengalihkan “upaya off-effort” untuk mendekati hewan untuk identifikasi spesies dan estimasi ukuran kelompok. Semua pengamat secara independen memberikan estimasi ukuran kelompok terbaik, tinggi, dan rendah dan persentase setiap spesies yang diidentifikasi dalam kelompok. Untuk setiap pengamat, estimasi terbaik dikalikan dengan persentase estimasi setiap spesies, dan angka yang dihasilkan dirata-ratakan di antara semua pengamat (yaitu, rata-rata aritmatika) untuk memperoleh estimasi ukuran kelompok tunggal untuk setiap penampakan. Protokol survei sama untuk semua tahun dan dijelaskan secara lebih rinci di tempat lain (misalnya, Barlow dan Forney 2007 ; Kinzey et al. 2000 ).
2.1.2 Survei Musim Dingin dan Musim Semi CalCOFI
Enam kapal penelitian yang berbeda digunakan selama periode survei 2005–2020, tetapi Douglas et al. ( 2014 ) tidak menemukan perbedaan signifikan dalam jarak pandang tegak lurus untuk kapal dengan ketinggian platform yang bervariasi. Semua survei dilakukan dalam mode lewat (yaitu, kapal tidak dialihkan untuk identifikasi spesies atau penghitungan ukuran kelompok setelah penampakan) dengan dua pengamat khusus mencari cetacea menggunakan mata telanjang dan teropong genggam 7 × 50. Penampakan, estimasi ukuran kelompok, upaya, dan data cuaca dicatat secara sistematis dan dimasukkan ke dalam rekaman elektronik. Kami hanya menyertakan penampakan yang dilakukan saat dua pengamat berusaha pada garis CalCOFI standar, tidak termasuk transit. Deskripsi terperinci dari protokol survei dapat ditemukan di Douglas et al. ( 2014 ) dan Campbell et al. ( 2015 ).
2.2 Pengolahan Data dan Variabel Habitat
Untuk membuat sampel untuk SWFSC musim panas dan gugur dan SDM musim dingin dan semi CalCOFI, bagian kontinu dari jalur survei on-effort dibagi menjadi segmen sekitar 5 km menggunakan metode yang dijelaskan oleh Becker et al. Panjang segmen 5 km dipilih agar cukup halus untuk menangkap perubahan skala lebih halus dalam kondisi habitat di area studi CCE (misalnya, batimetri), dan cukup kasar untuk mengurangi jumlah segmen tanpa penampakan (Barlow et al. 2009 ). Hanya data on-effort yang dikumpulkan dalam kondisi keadaan laut Beaufort ≤ 5 dalam area studi CCE yang digunakan dalam pengembangan model. Jumlah total penampakan spesifik spesies dan estimasi ukuran kelompok rata-rata terkait ditetapkan ke setiap segmen, dan kovariat habitat diturunkan berdasarkan titik tengah geografis segmen.
Variabel lingkungan dari implementasi CCE asimilasi data dari Regional Ocean Modeling System (ROMS), yang diproduksi oleh kelompok University of California Santa Cruz Ocean Modeling and Data Assimilation (Moore et al. 2011 ), digunakan sebagai prediktor dinamis, karena telah terbukti efektif dalam SDM untuk area studi ini (Abrahms et al. 2019 ; Becker et al. 2016 , 2017 , 2018 , 2020 ; Hazen et al. 2018 ; Lezama-Ochoa et al. 2024 ). Rata-rata harian untuk setiap variabel pada resolusi piksel sekitar 10 km × 10 km (yaitu, elemen gambar) dari keluaran ROMS digunakan dalam model. Rangkaian prediktor dinamis potensial mencakup suhu permukaan laut (SST) dan deviasi standarnya (SSTsd; dihitung untuk kotak 3 × 3 piksel di sekitar titik tengah segmen pemodelan), kedalaman lapisan campuran (MLD, didefinisikan oleh deviasi 0,5 °C dari SST), tinggi permukaan laut (SSH), dan deviasi standar tinggi permukaan laut (SSHsd; juga dihitung untuk kotak 3 × 3 piksel di sekitar titik tengah segmen pemodelan). Kedalaman air juga disertakan sebagai prediktor potensial, yang diturunkan dari model relief global 1-busur-menit ETOPO1 (Amante dan Eakins 2009 ). Semua nilai kovariat diperoleh untuk titik tengah setiap segmen transek.
Tidak seperti SDM sebelumnya yang dikembangkan menggunakan data survei CCE SWFSC yang sama (misalnya, Becker et al. 2020 ), tidak ada model yang ditawarkan kovariat spasial (lintang, bujur) atau temporal (tahun) karena istilah-istilah ini dapat membatasi kemampuan peramalan model (Becker et al. 2018 ). Misalnya, jika lintang disertakan dalam model musim dingin dan semi CalCOFI, itu akan menghalangi pembuatan prediksi di luar domain spasial asli tersebut ke area studi CCE yang lebih luas. Lebih jauh, tahun dapat bertindak sebagai proksi untuk variabel dinamis, yang dalam kasus ini dapat mengacaukan ekstrapolasi spasial dan musiman.
2.3 Metode Pemodelan
2.3.1 Pendekatan Umum
Bahasa Indonesia: Untuk menyediakan ekstrapolasi temporal, kami mengembangkan model kepadatan cetacea berbasis habitat menggunakan data penampakan yang dikumpulkan dari survei SWFSC yang dilakukan di seluruh area studi CCE pada musim panas dan gugur antara tahun 1991 dan 2018 (Tabel 1 ). Mengingat bahwa survei CalCOFI dilakukan tiap triwulan, kami awalnya menyelidiki potensi untuk menggabungkan data musim panas dan gugur SWFSC dan CalCOFI menjadi satu model tunggal, menggunakan data penampakan musim panas dan gugur untuk mengevaluasi kinerja. Kami menemukan bahwa pola distribusi musim panas dan gugur yang diprediksi dari hasil model gabungan tidak seakurat model yang dibangun hanya dengan data SWFSC. Karena data survei CalCOFI memberikan kontribusi yang relatif kecil terhadap ukuran sampel yang tersedia dari data SWFSC, dan daya prediksi tidak meningkat menggunakan model gabungan, kami mengembangkan model musim panas dan gugur hanya menggunakan data SWFSC. Untuk ekstrapolasi spasial, kami membangun model dari 29 survei CalCOFI di atas kapal yang dilakukan selama musim dingin dan semi antara tahun 2005 dan 2020 (Tabel 2 ).
Dengan menggunakan kerangka kerja Generalized Additive Modeling (GAM; Wood 2017 ) yang mapan berdasarkan pengambilan sampel jarak jauh (Buckland et al. 2001 ), model-model disesuaikan untuk empat spesies dengan ukuran sampel yang cukup tersedia di kedua set data untuk pemodelan (Tabel 3 ): lumba-lumba biasa berparuh pendek, lumba-lumba sisi putih Pasifik, lumba-lumba Dall, dan paus sirip. Variabel habitat mencakup prediktor statis dan dinamis yang terbukti penting dalam SDM sebelumnya (misalnya, Becker et al. 2014 , 2016 ; Hazen et al. 2017 ). Dengan demikian, kami menghasilkan dua set model untuk setiap spesies: (1) model musim panas dan gugur yang mencakup seluruh area studi CCE, dan (2) model musim dingin dan semi yang hanya mencakup area studi CalCOFI yang lebih terbatas di lepas pantai California tengah/selatan (Gambar 1 ). Baik model musim panas dan gugur SWFSC maupun model musim dingin dan semi CalCOFI kemudian digunakan untuk memprediksi kondisi habitat dari Desember hingga Februari (“musim dingin”) dan Maret hingga Mei (“musim semi”) untuk perairan di seluruh area studi CCE dari 2017 hingga 2021 menggunakan teknik yang dirancang untuk mengurangi atau menghindari artefak ekstrapolasi spasial dan temporal (Bouchet et al. 2019 ). Model yang dihasilkan memberikan perbandingan ekstrapolasi temporal (musiman) dari model musim panas dan gugur SWFSC dan ekstrapolasi spasial dari model musim dingin dan semi CalCOFI. Pola distribusi spasial dan estimasi kelimpahan area studi yang berasal dari dua set prediksi model dievaluasi berdasarkan apa yang saat ini diketahui tentang kemunculan spesies di lepas Pantai Barat AS pada musim dingin.
| SWFSC Musim Panas/Musim Gugur | CalCOFI Musim Dingin/Musim Semi | ||||
|---|---|---|---|---|---|
| Nama umum | Nama taksonomi | # Pemandangan | Rata-rata GS | # Pemandangan | Rata-rata GS |
| Lumba-lumba biasa berparuh pendek | Delphinus delphis delphis | tahun 1034 | 155.73 | 256 | 81.73 |
| Lumba-lumba sisi putih Pasifik | Lagenorhynchus miring | 296 | 54.7 | 48 | 34.72 |
| Lumba-lumba Dall | Phocoenoides dalli | 678 | 3.72 | 88 | 6.35 |
| Paus sirip | Burung Balaenoptera | 558 | 2.06 | 39 | 1.64 |
Catatan: Semua penampakan dilakukan saat melakukan upaya sistematis di kondisi laut Beaufort ≤ 5 dalam jarak pemotongan spesifik spesies (lihat teks untuk detailnya).
2.3.2 Parameter Deteksi
Dua parameter deteksi diperlukan untuk memperkirakan kepadatan dari survei garis-transek: (1) lebar strip efektif ( ESW ), yang menyediakan ukuran jarak dari garis lintasan tempat spesies terlihat berdasarkan fungsi deteksi probabilitas, dan (2) g (0), probabilitas deteksi langsung pada garis transek. Mengikuti metode Becker et al. ( 2016 ), estimasi spesifik spesies dan segmen dari ESW dan g (0) dimasukkan ke dalam model musim panas dan gugur berdasarkan kondisi deteksi yang terekam pada segmen tersebut dan menggunakan koefisien yang diestimasi khusus untuk dataset CCE berdasarkan metode Barlow et al. ( 2011 ) untuk ESW dan Barlow ( 2015 ) untuk g (0). Untuk segmen-segmen di mana keadaan laut Beaufort rata-rata adalah 0 (< 1% dari segmen), g (0) diasumsikan 1, yaitu, semua hewan langsung pada garis transek terdeteksi.
