Abstrak
- Variasi individu dalam perkembangan dapat berdampak kuat pada kepadatan populasi melalui dampaknya pada demografi. Ketika variasi dalam perkembangan ini disebabkan oleh plastisitas perkembangan, dampak yang dihasilkan pada kepadatan populasi dicirikan sebagai ‘dinamika eko-fenotipik’. Jika plastisitas pada gilirannya peka terhadap variasi dalam kepadatan populasi, ‘umpan balik eko-fenotipik’ dapat muncul. Ketika perkembangan individu bergantung pada kepadatan karena keterbatasan sumber daya, kami memperkirakan umpan balik eko-fenotipik antara laju perkembangan dan kepadatan populasi akan meluas. Namun, bagaimana tepatnya plastisitas dalam laju perkembangan, yang disebut ‘heterokairy’ berinteraksi dengan kepadatan populasi untuk memengaruhi dinamika eko-fenotipik masih belum jelas.
- Di sini, kami menyediakan titik awal konseptual dan empiris untuk menilai peran heterokairy dalam dinamika populasi eko-fenotipik. Kami menyajikan kerangka konseptual untuk mengidentifikasi bagaimana heterokairy memengaruhi, dan dipengaruhi oleh, kepadatan populasi. Kerangka ini divalidasi menggunakan bukti empiris yang ada, dengan fokus pada polifenisme yang terdiri dari fenotipe alternatif yang berbeda dalam tingkat perkembangan: ‘polifenisme heterokairik’.
- Kami menemukan bahwa polifenisme heterokairik biasanya terdiri dari fenotipe yang berkembang cepat (‘minor’) dan yang berkembang lambat (‘mayor’) yang lazim di antara ektotermik. Bagaimana heterokairik membentuk umpan balik eko-fenotipik bergantung pada apakah minor (atau mayor) sebagian besar diekspresikan pada kepadatan rendah (atau tinggi), atau sebaliknya. Sistem minor-pada-kepadatan-rendah, seperti yang ditemukan pada salamander, kutu daun, dan nematoda, mencerminkan prediksi riwayat hidup klasik bahwa populasi yang tumbuh, kepadatan rendah, mendukung waktu perkembangan yang singkat, sementara populasi kepadatan tinggi mendukung perkembangan yang lebih lambat. Sebagai alternatif, sistem minor-pada-kepadatan-tinggi memungkinkan juvenil yang tertekan sumber daya, paling umum pada kepadatan tinggi, untuk tetap dewasa tetapi sebagai minor dengan mengurangi biaya perkembangan dan metabolisme. Kami mengusulkan respons yang berlawanan terhadap kepadatan tersebut dapat mendorong dinamika dan umpan balik eko-fenotipik yang berbeda.
- Pekerjaan kami menyoroti peran penting yang dimainkan oleh perkembangan dalam pembentukan fenotipe, dan konsekuensi yang dapat ditimbulkannya bagi dinamika (meta)populasi dan evolusi fenotipe. Kami menyerukan pergeseran dari fokus pada hasil perkembangan di masa dewasa ke proses perkembangan yang membentuk fenotipe. Pada akhirnya, hal ini dapat mengarah pada pendekatan yang lebih mekanistik, dan dengan demikian prediktif, untuk memahami peran adaptif dari berbagai laju perkembangan dalam konteks ekologi dan evolusi.
Baca Ringkasan Bahasa Sederhana gratis untuk artikel ini di blog Jurnal.
1. PENDAHULUAN
Ukuran dan komposisi populasi diatur oleh faktor-faktor endogen yang bergantung pada kepadatan (lihat ‘Glosarium’ untuk definisi istilah yang dicetak tebal), dan faktor-faktor eksogen yang tidak bergantung pada kepadatan (Brook & Bradshaw, 2006 ; Sibly et al., 2005 ). Faktor-faktor eksogen mencakup, misalnya, suhu, presipitasi, atau salinitas (Hernandez Plaza et al., 2012 ), yang merupakan faktor-faktor yang tidak terpengaruh oleh kepadatan. Namun, faktor-faktor seperti ketersediaan makanan, oksigen, dan habitat bersifat endogen dan dapat habis karena kepadatan yang berlebihan atau dipengaruhi oleh komposisi dan struktur populasi (Turchin, 2003 ). Pada skala waktu kontemporer (Hendry & Kinnison, 1999 ), dalam masa hidup individu, faktor-faktor endogen dapat memengaruhi fenotipe individu. Salah satu cara tersebut adalah melalui plastisitas perkembangan , karena faktor endogen dapat bertindak sebagai isyarat untuk pengembangan fenotipe plastik (West-Eberhard, 2003 ). Namun, kita hanya tahu sedikit tentang bagaimana plastisitas yang dipengaruhi oleh faktor endogen memengaruhi dinamika populasi (Croll & de Roos, 2022 ).