Hanya data dari musim dingin dan musim semi yang digunakan untuk menyesuaikan model musim dingin dan musim semi; namun, fungsi deteksi disesuaikan dengan data penampakan CalCOFI 2005–2020 (sepanjang tahun) secara penuh dengan mengikuti metode Becker et al. ( 2017 ) untuk meningkatkan ukuran sampel guna menyesuaikan fungsi deteksi. Data CalCOFI musim panas dan musim gugur memberikan tambahan 196 penampakan lumba-lumba biasa berparuh pendek, 28 penampakan lumba-lumba sisi putih Pasifik, 7 penampakan lumba-lumba Dall, dan 180 penampakan paus sirip. Baik fungsi kunci setengah normal maupun fungsi kunci tingkat bahaya tanpa istilah penyesuaian disesuaikan, dan kriteria informasi Akaike (AIC; Akaike 1973 ) dan inspeksi visual plot deteksi (Thomas et al. 2010 ) digunakan untuk memilih model terbaik. Mengingat pengaruh kondisi laut Beaufort pada kemampuan deteksi (Barlow 2015 ; Barlow et al. 2001 , 2011 ), terdapat potensi heterogenitas dalam fungsi deteksi untuk membiaskan hasil (Rexstad et al. 2023 ); oleh karena itu, fungsi deteksi dihasilkan dengan kondisi laut Beaufort sebagai kovariat (Buckland et al. 2001 ; Marques et al. 2007 ) menggunakan paket R mrds (v. 2.2.3) dan Distance (v. 1.0.2). Estimasi ESW spesifik spesies dan segmen kemudian dimasukkan ke dalam model berdasarkan kondisi kondisi laut yang terekam pada segmen tersebut.
Estimasi spesifik segmen g (0) yang diturunkan secara khusus untuk dataset SWFSC CCE dan berdasarkan metode Barlow ( 2015 ) juga diterapkan pada dataset CalCOFI musim dingin dan musim semi. Estimasi g (0) ini diharapkan menjadi koreksi minimum (yaitu, kemungkinan meremehkan kepadatan) untuk tim pengamat dua orang selama survei CalCOFI, karena estimasi berdasarkan Barlow ( 2015 ) adalah untuk tim pengamat 3 orang termasuk dua pengamat yang mencari dengan teropong 25 × 150 yang dipasang pada alas.
2.3.3 Faktor Koreksi Tambahan
Bahasa Indonesia: Melaksanakan survei dalam mode lewat membatasi kemampuan pengamat untuk mengidentifikasi spesies secara positif, yang menghasilkan sejumlah besar paus besar tak teridentifikasi dan lumba-lumba biasa tak teridentifikasi (baik D. delphis delphis atau D. delphis bairdii ). Semua survei musim dingin dan semi CalCOFI dilaksanakan dalam mode lewat, seperti juga sebagian upaya pada survei SWFSC musim panas dan gugur 2018. Menghilangkan penampakan “paus besar tak teridentifikasi” dan “lumba-lumba biasa tak teridentifikasi” ini dari kumpulan data pemodelan akan menghasilkan estimasi kepadatan hewan yang terlalu rendah untuk paus sirip dan lumba-lumba biasa berparuh pendek. Untuk mengurangi potensi bias ke bawah ini, faktor koreksi khusus spesies diterapkan ke semua segmen CalCOFI dan ke segmen mode lewat survei SWFSC 2018 untuk memperhitungkan hewan tak teridentifikasi, menggunakan metode yang dijelaskan dalam Becker et al. ( 2017 ) dan Becker et al. ( 2020 ).
Baik untuk kelompok paus besar maupun lumba-lumba biasa, faktor koreksi c diperkirakan dari data penampakan menurut rumus yang disederhanakan:
![]()
di mana t tgt adalah jumlah individu yang diidentifikasi sebagai spesies target, t oth adalah jumlah individu yang diidentifikasi sebagai spesies lain dalam kelompok spesies yang lebih luas, dan t unid adalah jumlah individu yang tidak teridentifikasi dalam kelompok spesies tersebut. Karena potensi efek keadaan laut Beaufort pada detektabilitas (Barlow 2015 ; Barlow et al. 2001 , 2011 ), faktor koreksi dievaluasi untuk menentukan apakah mereka bervariasi menurut keadaan laut. Jika demikian, faktor koreksi terpisah dikembangkan menurut keadaan laut; jika tidak, satu faktor koreksi diterapkan. Faktor koreksi diterapkan pada jumlah hewan yang diperkirakan per segmen dalam SDM untuk lumba-lumba biasa berparuh pendek dan paus sirip (lihat Persamaan 2 di bawah).
2.3.4 Model Habitat
Metode pemodelan sebagian besar mengikuti yang dijelaskan dalam Becker et al. ( 2020 ) untuk model musim panas dan gugur serta Becker et al. ( 2017 ) untuk model musim dingin dan semi. Untuk kedua set model, GAM dikembangkan dalam R (v. 4.0.2; R Core Team 2022 ) menggunakan paket “ mgcv ” (v. 1.8–31; Wood 2011 ). Kemungkinan maksimum terbatas (REML) digunakan untuk memperoleh estimasi parameter (Marra dan Wood 2011 ). Pendekatan penyusutan Marra dan Wood ( 2011 ) digunakan untuk berpotensi menghilangkan istilah dari setiap model dengan memodifikasi penalti penghalusan, yang memungkinkan efek penghalusan menyusut menjadi nol. Selain itu, untuk menghindari overfitting, proses seleksi maju/mundur berulang digunakan untuk menghilangkan variabel yang memiliki nilai p > 0,05 (Redfern et al. 2017 ; Roberts et al. 2016 ).
Dua kerangka kerja pemodelan khusus spesies yang berbeda digunakan, tergantung pada karakteristik ukuran kelompok. Untuk semua spesies kecuali lumba-lumba biasa berparuh pendek, GAM disesuaikan menggunakan jumlah individu spesies tertentu per segmen transek sebagai variabel respons menggunakan semua segmen transek dan distribusi Tweedie untuk memperhitungkan penyebaran berlebih. Untuk lumba-lumba biasa berparuh pendek, spesies yang memiliki ukuran kelompok yang sangat besar dan bervariasi (misalnya, 1 hingga 2000 hewan per penampakan), model tingkat perjumpaan kelompok yang terpisah dikembangkan. Model tingkat perjumpaan kelompok dibangun menggunakan semua segmen transek, terlepas dari apakah mereka menyertakan penampakan, menggunakan kelompok yang terlihat per segmen sebagai variabel respons dan distribusi Tweedie untuk memperhitungkan penyebaran berlebih (Miller et al. 2013 ). Ukuran kelompok yang diharapkan berdasarkan metode regresi bias ukuran (Buckland et al. 2001 ; Thomas et al. 2010 ) kemudian digunakan dalam persamaan kepadatan (lihat Persamaan 2 di bawah). Rangkaian lengkap prediktor habitat potensial ditawarkan kepada tingkat perjumpaan kelompok dan GAM respons tunggal.
Prediksi dari model akhir dimasukkan ke dalam persamaan garis-transek standar (Buckland et al. 2001 ) untuk memperkirakan kepadatan ( D ; jumlah hewan per km 2 ):
![]()
di mana i adalah segmen, n adalah jumlah penampakan pada segmen i , s adalah ukuran kelompok rata-rata (yaitu, jumlah spesies tertentu yang hadir dalam suatu kelompok) pada segmen i , c adalah faktor koreksi khusus spesies untuk lumba-lumba umum yang tidak teridentifikasi atau paus besar (diturunkan dalam Persamaan 1 dan diasumsikan 1 untuk semua spesies lainnya) berdasarkan kondisi keadaan laut pada segmen i , dan A adalah area efektif yang dicari untuk segmen i :
![]()
di mana L i adalah panjang segmen upaya i , ESW i adalah setengah lebar strip efektif, dan g (0) i adalah probabilitas deteksi pada garis transek.
Performa model dievaluasi menggunakan metrik yang ditetapkan, termasuk persentase deviasi yang dijelaskan, area di bawah kurva karakteristik operasi penerima (AUC; Fawcett 2006 ), statistik keterampilan sebenarnya (TSS; Allouche et al. 2006 ), dan inspeksi visual dari distribusi yang diprediksi dan diamati selama survei cetacea musim panas dan gugur 1991–2018 dan musim dingin dan semi 2005–2020 (Barlow et al. 2009 ; Becker et al. 2016 , 2017 ; Forney et al. 2012 ). AUC mengukur akurasi prediksi kehadiran dan ketidakhadiran yang diamati; nilai berkisar dari 0 hingga 1, di mana skor > 0,5 menunjukkan keterampilan yang lebih baik daripada keterampilan acak. TSS memperhitungkan kesalahan negatif palsu dan kesalahan positif palsu dan berkisar dari −1 hingga +1, di mana +1 menunjukkan kesepakatan sempurna dan nilai nol atau kurang menunjukkan kinerja yang tidak lebih baik daripada acak. Untuk menghitung TSS, pendekatan maksimalisasi jumlah sensitivitas-spesifisitas (Liu et al. 2005 ) digunakan untuk menentukan ambang batas untuk mengubah kepadatan spesies menjadi ada/tidaknya.
Selain itu, estimasi kelimpahan berbasis model untuk area studi musim panas dan gugur serta musim dingin dan semi masing-masing berdasarkan jumlah prediksi segmen pemodelan individual dibandingkan dengan estimasi garis transek standar yang diperoleh dari kumpulan data yang sama yang digunakan untuk pemodelan guna menilai potensi bias dalam prediksi model berbasis habitat (Becker et al. 2022 ). Estimasi garis transek standar diperoleh dari data survei menggunakan Persamaan ( 2 ) dan ( 3 ) di atas, tetapi tanpa penyertaan prediktor habitat (yaitu, kepadatan yang diamati daripada yang diprediksi).
Untuk setiap spesies, model akhir digunakan untuk memprediksi kepadatan di setiap sel grid 10 × 10 km di seluruh area studi CCE untuk rata-rata harian yang berbeda dari kondisi lingkungan di musim dingin (Desember-Februari) dan musim semi (Maret-Mei) selama 5 tahun terakhir (2017-2021). Sementara data survei tidak mencakup periode waktu yang tepat ini, kami memilih 5 tahun terakhir untuk prediksi karena mereka paling relevan untuk tujuan manajemen dan pekerjaan sebelumnya telah menunjukkan bahwa model mampu menangkap variabilitas antartahunan dalam area studi (Becker et al. 2018 ). Sebelum membuat prediksi, alat deteksi ekstrapolasi dalam paket R ” dsmextra ” (Bouchet et al. 2020 ) digunakan untuk menilai sumber dan tingkat ekstrapolasi dalam prediksi musim dingin dan musim semi untuk kedua set model. Piksel dengan kovariat habitat di luar ruang data yang digunakan untuk membangun model masing-masing kemudian dihilangkan dari setiap permukaan harian, karena ini merupakan metode yang sederhana dan efektif untuk menghindari ekstrapolasi (Bouchet et al. 2019 ).
Prediksi grid resolusi 10 km harian yang terpisah dirata-ratakan untuk menghasilkan grid yang eksplisit secara spasial dari kepadatan spesies musim dingin dan musim semi rata-rata dan ukuran ketidakpastian. Untuk masing-masing dari empat spesies, ada dua prediksi terpisah untuk setiap periode musiman berdasarkan prediksi dari model musim panas dan musim gugur SWFSC (selanjutnya disebut “model yang diekstrapolasi secara musiman”) dan model musim dingin dan musim semi CalCOFI (selanjutnya disebut “model yang diekstrapolasi secara spasial”).
Sebagai langkah validasi awal, kami menggunakan data dari empat survei CalCOFI musim dingin dan musim semi terkini (2017–2020; data tidak tersedia untuk tahun 2021) untuk mengevaluasi ekstrapolasi musiman model musim panas dan musim gugur SWFSC dalam area studi CalCOFI. Untuk setiap spesies, model yang diekstrapolasi secara musiman dan spasial digunakan untuk memperkirakan kelimpahan khusus untuk area studi CalCOFI, dan rasio estimasi tahunan dihitung. Selain itu, penampakan dari survei CalCOFI 2017–2020 ditumpangkan pada permukaan kepadatan yang diekstrapolasi secara musiman untuk membandingkan distribusi yang diprediksi dan diamati untuk setiap tahun.
2.3.5 Ketidakpastian Model
Variasi dalam kondisi lingkungan telah menjadi salah satu sumber ketidakpastian terbesar saat memprediksi kepadatan sebagai fungsi variabel habitat dalam ekosistem yang sangat dinamis seperti California Current, dan variasi ini telah digunakan dalam publikasi sebelumnya untuk menyediakan ukuran varians eksplisit spasial untuk prediksi model musim panas dan gugur SWFSC dan musim dingin dan semi CalCOFI (Barlow et al. 2009 ; Becker et al. 2016 , 2017 , 2020 ; Forney et al. 2012 ). Miller et al. ( 2022 ) baru-baru ini mengembangkan teknik untuk mendapatkan ukuran ketidakpastian yang lebih komprehensif dalam prediksi GAM yang memperhitungkan ketidakpastian gabungan dari variabilitas lingkungan, koefisien GAM, ESW , dan g (0). Teknik-teknik ini termasuk menghasilkan beberapa permukaan kepadatan harian (untuk raster kovariat pada setiap irisan waktu) yang memperhitungkan ketidakpastian parameter model (melalui pengambilan sampel posterior dari parameter model) dan menyediakan berbagai kemungkinan estimasi kepadatan yang darinya varians dapat dihitung. Untuk studi ini, Miller et al. ( 2022 ) metode diterapkan untuk memperkirakan ukuran varians eksplisit spasial yang memperhitungkan sumber ketidakpastian gabungan ini.
Estimasi g (0) yang dikembangkan oleh Barlow ( 2015 ) dan digunakan untuk pengembangan model didasarkan pada kondisi keadaan laut Beaufort spesifik segmen, yang saat ini tidak kompatibel dengan metode Miller et al. ( 2022 ) untuk memperkirakan varians. Oleh karena itu, untuk menggabungkan varians g (0) ke dalam estimasi spesifik piksel, estimasi keseluruhan ketidakpastian dalam g (0) diturunkan menggunakan estimasi varians untuk parameter ini yang dibobot dengan proporsi upaya survei yang dilakukan dalam setiap kategori keadaan laut Beaufort untuk masing-masing survei musim panas dan gugur SWFSC dan survei musim dingin dan semi CalCOFI, dan diestimasi berdasarkan 10.000 nilai bootstrap. Estimasi g ( 0) Barlow ( 2015 ) untuk lumba-lumba sisi putih Pasifik dianggap sebagai outlier (yaitu, 1 untuk semua keadaan laut), mungkin karena ukuran sampel yang kecil, jadi untuk spesies ini estimasi g (0) untuk lumba-lumba umum digunakan karena mereka memiliki karakteristik penampakan yang serupa (Jefferson et al. 2015 ).
Untuk lumba-lumba biasa berparuh pendek, estimasi ketidakpastian berbasis piksel didasarkan pada model tingkat pertemuan kelompok, yang tidak menyertakan ketidakpastian dalam estimasi ukuran kelompok karena ukuran kelompok yang diharapkan dimasukkan ke dalam persamaan kepadatan (Persamaan 2 ) sebagai konstanta berdasarkan metode regresi bias ukuran. Oleh karena itu, estimasi varians berbasis piksel untuk spesies ini sedikit diremehkan. Namun, analisis kami mencakup sumber ketidakpastian yang dominan pada tingkat yang lebih besar daripada penelitian serupa sebelumnya (misalnya, Becker et al. 2017 , 2020 ).
2.3.6 Perbandingan Model
Perbedaan dalam dua set distribusi spasial yang diprediksi model dan estimasi kelimpahan spesies di area studi dievaluasi relatif terhadap pengetahuan terkini tentang kemunculan spesies di lepas Pantai Barat AS pada musim dingin. Ini termasuk analisis data survei sistematis yang dikumpulkan di berbagai wilayah area studi CCE antara tahun 1975 dan 1992, termasuk survei kapal dan udara yang dilakukan di Southern California Bight (Dohl et al. 1978 ), survei udara yang dilakukan di perairan dekat pantai di lepas pantai California (Dohl et al. 1983 ; Forney dan Barlow 1998 ), dan survei kapal dan udara yang dilakukan di lepas pantai Oregon dan Washington (Green et al. 1992 ). Data yang lebih baru digunakan untuk mengevaluasi kinerja model mencakup prediksi kepadatan cetacea relatif di lepas pantai Washington berdasarkan beberapa sumber data (Menza et al. 2016 ), prediksi kesesuaian habitat untuk paus sirip di California Current berdasarkan data tag (Scales et al. 2017 ), dan model distribusi spesies untuk rorqual di lepas pantai Oregon (Derville et al. 2022 ). Selain itu, estimasi kepadatan cetacea berbasis desain dan model sebelumnya berdasarkan survei kapal CalCOFI 2005 hingga 2015 di lepas pantai California selatan dan tengah (Becker et al. 2017 ; Campbell et al. 2015 ) juga digunakan untuk tujuan validasi silang.
Untuk setiap spesies, estimasi kelimpahan berbasis model musim dingin dan musim semi untuk area studi CCE dihitung sebagai jumlah estimasi kelimpahan sel grid individual, yang diperoleh dengan mengalikan area sel (dalam km 2 ) dengan kepadatan sel grid yang diprediksi, eksklusif dari setiap bagian sel yang terletak di luar area studi atau di daratan. Perhitungan area diselesaikan menggunakan paket R geosphere dan gpclib dalam R (versi 2.15.0). Ketidakpastian untuk estimasi kelimpahan area studi CCE diestimasi menggunakan teknik Miller et al. ( 2022 ) yang dijelaskan di atas dan dengan demikian memperhitungkan ketidakpastian gabungan dari variabilitas lingkungan, koefisien GAM, ESW , dan g (0). Untuk lumba-lumba biasa berparuh pendek, ketidakpastian ukuran kelompok dari estimasi regresi bias ukuran digabungkan ke dalam estimasi varians area studi menggunakan metode delta (Seber 1982 ).
3 Hasil
Sebanyak 89.279 km data survei on-effort yang dikumpulkan pada 9 survei SWFSC dari Juli hingga November antara 1991 dan 2018 dalam area studi CCE digunakan untuk mengembangkan model musim panas dan gugur. Jumlah penampakan dalam jarak pemotongan spesifik spesies dan tersedia untuk pemodelan berkisar antara 296 hingga 1034 (Tabel 3 ). Sebanyak 33.995 km data on-effort dari 29 survei CalCOFI yang dilakukan selama Januari hingga April 2005–2020 digunakan untuk mengembangkan model musim dingin dan semi. Jumlah penampakan yang digunakan untuk pemodelan jauh lebih rendah daripada yang tersedia dari survei musim panas dan gugur, berkisar antara 39 hingga 256 (Tabel 3 ).
Ada enam spesies tambahan yang penampakannya ada di kedua set survei tetapi tidak cukup penampakannya di dataset musim dingin dan semi CalCOFI untuk mendukung pengembangan model yang diekstrapolasi secara spasial, termasuk lumba-lumba belang ( Stenella coeruleoalba ), lumba-lumba biasa berparuh panjang ( Delphinus delphis bairdii ), lumba-lumba hidung botol biasa ( Tursiops truncatus ), lumba-lumba Risso ( Grampus griseus ), lumba-lumba paus sikat utara ( Lissodelphis borealis ), dan paus minke ( Balaenoptera acutorostrata ). Model yang diekstrapolasi secara musiman dikembangkan untuk spesies ini dari model musim panas dan gugur SWFSC. Untuk kelengkapan, hasil model untuk spesies ini disajikan dalam Informasi Pendukung S.3 .
Rentang nilai kovariat yang disertakan dalam dataset pemodelan musim panas dan gugur SWFSC secara umum lebih besar daripada dataset musim dingin dan semi CalCOFI, yang tidak mengejutkan mengingat rentang lintang yang jauh lebih besar yang disertakan dalam area studi CCE SWFSC penuh (Tabel 4 ). Namun, tingkat ekstrapolasi dalam permukaan prediksi harian musim dingin dan semi secara umum lebih besar untuk dataset pemodelan musim panas dan gugur SWFSC, meskipun hanya ekstrapolasi univariat (vs. ekstrapolasi kombinatorial) yang terbukti untuk kedua dataset pemodelan (Informasi Pendukung S.1 ). Untuk menghindari artefak ekstrapolasi, piksel dengan kovariat di luar rentang dataset pemodelan masing-masing dibuang dari permukaan kepadatan harian sebelum membuat prediksi dari model. Persentase piksel yang dihilangkan sepanjang tahun berkisar dari 3,4% (model musim dingin dan musim semi CalCOFI lumba-lumba biasa berparuh pendek) hingga 14,7% (model SWFSC musim panas dan musim gugur paus sirip) dari total 10.942.021 nilai piksel potensial sepanjang tahun (12.011 piksel per permukaan kepadatan dan 911 permukaan harian).
| Variabel | SWFSC musim panas dan gugur | CalCOFI musim dingin dan musim semi | Permukaan prediksi | |
|---|---|---|---|---|
| Suhu permukaan laut (°C) | Berarti | 16.76 | 14.68 | 12.81 |
| menit | 9.96 | 9.17 | 6.39 | |
| Maksimal | Tanggal 23.18 | 18.92 | 22.41 | |
| SSH (cm) | Berarti | 0.13 | 0.14 | 0.13 |
| menit | -0,09 | -0,07 | -0,02 | |
| Maksimal | 0.4 | 0.36 | 0,59 | |
| MLD (m) | Berarti | 22.46 | 46.91 | 46.1 |
| menit | 0.21 | 0.42 | 0.18 | |
| Maksimal | 102.67 | 170.8 | 315.11 | |
| SSTsd | Berarti | 0.21 | 0.18 | 0.14 |
| menit | 0,006 | 0,01 | 0,0005 | |
| Maksimal | 2.16 | 1.69 | 4.22 | |
| SSH | Berarti | 0,01 | 0,009 | 0,01 |
| menit | 0,0003 | 0,0003 | 0,0000007 | |
| Maksimal | 0,05 | 0,04 | 0.11 | |
| Kedalaman (m) | Berarti | -2914 | -2783 | -3187,97 |
| menit | -5036 | -4787 | -5060 | |
| Maksimal | -15 | -13 | angka 0 |
Catatan: Nilai mencakup nilai rata-rata, minimum (min), dan maksimum (maks) yang diamati selama survei masing-masing berdasarkan nilai Regional Ocean Modeling System (ROMS) untuk semua segmen pemodelan dari survei masing-masing. Juga ditunjukkan nilai kovariat untuk semua permukaan harian berbasis ROMS yang digunakan untuk membuat prediksi musim dingin dan musim semi area studi Ekosistem Arus California secara menyeluruh (kovariat habitat di luar ruang data yang digunakan untuk membangun model masing-masing dihilangkan sebelum membuat prediksi). Kovariat mencakup suhu permukaan laut (SST), tinggi permukaan laut (SSH), kedalaman lapisan campuran (MLD), kedalaman batimetri (kedalaman), dan deviasi standar (sd) dari SST dan SSH (lihat teks untuk detailnya). 3.1 Fungsi Deteksi Untuk data musim dingin dan semi CalCOFI, model setengah normal dengan keadaan laut Beaufort sebagai kovariat memberikan kecocokan terbaik dengan data jarak tegak lurus untuk keempat spesies. Jarak pemotongan untuk lumba-lumba umum adalah 1,5 km, menghilangkan 18% penampakan terjauh. Ini lebih tinggi dari persentase yang direkomendasikan sebesar 5%–15% (Buckland et al. 2001 ), tetapi dalam mode lewat, estimasi ukuran kelompok menurun seiring bertambahnya jarak, yang menunjukkan bias estimasi pada jarak yang lebih jauh. Dengan mengecualikan persentase penampakan yang lebih besar, bias potensial dalam estimasi ukuran kelompok berkurang. Lebih lanjut, metode regresi bias ukuran digunakan untuk memperkirakan ukuran kelompok rata-rata yang diharapkan dalam jarak pemotongan ini. Jarak pemotongan untuk spesies lainnya adalah 1,2 km untuk lumba-lumba sisi putih Pasifik (menghilangkan 11% penampakan terjauh), 1,3 km untuk lumba-lumba Dall (menghilangkan 13% penampakan terjauh) dan 3,6 km untuk paus besar (menghilangkan 14% penampakan terjauh).
3.2 Faktor Koreksi
Untuk memperhitungkan hewan tak dikenal pada survei SWFSC 2018, faktor koreksi untuk paus besar tak dikenal diterapkan secara terpisah menurut kategori kondisi laut Beaufort pada penampakan paus sirip, karena proporsi paus tak dikenal meningkat seiring dengan meningkatnya kondisi laut. Faktor koreksi ini adalah 1,04, 1,08, 1,10, 1,30, dan 1,46 untuk kondisi laut 0–1, 2, 3, 4, dan 5, secara berurutan. Untuk kelompok lumba-lumba biasa, pengali yang lebih besar tidak dikaitkan dengan kondisi laut yang lebih besar, sehingga faktor koreksi seragam sebesar 1,71 diterapkan di semua kondisi laut untuk penampakan lumba-lumba biasa berparuh pendek pada survei tahun 2018.
Mirip dengan data CCE SWFSC, faktor koreksi yang diestimasikan dari data musim dingin dan musim semi CalCOFI untuk paus sirip bervariasi menurut kondisi laut, karena proporsi paus besar yang tidak teridentifikasi meningkat seiring dengan meningkatnya kondisi laut, sehingga menghasilkan faktor koreksi sebesar 1,09, 1,45, 1,54, 1,95, dan 2,12 untuk kondisi laut Beaufort 0–1, 2, 3, 4, dan 5, secara berurutan. Untuk kelompok lumba-lumba umum, pengali lebih besar di kondisi laut yang lebih rendah—mungkin karena efek pengganggu dari ukuran kelompok yang diestimasikan lebih besar di kondisi laut yang lebih rendah saat dalam mode lewat; oleh karena itu, faktor koreksi seragam sebesar 1,33 digunakan di semua kondisi laut.
3.3 Metrik Model dan Kinerja Penjelasan
Ukuran kelompok yang diharapkan untuk lumba-lumba biasa berparuh pendek berdasarkan metode regresi bias ukuran dan digunakan dalam persamaan kepadatan masing-masing (lihat Persamaan 2 ) adalah 108,16 (CV = 0,061) untuk data musim panas dan gugur SWFSC dan 47,06 (CV = 0,076) untuk data musim dingin dan semi CalCOFI.
Kinerja penjelasan serupa dengan SDM masa lalu yang dikembangkan untuk spesies ini di area studi CCE (Becker et al. 2017 , 2018 , 2020 ), dengan deviasi yang dijelaskan berkisar dari sekitar 10% hingga 39%, nilai AUC dari 0,66 hingga 0,87, nilai TSS dari 0,24 hingga 0,58, dan rasio kepadatan yang diamati: diprediksi lebih besar dari 0,79, dengan mayoritas lebih besar dari 0,93, yang menunjukkan bahwa estimasi kelimpahan absolut yang diprediksi model di masing-masing area studi serupa dengan nilai yang diperoleh dari metode transek garis berbasis desain (Tabel 5 ). Variabel prediktor utama (yaitu, yang paling berpengaruh pada model) serupa untuk kedua set model, meskipun model musim panas dan gugur SWFSC secara konsisten menyertakan lebih banyak kovariat daripada model musim dingin dan semi CalCOFI (Tabel 5 ) karena jangkauan geografis yang lebih besar dan ukuran sampel yang lebih besar dari survei SWFSC. Bentuk fungsional dari variabel prediktor secara umum serupa antara kumpulan data dalam rentang nilai kovariat yang sama, dengan pengecualian kedalaman untuk lumba-lumba biasa berparuh pendek dan SSHsd untuk lumba-lumba Dall (Gambar 2 ). Di musim dingin dan semi, lumba-lumba biasa berparuh pendek diasosiasikan dengan perairan terdalam di area studi CalCOFI, sementara di musim panas dan gugur di seluruh area studi CCE, bentuk fungsional mengindikasikan distribusi bimodal, dengan jumlah lumba-lumba terbanyak ditemukan di perairan sekitar 500 m dalam, atau lepas pantai di perairan sekitar 4000 m dalam (Gambar 2a ). Berdasarkan model musim panas dan gugur SWFSC, jumlah lumba-lumba Dall yang lebih banyak ditemukan di perairan dengan variabilitas tertinggi di SSH (yaitu, wilayah frontal), sementara di area studi CalCOFI di musim dingin dan semi, lumba-lumba Dall diasosiasikan dengan perairan dengan sedikit atau tidak ada variabilitas di SSH (Gambar 2c ).
| Jenis | Variabel prediktor | Penjelasan. Pengembangan. | AUC | TSS | Obs:Pred |
|---|---|---|---|---|---|
| Lumba-lumba biasa berparuh pendek | |||||
| Bahasa Inggris SWFSC | SST + kedalaman + SSH + SSHsd | 9.88 | 0.73 | 0.36 | 0,99 |
| Kalsium COFI | SST + kedalaman + MLD | 10.80 | 0.71 | 0,35 | 0,98 |
| Lumba-lumba sisi putih Pasifik | |||||
| Bahasa Inggris SWFSC | Kedalaman + SSH + SST + MLD + SSHsd | pukul 39.30 | 0.82 | 0.51 | 0,79 |
| Kalsium COFI | MLD + SST + SSTsd | 39.40 | 0.76 | 0,50 | 1.10 |
| Lumba-lumba Dall | |||||
| Bahasa Inggris SWFSC | SSH + SST + kedalaman + SSHsd + SSTsd | 27.40 | 0,87 | 0,58 | 0,94 |
| Kalsium COFI | SST + SSH + SSHSD | 19.70 | 0,78 | 0.47 | 0,93 |
| Paus sirip | |||||
| Bahasa Inggris SWFSC | SSH + SST + kedalaman + MLD + SSTsd | tanggal 13.20 | 0.66 | 0.24 | 0,87 |
| Kalsium COFI | Kedalaman + SSH + SST | Tanggal 20.10 | 0.84 | 0.57 | 1.03 |
Catatan: Variabel dicantumkan dalam urutan signifikansinya, dan singkatannya adalah sebagai berikut: kedalaman, kedalaman batimetri; MLD, kedalaman lapisan campuran; SSH, tinggi permukaan laut; SSHsd, deviasi standar SSH; SST, suhu permukaan laut; SSTsd, deviasi standar SST. Semua model dikoreksi untuk upaya dengan offset untuk area efektif yang dicari (lihat teks untuk detailnya). Metrik kinerja komparatif mencakup persentase deviasi yang dijelaskan (Exp.Dev.), area di bawah kurva karakteristik operasi penerima (AUC), statistik keterampilan sebenarnya (TSS), dan rasio kepadatan yang diamati terhadap yang diprediksi untuk area studi (Obs:Pred).

3.4 Kinerja Prediktif dan Validasi Model
Kinerja prediktif bervariasi menurut spesies dan jenis model, dan data yang tersedia untuk memvalidasi prediksi model juga bervariasi menurut spesies dan lokasi dalam area studi CCE. Oleh karena itu, kemampuan model untuk memprediksi pola distribusi musim dingin dan musim semi dan memperkirakan kelimpahan absolut untuk area studi CCE dibahas secara terpisah untuk setiap spesies di bawah ini. Selain data yang diterbitkan sebelumnya yang digunakan untuk validasi silang, data musim dingin CalCOFI 2017–2020 yang digunakan untuk mengembangkan model yang diekstrapolasi secara spasial juga digunakan untuk mengevaluasi ekstrapolasi musiman model musim panas dan gugur SWFSC dalam area studi CalCOFI. Dengan pengecualian lumba-lumba biasa berparuh pendek, penampakan tahunan terlalu sedikit untuk memberikan perbandingan visual yang berarti dari pengamatan musim dingin dengan pola kepadatan yang diprediksi dari model yang diekstrapolasi secara musiman, tetapi estimasi kelimpahan tahunan 2017–2020 yang diperoleh untuk wilayah CalCOFI yang lebih kecil memberikan kumpulan data validasi silang tambahan untuk bagian area studi ini (Informasi Pendukung S.2 ).
3.4.1 Lumba-lumba Biasa Berparuh Pendek
Kelimpahan . Model yang diekstrapolasi secara musiman dan spasial menghasilkan jumlah lumba-lumba biasa berparuh pendek yang lebih banyak di dalam area studi CCE selama musim dingin dibandingkan musim semi (Gambar 3 ; Tabel 6 ), konsisten dengan estimasi berbasis desain musiman sebelumnya untuk perairan di lepas pantai California Selatan (Campbell et al. 2015 ). Kedua model juga menunjukkan pola variabilitas antartahunan yang serupa dalam kelimpahan. Misalnya, kedua model memperkirakan kelimpahan musim dingin dan musim semi terbesar pada tahun 2018 dan penurunan substansial dalam jumlah lumba-lumba biasa berparuh pendek yang ada di area studi CCE pada tahun 2020 dan 2021 (Gambar 4 , Tabel 6 ).

| Tahun 2017 | Tahun 2018 | Tahun 2019 | Tahun 2020 | Tahun 2021 | Rata-rata 2017–2021 | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Jenis | Jenis ekstrapolasi | Berlimpah | Riwayat Hidup | Berlimpah | Riwayat Hidup | Berlimpah | Riwayat Hidup | Berlimpah | Riwayat Hidup | Berlimpah | Riwayat Hidup | Berlimpah | Riwayat Hidup |
| Lumba-lumba biasa berparuh pendek | |||||||||||||
| Musim dingin | Musiman | 117.262 orang | 0,095 | 160.199 | 0,086 tahun | 140.468 | 0,088 | 118.833 | 0,093 | 68.833 | 0.121 | 121.114 tahun | 0.439 |
| Spasial | 463.940 | 0.103 | 493.511 | 0.107 | 485.886 | 0,105 | 437.901 | 0.109 | 363.556 orang | 0.107 | 448.934 | 0.253 | |
| Musim semi | Musiman | 85.254 orang | 0.109 | 107.572 orang | 0,099 tahun | 77.310 orang | 0.114 | 53.750 orang | 0.141 | 26.790 orang | 0.203 | 70.135 | 0.493 |
| Spasial | 355.067 juta | 0.109 | 364.442 orang | 0,108 | 323.892 | 0.111 | 203.145 orang | 0,139 | 166.221 orang | 0,155 | 282.553 orang | 0.340 | |
| Lumba-lumba sisi putih Pasifik | |||||||||||||
| Musim dingin | Musiman | 59.060 | 0.492 | 61.007 orang | 0,544 tahun | 64.470 orang | 0,576 tahun | 61.169 tahun | 0,578 | 36.369 orang | 0,589 | 56.415 orang | 0.405 |
| Spasial | 671.894 | 0,568 tahun | 628.988 | 0,547 tahun | 585.137 | 0.511 | 555.016 ribu | 0,509 | 736.617 juta | 0,556 | 635.352 orang | 0.817 | |
| Musim semi | Musiman | 88.792 | 0.460 | 99.656 orang | 0.405 | 73.593 orang | 0.444 | 87.498 | 0.411 | 66.140 orang | 0,501 tahun | 83.145 | 0,538 |
| Spasial | 1.122.327 tahun | 0.706 | 1.049.909 | 0.751 | 1.401.424 tahun | 0.724 | 970.674 juta | 0.691 | 968.372 orang | 0.719 | 1.102.541 tahun | 0,775 tahun | |
| Lumba-lumba Dall | |||||||||||||
| Musim dingin | Musiman | 58.999 | 0.186 | 59.354 orang | 0.161 | 46.503 orang | 0,179 tahun | 43.679 juta | 0.183 | 42.095 orang | 0.202 | 50.112 | 0.319 |
| Spasial | 240.927 juta | 0,378 tahun | 187.014 orang | 0.320 | 152.179 orang | 0.319 | 173.563 orang | 0,398 | 192.325 | 0.390 | 189.167 orang | 0.411 | |
| Musim semi | Musiman | 46.194 orang | 0.252 | 53.937 orang | 0.227 | 39.902 orang | 0.252 | 43.999 | 0.248 | 24.482 orang | 0,277 tahun | 41.703 orang | 0.334 |
| Spasial | 335.181 | 0.437 | 358.291 | 0.430 | 363.949 | 0.474 | 356.507 orang | 0.474 | 256.249 orang | 0,529 | 334.033 orang | 0.212 | |
| Paus sirip | |||||||||||||
| Musim dingin | Musiman | 5309 | 0.301 | 5838 | 0.240 | 4872 | 0.257 | 4293 | 0.241 | tahun 2906 | 0.215 | 4643 | 0.324 |
| Spasial | tahun 1219 | 0.300 | tahun 1566 | 0.288 | tahun 1207 | 0.281 | tahun 1048 | 0,278 | 924 | 0.269 | tahun 1194 | 0.223 | |
| Musim semi | Musiman | 5238 | 0.392 | 6983 | 0.420 | 4533 | 0,364 tahun | 4468 | 0,398 | tahun 1743 | 0.301 | 4593 | 0.431 |
| Spasial | tahun 1515 | 0.300 | tahun 1707 | 0.309 | tahun 1399 | 0.271 | tahun 1611 | 0.268 | 871 | 0.273 | tahun 1422 | 0.273 | |
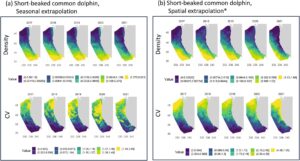
Perkiraan kelimpahan musim dingin dan musim semi untuk lumba-lumba biasa berparuh pendek yang diprediksi dari dua model tersebut sangat berbeda, namun, karena perkiraan rata-rata 2017–2021 yang diekstrapolasi secara spasial sekitar empat kali lebih besar daripada yang diprediksi oleh model yang diekstrapolasi secara musiman untuk seluruh area studi CCE (Tabel 6 ), dan hingga tiga kali lebih besar untuk area studi CalCOFI yang lebih kecil (Informasi Pendukung S.2 ). Lumba-lumba biasa berparuh pendek adalah spesies beriklim hangat yang distribusinya umumnya bergeser ke utara ke area studi CCE ketika kondisi lautan hangat dan ke selatan ke perairan lepas pantai Meksiko ketika kondisinya dingin (Barlow 2016 ; Becker et al. 2014 , 2018 , 2022 ; Dohl et al. 1986 ; Forney dan Barlow 1998 ; Heyning dan Perrin 1994 ). Dengan demikian, lebih sedikit lumba-lumba biasa berparuh pendek yang diharapkan dalam area studi CCE selama musim dingin dan musim semi daripada selama musim panas dan musim gugur; Namun, kelimpahan yang diprediksi oleh model yang diekstrapolasi secara musiman (121.114, CV = 0,439) lebih rendah daripada estimasi musim dingin berbasis desain sebelumnya untuk perairan California Selatan saja (225.949, CV = 0,32; Campbell et al. 2015 ), dan kurang dari seperdelapan estimasi kelimpahan musim panas dan gugur berbasis model sebesar 1.056.308 (CV = 0,207) untuk wilayah studi CCE (Becker et al. 2020 ). Sebaliknya, rata-rata estimasi musim dingin dan musim semi 2017–2021 dari model yang diekstrapolasi secara spasial (365.744 lumba-lumba; Tabel 6 ) serupa dengan estimasi kelimpahan 305.694 (CV = 0,340) lumba-lumba biasa yang dibuat untuk perairan dekat pantai di lepas pantai California berdasarkan data survei udara yang dikumpulkan antara Februari dan April tahun 1991 dan 1992 (Forney dan Barlow 1998 ). Mengingat peningkatan yang terdokumentasi dalam kelimpahan lumba-lumba biasa berparuh pendek di CCE selama dua dekade terakhir (Barlow 2016 ; Becker et al. 2018 ), prediksi model yang diekstrapolasi secara spasial tampaknya lebih akurat daripada prediksi yang diekstrapolasi secara musiman.
Distribusi . Kedua model memperkirakan kepadatan tertinggi lumba-lumba biasa berparuh pendek di bagian selatan area studi selama musim dingin dan semi (Gambar 3 ), konsisten dengan pergeseran selatan yang diharapkan selama periode air dingin (Barlow 2016 ; Becker et al. 2014 , 2018 , 2022 ; Dohl et al. 1986 ; Forney dan Barlow 1998 ; Heyning dan Perrin 1994 ). Model yang diekstrapolasi secara spasial mengidentifikasi pita kepadatan rendah yang membentang ke tenggara dari Point Conception (34,45° LU), memisahkan area kepadatan tinggi di bagian barat daya area studi dari wilayah kepadatan sedang di Southern California Bight (Gambar 3b ). Area ini tercakup dalam batas-batas data survei CalCOFI yang digunakan untuk membangun model (yaitu, model tidak melakukan ekstrapolasi di wilayah ini), dan pola distribusi ini serupa dengan yang sebelumnya diidentifikasi dalam penelitian lain (Becker et al. 2017 ; Campbell et al. 2015 ; Forney dan Barlow 1998 ).
Model yang diekstrapolasi secara musiman memperkirakan kepadatan tinggi di sebagian besar Southern California Bight dan juga menangkap pita kepadatan rendah yang serupa; namun, kepadatan di dekat pantai diperkirakan lebih tinggi daripada yang didokumentasikan selama studi sebelumnya, yang mengidentifikasi kepadatan terbesar di perairan lepas pantai selama musim dingin (Becker et al. 2017 ; Campbell et al. 2015 ). Lebih lanjut, perbandingan kepadatan yang diprediksi diekstrapolasi secara musiman dengan penampakan aktual dari survei musim dingin CalCOFI 2017–2020 menunjukkan model tersebut meremehkan kepadatan di area lepas pantai tempat sebagian besar penampakan terjadi (Informasi Pendukung S.2 ).
Berdasarkan perkiraan yang lebih rendah dari perkiraan kelimpahan pada model yang diekstrapolasi secara musiman dan kegagalan dalam menangkap pergeseran distribusi lumba-lumba biasa berparuh pendek di lepas pantai pada musim dingin, model yang diekstrapolasi secara spasial menunjukkan kinerja yang lebih baik di area studi ini.
3.4.2 Lumba-lumba Sisi Putih Pasifik
Kelimpahan . Kedua model tersebut memperkirakan jumlah lumba-lumba sisi putih Pasifik yang jauh lebih besar di area studi CCE selama musim semi dibandingkan musim dingin (Gambar 5 , Tabel 6 ). Hasil ini konsisten dengan estimasi kelimpahan berbasis desain musiman dari Campbell et al. ( 2015 ), dan dengan hasil survei udara yang dilakukan di lepas pantai Oregon dan Washington antara April 1989 dan September 1990 yang mendokumentasikan kepadatan terbesar selama akhir musim semi dan awal musim panas dan kepadatan terendah selama musim dingin (Green et al. 1992 ). Variabilitas antartahunan dalam kelimpahan juga tampak untuk kedua set model (Tabel 6 ).
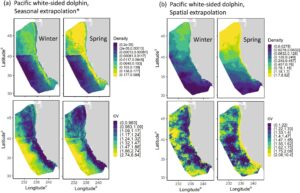
Akan tetapi, ada perbedaan orde-magnitudo dalam kelimpahan yang diprediksi antara model yang diekstrapolasi secara musiman dan yang diekstrapolasi secara spasial (Tabel 6 ). Prediksi model yang diekstrapolasi secara spasial terendah untuk seluruh area studi CCE selama musim dingin dan semi adalah 15 hingga 44 kali lebih tinggi daripada estimasi musim panas dan gugur 2018 berbasis model yang diterbitkan sebelumnya sebesar 39.999 (CV = 0,222; Becker et al. 2020 ). Kelimpahan spesies beriklim dingin ini diperkirakan akan meningkat di area studi CCE selama musim dingin (Becker et al. 2014 ; Campbell et al. 2015 ; Forney dan Barlow 1998 ; Green et al. 1992 ); namun, besarnya peningkatan yang diprediksi oleh model yang diekstrapolasi secara spasial terlalu besar untuk diperhitungkan oleh pergeseran distribusi atau potensi peningkatan populasi. Estimasi kelimpahan rata-rata musim semi tahun 2017–2021 dari model yang diekstrapolasi secara spasial adalah 1.102.541 (CV = 0,775), yang kira-kira 8% lebih tinggi dari estimasi sebelumnya di seluruh wilayah yaitu 931.000 (CV = 0,90) lumba-lumba sisi putih Pasifik di seluruh Pasifik Utara (Buckland 1993 ). Estimasi kelimpahan yang diperoleh dari model yang diekstrapolasi secara spasial tampaknya tidak masuk akal secara ekologis.
Estimasi kelimpahan rata-rata 2017–2020 dari model yang diekstrapolasi secara musiman dan model yang diekstrapolasi secara spasial untuk area studi CalCOFI yang lebih kecil (area tempat model yang terakhir tidak melakukan ekstrapolasi) dibandingkan dengan estimasi berbasis desain Campbell et al. ( 2015 ) untuk musim dingin (16.718) dan musim semi (23.983) untuk bagian selatan area studi CalCOFI (Informasi Pendukung S.2 ). Hal ini menunjukkan bahwa estimasi kelimpahan substansial di seluruh CCE yang diperoleh dari model yang diekstrapolasi secara spasial terutama disebabkan oleh ekstrapolasi model di bagian utara area studi.
Distribusi . Kedua model tersebut memperkirakan kepadatan lumba-lumba sisi putih Pasifik terbesar di seluruh perairan lepas pantai Washington dan Oregon (yaitu, utara 42° LU) dan meluas ke selatan sepanjang pita pantai sempit ke Point Conception (Gambar 5 ). Model yang diekstrapolasi secara musiman memperkirakan wilayah yang jelas dengan kepadatan yang lebih rendah di sepanjang sebagian besar Pantai Barat AS, yang konsisten dengan penelitian sebelumnya yang mendokumentasikan sebagian besar lumba-lumba sisi putih Pasifik di perairan lereng dan lepas pantai daripada di perairan landas yang lebih dangkal (Forney dan Barlow 1998 ; Green et al. 1992 ). Perbedaan dalam pola distribusi pesisir ini juga terlihat dalam pola distribusi musim dingin gabungan tahunan 2017–2021 yang diprediksi (Gambar 6 ). Model yang diekstrapolasi secara musiman juga meramalkan kepadatan yang relatif tinggi hingga menengah di seluruh Teluk California Selatan, konsisten dengan pergerakan ke selatan yang terdokumentasi dan beberapa penampakan spesies ini di perairan selatan Point Conception pada musim dingin (Becker et al. 2014 ; Campbell et al. 2015 ; Forney dan Barlow 1998 ; Green et al. 1992 ).

Berdasarkan estimasi kelimpahan yang sangat tinggi dan tidak realistis yang diprediksi oleh model yang diekstrapolasi secara spasial, dan ketidaksesuaian dalam pola distribusi yang diprediksi di sepanjang pantai, kami menganggap model yang diekstrapolasi secara musiman telah menunjukkan kinerja yang lebih baik untuk lumba-lumba sisi putih Pasifik di area studi ini.
3.4.3 Lumba-lumba Dall
Kelimpahan . Hasil kelimpahan musiman dari dua model lumba-lumba Dall tidak konsisten. Model yang diekstrapolasi secara spasial menunjukkan jumlah lumba-lumba Dall yang jauh lebih besar di dalam area studi CCE selama musim semi daripada selama musim dingin, sementara model yang diekstrapolasi secara musiman menunjukkan lebih sedikit variabilitas antara periode musim dingin dan musim semi, meskipun kelimpahan secara umum diprediksi lebih besar di musim dingin (Tabel 6 ). Mirip dengan hasil untuk lumba-lumba sisi putih Pasifik, estimasi kelimpahan absolut berbasis model untuk seluruh area studi CCE sangat berbeda antara dua set data, dengan model yang diekstrapolasi secara spasial memprediksi lebih banyak lumba-lumba Dall selama musim dingin dan musim semi daripada model yang diekstrapolasi secara musiman (Tabel 6 ). Berdasarkan penelitian sebelumnya yang menghasilkan estimasi kelimpahan sebesar 16.498 (CV = 0,608) selama musim panas dan gugur 2018 (Becker et al. 2020 ) dan 26.111 (CV = 0,296) selama musim dingin dan semi 1991–1992 (Forney dan Barlow 1998 ), estimasi kelimpahan total yang diekstrapolasi secara spasial yang rata-rata mencapai 261.600 ekor lumba-lumba selama musim dingin dan semi (Tabel 6 ) kemungkinan bias tinggi, khususnya di bagian utara area penelitian di mana estimasi kepadatan mencapai 1,12 ekor lumba-lumba km −2 (Gambar 7b ).

Estimasi kelimpahan tahunan 2017–2020 untuk area studi CalCOFI yang lebih kecil serupa untuk model yang diekstrapolasi secara musiman dan model yang diekstrapolasi secara spasial, dengan estimasi rata-rata masing-masing 12.907 dan 17.502 lumba-lumba (Informasi Pendukung S.2 ). Dengan demikian, estimasi kelimpahan CCE yang jauh lebih besar untuk model yang diekstrapolasi secara spasial (Tabel 6 ) dapat dikaitkan dengan ekstrapolasi model di bagian utara area studi.
Distribusi . Pola distribusi musim dingin dan musim semi yang diprediksi oleh kedua model secara umum serupa, dengan kepadatan terbesar diprediksi untuk bagian utara area studi dan kepadatan terendah di barat daya, dengan kepadatan tingkat menengah hingga tinggi meluas ke selatan di sepanjang wilayah dekat pantai ke Southern California Bight (Gambar 7 ). Kedua model juga menunjukkan wilayah kepadatan lebih rendah di sepanjang pantai Washington dan Oregon, konsisten dengan data penampakan musim dingin dan musim semi dan prediksi model kepadatan relatif yang menunjukkan bahwa sebagian besar lumba-lumba Dall diamati di perairan lereng dan lepas pantai, dan paling sedikit di landas kontinen (Green et al. 1992 ; Menza et al. 2016 ). Menariknya, pola distribusi yang diprediksi oleh model yang diekstrapolasi secara musiman untuk area studi selatan 38° LU sangat mirip dengan yang diperoleh dari SDM musim dingin dan musim semi sebelumnya yang dikembangkan dengan sebagian kecil data penampakan CalCOFI yang digunakan di sini (Becker et al. 2017 ).
Pola distribusi gabungan musim dingin tahunan 2017–2021 yang diprediksi untuk lumba-lumba Dall serupa dengan yang dijelaskan di atas, dengan variabilitas antartahunan tampak jelas untuk kedua set model (Gambar 8 ). Untuk gabungan musim dingin, kedua model memprediksi jumlah lumba-lumba Dall terendah yang ada di area studi CCE selama tahun 2021 dibandingkan dengan 4 tahun sebelumnya (Gambar 8 , Tabel 6 ).
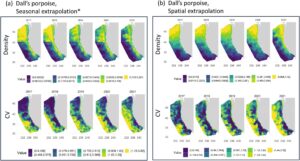
Berdasarkan estimasi kelimpahan ekstrem yang diprediksi oleh model yang diekstrapolasi secara spasial, khususnya di bagian utara area studi, model yang diekstrapolasi secara musiman tampaknya menunjukkan kinerja yang lebih baik untuk lumba-lumba Dall di area studi ini.
3.4.4 Paus Sirip
Kelimpahan . Paus sirip ditemukan sepanjang tahun di Southern California Bight, meskipun jumlahnya jauh lebih banyak selama musim panas dan gugur (Campbell et al. 2015 ; Carretta et al. 1995 ; Dohl et al. 1978 , 1983 ; Forney and Barlow 1998 ). Kedua estimasi kelimpahan yang diprediksi oleh model konsisten dengan pola musiman ini, karena estimasi kelimpahan musim dingin dan semi jauh lebih rendah daripada estimasi kelimpahan paus sirip musim panas dan gugur terkini sebesar 11.065 paus untuk area studi CCE (Becker et al. 2020 ). Akan tetapi, model yang diekstrapolasi secara musiman memprediksi jumlah paus sirip tiga hingga empat kali lebih banyak di area studi CCE selama musim dingin daripada model yang diekstrapolasi secara spasial (Tabel 6 ), dan sekitar empat kali lebih banyak paus sirip di area studi CalCOFI (Informasi Pendukung S.2 ). Estimasi kelimpahan rata-rata 2017–2020 sebanyak 980 paus sirip yang diperoleh dari model ekstrapolasi spasial untuk area studi CalCOFI (di mana tidak ada ekstrapolasi) lebih konsisten dengan estimasi berbasis desain untuk bagian selatan area studi CalCOFI (155 untuk musim dingin dan 432 untuk musim semi; Campbell et al. 2015 ) daripada yang diperoleh dari model ekstrapolasi musiman (4031 paus sirip). Perbandingan ini menunjukkan bahwa model ekstrapolasi musiman menghasilkan estimasi kelimpahan tinggi yang bias untuk area studi.
Distribusi . Kedua model tersebut memperkirakan kepadatan paus sirip tertinggi di selatan Point Conception (34,45° LU) selama musim dingin dan semi, konsisten dengan pola tempat tinggal mereka yang telah ditetapkan sepanjang tahun di lepas pantai California Selatan (Gambar 9 ; Campbell et al. 2015 ; Carretta et al. 1995 ; Dohl et al. 1978 , 1983 ; Falcone et al. 2022 ; Forney dan Barlow 1998 ). Pola distribusi yang diprediksi oleh model yang diekstrapolasi secara spasial lebih cocok dengan pola kejadian yang telah dijelaskan untuk spesies ini dalam penelitian sebelumnya, dengan mayoritas penampakan musim dingin dan semi di perairan dekat pantai dan di atas landas kontinen (Gambar 9b ; Campbell et al. 2015 , Falcone et al. 2022 ; Forney dan Barlow 1998 ; Scales et al. 2017 ). Selama musim panas dan gugur, paus sirip cenderung lebih tersebar di lepas pantai dan distribusinya meluas lebih jauh ke utara; namun, kepadatan tinggi belum terdokumentasikan untuk bagian barat daya area studi (Becker et al. 2020 ; Scales et al. 2017 ), di mana model yang diekstrapolasi secara musiman memperkirakan kepadatan tertinggi. Kedua model juga memperkirakan beberapa kepadatan sedang di lepas pantai Washington dan Oregon, konsisten dengan model sebelumnya tentang kesesuaian habitat dan data penampakan lainnya (Derville et al. 2022 ; Green et al. 1992 ; Scales et al. 2017 ).

Mirip dengan model untuk tiga spesies lainnya, pola distribusi gabungan musim dingin tahunan 2017–2021 untuk paus sirip secara umum mirip dengan yang dijelaskan di atas untuk masing-masing model musiman, dengan variabilitas antartahunan dalam kelimpahan yang ditunjukkan oleh kedua prediksi model (Gambar 10 , Tabel 6 ). Kedua model memperkirakan paus sirip yang jauh lebih sedikit di area studi CCE pada tahun 2021, khususnya di lepas pantai Washington dan Oregon, dan jumlah paus terbanyak pada tahun 2018 (Gambar 10 , Tabel 6 ).

Berdasarkan estimasi kelimpahan tinggi yang tampak bias yang diperoleh dari model yang diekstrapolasi secara musiman, dan ketidaksesuaian dalam pola distribusi yang diprediksi di Southern California Bight, kami menganggap model yang diekstrapolasi secara spasial telah menunjukkan kinerja yang lebih baik untuk paus sirip di wilayah studi ini.
4 Diskusi
Penggunaan ekstrapolasi untuk SDM laut telah menjadi topik beberapa penelitian yang menggambarkan tantangan yang terlibat. Ekstrapolasi mungkin tidak akurat jika dilakukan di seluruh ekosistem (Mannocci et al. 2020 ; Redfern et al. 2017 ), tetapi ketika dilakukan dengan hati-hati dan dalam ekoregion yang sama, mereka dapat memberikan informasi yang sangat dibutuhkan untuk pengelolaan dan konservasi spesies laut. Ketika data spesifik waktu dan lokasi tidak tersedia, ekstrapolasi yang cermat dapat menginformasikan keputusan pengelolaan yang seharusnya dibuat tanpa adanya informasi tentang distribusi dan kepadatan spesies (Mannocci et al. 2017 ). Namun, penting untuk memahami kapan dan dalam keadaan apa ekstrapolasi dapat diterima versus bias yang tidak dapat diterima, dan metode mana yang menghasilkan hasil yang paling dapat diandalkan. Dalam penelitian ini kami membandingkan prediksi model yang diekstrapolasi secara spasial dan musiman untuk empat spesies dalam ekosistem yang sangat dinamis untuk meningkatkan pemahaman kita dan meningkatkan aplikasi masa depan dari prediksi SDM yang diekstrapolasi.
Mengingat meningkatnya permintaan untuk menggunakan SDM untuk membuat prediksi untuk wilayah geografis yang sampelnya tidak mencukupi dan selama periode waktu baru, perangkat telah dikembangkan untuk menilai besarnya prediksi yang diekstrapolasi, mengurangi atau menghilangkan ekstrapolasi sejauh mungkin, melakukan validasi silang prediksi model jika data tersedia, dan secara transparan menyajikan ketidakpastian dalam prediksi (Conn et al. 2015 ; Mannocci et al. 2017 ; Sequeira et al. 2018 ). Dalam studi ini, ukuran penghindaran yang sederhana namun sering kali efektif digunakan untuk mengevaluasi dan membuang prediksi model apa pun yang dibuat di luar ruang multivariat data yang digunakan untuk mengembangkan model asli (Bouchet et al. 2019 ). Prediksi model kemudian divalidasi silang sejauh mungkin menggunakan data yang tersedia dalam area studi, dan ketidakpastian dicirikan menggunakan teknik yang baru dikembangkan untuk memperoleh ukuran varians yang komprehensif dalam prediksi GAM (Miller et al. 2022 ).
Istilah tahun berkelanjutan disertakan sebagai kovariat dalam SDM sebelumnya yang dikembangkan menggunakan data survei SWFSC CCE yang sama, dan berhasil menangkap tren populasi untuk lumba-lumba biasa berparuh pendek dan paus sirip yang kelimpahannya di musim panas dan gugur telah meningkat secara substansial selama periode waktu 1991 hingga 2018. Tahun secara khusus tidak ditawarkan kepada model dalam studi ini karena tahun dapat bertindak sebagai proksi untuk variabel dinamis, yang dalam hal ini dapat mengacaukan ekstrapolasi spasial dan musiman. Potensi “kebisingan” yang akan diperkenalkan oleh istilah tahun ke dalam proses ekstrapolasi yang sudah rumit tidak dianggap tepat untuk analisis ini. Untuk dua spesies dengan peningkatan populasi yang terdokumentasi dalam area studi di musim panas dan gugur, jika ada peningkatan serupa di musim dingin dan semi, estimasi kepadatan yang dihasilkan mungkin bias rendah.
Untuk meningkatkan kemampuan prediktif SDM, studi sebelumnya telah menggabungkan set data yang berbeda untuk membangun model gabungan tunggal dalam upaya untuk menangkap kekuatan set data individual (misalnya, Redfern et al. 2017 ). Strategi ini tidak dianggap tepat dalam kasus ini mengingat perubahan musiman dinamis yang terdokumentasi dalam kelimpahan dan distribusi cetacea di CCE, khususnya di Southern California Bight. Menggabungkan set data musim panas dan gugur dan musim dingin dan musim semi secara inheren mengasumsikan bahwa bentuk fungsional variabel habitat akan konsisten antara musim panas dan gugur CCE SWFSC dan set data musim dingin dan musim semi khusus CalCOFI California Selatan (yang seperti yang ditunjukkan untuk lumba-lumba biasa berparuh pendek dan paus sirip, sangat berbeda untuk variabel kedalaman). Selain itu, data SWFSC memiliki penampakan yang jauh lebih banyak dan akan mendominasi sinyal dalam model. (2019) menyajikan pendekatan pemodelan hierarkis terpadu yang secara teori dapat memungkinkan survei musim panas dan gugur SWFSC dan survei musim dingin dan semi CalCOFI untuk dimasukkan dalam satu model. Namun, pendekatan ini tidak dianggap tepat untuk analisis kami karena hal ini akan meningkatkan kompleksitas model di luar apa yang didukung oleh ukuran sampel terbatas selama survei CalCOFI. Lebih jauh, ada perbedaan oseanografi dan ekologi yang nyata antara area studi CalCOFI dan CCE yang lebih luas (misalnya, Daily et al. 1993 ; Miller 2023 ) yang dapat membingungkan pendekatan semacam itu. Akhirnya, menggabungkan kumpulan data tidak akan memungkinkan perbandingan ekstrapolasi musiman versus spasial.
Berdasarkan analisis kami, prediksi yang diekstrapolasi secara spasial lebih baik untuk lumba-lumba biasa berparuh pendek dan paus sirip, sementara prediksi model yang diekstrapolasi secara musiman lebih baik untuk lumba-lumba sisi putih Pasifik dan lumba-lumba Dall. Mengingat variabilitas dalam hasil ini, penting untuk mempertimbangkan karakteristik khusus spesies atau khusus area studi apa yang mungkin memengaruhi hasil ini. Dua faktor utama tampak jelas: (1) sejauh mana model menangkap habitat inti spesies, yaitu, apakah kurva fungsional kovariat menangkap puncak; dan (2) efek prediktor statis (misalnya, kedalaman, jarak ke pantai, jarak ke isobath, dll.) yang menjangkarkan spesies dalam ruang geografis dan mencegah identifikasi pergeseran distribusi dalam ruang atau periode waktu baru. Ini dijelaskan secara lebih rinci di bawah ini.
4.1 Habitat Inti
Habitat inti untuk lumba-lumba biasa berparuh pendek dan paus sirip dalam area studi CCE adalah Teluk California Selatan (Becker et al. 2022 ; Scales et al. 2017 ), yang tersampel dengan baik oleh survei CalCOFI. Sebaliknya, distribusi inti lumba-lumba sisi putih Pasifik beriklim dingin dan lumba-lumba Dall berada jauh di utara area survei CalCOFI, dengan Ekosistem Arus California selatan mewakili batas jangkauan selatan mereka di Pasifik Utara bagian timur (lihat misalnya, Hamilton et al. 2009 ). Model CalCOFI untuk kedua spesies beriklim dingin ini dengan demikian didasarkan pada bagian selatan habitat mereka, dengan bentuk fungsional hanya mencerminkan sebagian kecil dari jangkauan habitat mereka yang lebih besar (yaitu, ekor habitat optimal setiap spesies). Misalnya, bentuk fungsional SST dalam model CalCOFI untuk lumba-lumba sisi putih Pasifik dan lumba-lumba Dall adalah linear negatif, dengan jumlah lumba-lumba/lumba-lumba tertinggi ditemukan di perairan terdingin di area studi (Gambar 2b,c ). Ketika model-model ini digunakan untuk memprediksi kondisi musim dingin dan musim semi di seluruh area studi CCE, kejadian luas air dingin di lepas pantai Washington dan Oregon kemungkinan menyebabkan prediksi kepadatan ekstrem di bagian utara area studi (Gambar 5b dan 7b ). Efek linear hanya mencerminkan sebagian plot fungsional dari ~10°C hingga 18°C, bukan hubungan unimodal yang ditangkap oleh model musim panas dan gugur SWFSC yang mencakup suhu dari ~10°C hingga 23°C.
Model SWFSC mencakup habitat inti untuk lumba-lumba sisi putih Pasifik dan lumba-lumba Dall di perairan utara area studi, meskipun di musim panas dan gugur, tetapi kumpulan data memiliki sampel yang cukup di perairan terdingin untuk musim-musim ini dan kurva fungsional untuk SST menunjukkan penurunan (lumba-lumba sisi putih Pasifik) dan respons monotonik (lumba-lumba Dall) dalam kepadatan di dalam perairan yang lebih dingin dari sekitar 13°C (Gambar 2b,c ). Nilai SST terendah yang diambil sampelnya tidak jauh berbeda antara kedua kumpulan data (9,96°C untuk model SWFSC versus 9,17°C untuk model CalCOFI; Tabel 4 ), dan keduanya lebih hangat daripada perairan terdingin yang biasanya ditemui di lepas pantai Washington di musim dingin (misalnya, sekitar 8°C, mengingat variabilitas antartahunan; Legaard et al. 2006 ; Venegas et al. 2008 ). Akan tetapi, meskipun kumpulan data CalCOFI menyertakan nilai SST yang lebih dingin daripada kumpulan data SWFSC, hubungan yang lebih unimodal yang ditangkap oleh model musim panas dan gugur SWFSC mampu mendefinisikan dengan lebih baik hubungan habitat SST-spesies di habitat inti spesies ini dalam area studi CCE. Prediksi yang diekstrapolasi secara musiman untuk kedua spesies ini menghasilkan estimasi kelimpahan yang wajar dan pola distribusi spasial yang konsisten dengan yang didokumentasikan untuk spesies ini di musim dingin (Forney dan Barlow 1998 ; Green et al. 1992 ; Menza et al. 2016 ).
Hasil ini menggambarkan dengan kuat bahwa, bahkan ketika menggunakan pemotongan sebagai metode utama untuk menghilangkan atau mengurangi potensi artefak ekstrapolasi, prediksi yang dihasilkan bisa sangat bias. Metode penghindaran yang digunakan dalam penelitian ini membuang prediksi yang dibuat di luar rentang kovariat yang digunakan untuk mengembangkan model CalCOFI (yaitu, dalam contoh di atas piksel apa pun dengan suhu air < 9,17°C tidak digunakan; Tabel 4 ); namun, estimasi kepadatan yang dihasilkan dari model yang diekstrapolasi secara spasial untuk lumba-lumba sisi putih Pasifik dan lumba-lumba Dall masih sangat bias-tinggi.
4.2 Prediktor Statis
Batimetri telah terbukti menjadi prediktor efektif dalam SDM cetacea di area studi CCE (Abrahms et al. 2019 ; Becker et al. 2016 , 2020 ; Forney 2000 ; Forney et al. 2012 , Hazen et al. 2017 ), dan kedalaman ditawarkan sebagai kovariat potensial dalam studi ini, serupa dengan pendekatan yang diambil oleh Mannocci et al. ( 2017 ). Kedalaman juga disertakan dalam SDM sebelumnya (Becker et al. 2018 ) untuk delapan spesies cetacea (termasuk empat spesies yang dibahas dalam studi ini) yang berhasil digunakan untuk memprediksi pergeseran distribusi substansial selama tahun anomali hangat baru di area studi CCE. Pendekatan ini mengasumsikan bahwa “habitat kedalaman” yang disukai spesies akan tetap cukup konstan dari waktu ke waktu dan ruang. Akan tetapi, seperti yang ditunjukkan di sini, penyertaan kedalaman dapat menghambat prediksi baru jika suatu spesies mengalihkan distribusi ke perairan dengan batimetri yang sangat berbeda dalam ruang atau periode waktu baru. Untuk lumba-lumba biasa berparuh pendek dan paus sirip, bentuk fungsional kedalaman dalam model musim panas dan gugur SWFSC menghasilkan beberapa pola distribusi yang tidak cocok yang diprediksi untuk musim dingin dan musim semi, khususnya di habitat inti mereka di Teluk California Selatan, suatu area yang dicirikan oleh batimetri yang kompleks (Hickey 1979 ).
Bahasa Indonesia: Di musim panas dan gugur dalam area studi CCE, lumba-lumba biasa berparuh pendek menunjukkan distribusi kedalaman bimodal, dengan kepadatan tertinggi terjadi di perairan yang relatif dangkal di seluruh Southern California Bight dan juga di perairan dalam (sekitar 4000 m dalam) di lepas pantai California tengah dan utara (Becker et al. 2016 , 2018 , 2020 ). Pergeseran signifikan ke selatan dalam distribusi lumba-lumba biasa berparuh pendek terjadi di musim dingin dan semi, yang berhasil ditangkap oleh prediksi model SWFSC (Gambar 3a ). Di Southern California Bight, ada juga pergeseran yang jelas dalam distribusi lumba-lumba biasa berparuh pendek di lepas pantai ke perairan pelagis di bagian barat daya area studi CCE (Becker et al. 2017 ; Campbell et al. 2015 ). Pergeseran dari perairan di atas landas kontinen tidak tertangkap dalam prediksi musim dingin dan semi dari model musim panas dan gugur SWFSC, kemungkinan karena dimasukkannya fungsi kedalaman bimodal yang menghasilkan prediksi lebih tinggi di dekat pantai (Gambar 2a ).
Situasi serupa terjadi pada paus sirip, spesies yang muncul sepanjang tahun di Southern California Bight tetapi memiliki distribusi yang lebih luas di musim panas dan gugur (Becker et al. 2022 ; Forney dan Barlow 1998 ; Scales et al. 2017 ). Mirip dengan lumba-lumba biasa berparuh pendek, model musim panas dan gugur SWFSC mencakup fungsi kedalaman bimodal (Gambar 2d ), yang menghasilkan prediksi kepadatan tinggi di lepas pantai untuk musim dingin dan semi, tidak konsisten dengan distribusi musim dingin mereka di perairan dekat pantai di atas landas kontinen (Campbell et al. 2015 ; Falcone et al. 2022 ; Forney dan Barlow 1998 ; Scales et al. 2017 ).
4.3 Model Ekstrapolasi Musiman vs. Model Ekstrapolasi Spasial
Berdasarkan hasil studi ini, prediksi musim dingin dan musim semi terbaik dihasilkan oleh model ekstrapolasi spasial untuk lumba-lumba biasa berparuh pendek dan paus sirip, dan oleh model ekstrapolasi musiman untuk lumba-lumba sisi putih Pasifik dan lumba-lumba Dall. Prediksi model ini mampu menangkap perbedaan variabel musiman dalam kelimpahan dan distribusi yang konsisten dengan pola yang diketahui atau diduga. Prediksi model yang dipilih untuk keempat spesies ini memberikan prediksi kepadatan eksplisit spasial yang sangat berbeda dari musim panas dan gugur, dan dapat menginformasikan kebutuhan manajemen untuk musim dingin di area studi CCE. Meskipun ketidakpastian yang lebih besar, model-model ini lebih disukai daripada alternatif menggunakan model musim panas dan gugur untuk mencerminkan pola kelimpahan dan distribusi tahunan. Sampai SDM khusus musim dingin dapat dikembangkan, dan dengan penerapan kehati-hatian, model-model ini menyediakan alat untuk menilai risiko dan mengembangkan langkah-langkah mitigasi untuk spesies ini di area studi CCE di musim dingin dan musim semi.
4.4 Studi Masa Depan
Bahasa Indonesia: Untuk mengurangi potensi bias dari ekstrapolasi, kami menggunakan pemotongan, yaitu, kami membuang prediksi model apa pun yang dibuat di luar ruang multivariat data yang digunakan untuk mengembangkan model asli (Bouchet et al. 2019 ). Pendekatan ini dipilih karena studi pemodelan sebelumnya yang menggunakan subset data SWFSC CCE menghasilkan prediksi yang sangat tinggi dari prediksi yang tidak dibatasi (Becker et al. 2012 ). Lebih jauh, karena tujuan dari studi ini adalah untuk membandingkan ekstrapolasi musiman versus spasial, kami ingin menggunakan metode konsisten yang dapat diterapkan pada kedua set model daripada menerapkan pendekatan khusus model yang dapat mengacaukan hasil.
Metode pemotongan menghilangkan piksel dari permukaan kepadatan rata-rata akhir, dan dengan demikian rata-rata hanya didasarkan pada irisan waktu dalam rentang lainnya. Bergantung pada persentase piksel yang dihilangkan dan sejauh mana mereka dikelompokkan bersama secara spasial atau temporal, ini dapat memperkenalkan bias dalam prediksi. Metode alternatif untuk menangani ekstrapolasi dalam prediksi SDM mencakup “penjepitan” atau “pembatasan” nilai kovariat ke ekstrem sampelnya (Bouchet et al. 2019 ), yang akan mempertahankan semua piksel prediksi. Namun, pendekatan penjepitan atau pembatasan bergantung pada penilaian direksionalitas setiap kurva respons pada titik pemotongan (Guevara et al. 2018 ), dan akan mempersulit interpretasi hasil dalam studi ini yang diperkenalkan dengan menggunakan pendekatan ekstrapolasi khusus model variabel. Metode terbaik untuk menangani ekstrapolasi kemungkinan bergantung pada seberapa banyak hubungan unimodal (yang diduga) telah ditangkap. Untuk hubungan linear, ini mungkin hanya satu sisi dari kurva unimodal, jadi penjepitan mungkin lebih baik; Namun, jika plot fungsional menangkap sebagian besar hubungan unimodal, maka mengizinkan ekstrapolasi mungkin lebih baik (misalnya, Guevara et al. 2018 ). Mengevaluasi metode alternatif untuk menangani ekstrapolasi, seperti penjepitan, akan menjadi fokus analisis mendatang dalam bidang studi ini.
5 Kesimpulan
SDM kini telah ditetapkan dengan kuat sebagai alat konservasi dan pengelolaan yang efektif, dan penggunaan prediksi model yang diekstrapolasi untuk wilayah dan periode waktu yang miskin data diharapkan akan meningkat di masa mendatang. Studi ini telah menunjukkan bahwa ada faktor khusus spesies dan khusus area studi yang perlu dipertimbangkan saat membuat prediksi model yang diekstrapolasi. Pertama, ekstrapolasi spasial cenderung menghasilkan prediksi yang sangat bias saat SDM dikembangkan di luar area habitat inti (yaitu, di ekor nilai kovariat optimum spesies). Bahkan saat membatasi prediksi pada ruang univariat dan multivariat yang digunakan untuk membangun model, pola dapat dibesar-besarkan di habitat inti. Saat spesies diketahui atau diduga menunjukkan pergeseran temporal dalam distribusi, variabel statis harus dievaluasi dengan hati-hati sebelum memasukkan model. Dalam semua kasus, ekstrapolasi harus selalu ditafsirkan dengan hati-hati, bahkan saat metode telah digunakan untuk mengendalikan prediksi. Saat data tambahan dikumpulkan dalam area studi ini selama musim dingin, validasi silang yang lebih komprehensif harus dilakukan untuk memberikan wawasan tambahan tentang penggunaan dan penerapan prediksi SDM yang diekstrapolasi di masa mendatang.