Jika plastisitas memengaruhi riwayat hidup individu, yaitu pertumbuhan, reproduksi, dan mortalitasnya, plastisitas dapat menimbulkan konsekuensi populasi yang signifikan. Karena riwayat hidup individu bergantung pada lingkungan sumber daya, yang pada gilirannya dapat dipengaruhi oleh kepadatan populasi, plastisitas dan riwayat hidup dapat menciptakan umpan balik eko-fenotipik (Gibert et al., 2022 ). Misalnya, kita akan memperkirakan variasi plastis dalam laju pertumbuhan dan waktu peristiwa perkembangan, yang disebut heterokairy (Rundle & Spicer, 2016 ; Spicer et al., 2018 ), akan memengaruhi pertumbuhan populasi karena secara langsung memengaruhi proses demografi, seperti usia dewasa dan waktu reproduksi (Gibert et al., 2022 , 2023 ; Spicer et al., 2018 ). Hal ini terbukti pada banyak spesies salamander (misalnya salamander harimau Arizona Ambystoma mavortium nebulosum , Lackey et al., 2019 ) yang mengekspresikan paedomorphosis , retensi karakteristik juvenil. Pada kepadatan populasi rendah, individu mengekspresikan paedomorphosis untuk bereproduksi lebih awal dalam hidup dan mendapat keuntungan dari ketersediaan makanan yang tinggi (ten Brink et al., 2020 ). Namun, seiring peningkatan kepadatan, lebih banyak individu akan memperpanjang perkembangan untuk bermetamorfosis dan menyebar untuk menghindari persaingan makanan, sehingga mengurangi kepadatan populasi lokal. Akhirnya, kepadatan populasi yang menurun akan memberi umpan balik untuk memengaruhi ekspresi paedomorphosis (Lackey et al., 2019 ; Wakano & Whiteman, 2008 ). Selain contoh-contoh ini, sedikit yang diketahui secara umum tentang bagaimana plastisitas adaptif dalam pertumbuhan individu dan sifat riwayat hidup lainnya mendorong dinamika ekologi populasi (Croll & de Roos, 2022 ; Rundle & Spicer, 2016 ; Spicer & Burggren, 2003 ), yaitu, dinamika eko-fenotipik suatu populasi (Gibert et al., 2022 ). Pertanyaan-pertanyaan penting masih belum terjawab. Misalnya, seberapa besar peran heterokairy dalam dinamika populasi eko-fenotipik? Dan bagaimana dinamika eko-fenotipik memengaruhi dinamika eko-evolusi ?
Di sini, kami bertujuan untuk memberikan titik awal konseptual dan empiris untuk menilai peran yang dapat dimainkan oleh heterokairy dalam dinamika populasi eko-fenotipik. Kami memfokuskan studi kami pada polifenisme (Kotak 1 ), di mana satu genotipe menghasilkan dua atau lebih fenotipe alternatif yang berbeda secara perilaku atau fisik . Kami melakukan ini karena, bagi sebagian besar, kami memiliki pemahaman yang baik tentang peran adaptif mereka (Nettle & Bateson, 2015 ; Nijhout, 2003 ) dan mereka adalah sistem model yang sangat baik untuk memahami evolusi plastisitas (West-Eberhard, 2003 ). Namun, meskipun fenotipe alternatif dalam polifenisme dapat berbeda dalam waktu pengembangan, aspek heterokairic ini kurang dihargai (Rundle & Spicer, 2016 ). Akibatnya, peran heterokairy dalam dinamika eko-fenotipik, sejauh pengetahuan kami, masih belum sepenuhnya terungkap (tetapi lihat: Frankenhuis & Walasek, 2020 ; Walasek et al., 2024 ). Dengan memfokuskan perspektif kami pada polifenisme, kami juga berharap untuk menyoroti penggunaan dan jalur penelitian tambahan untuk sistem model yang sering dipelajari ini.
KOTAK 1. Apa itu polifenisme?
Polifenisme adalah kasus plastisitas perkembangan di mana satu dari serangkaian fenotipe alternatif diskret diproduksi oleh satu genotipe sebagai respons terhadap isyarat lingkungan (West-Eberhard, 2003 ). Isyarat tersebut dapat sangat bervariasi yang mencakup faktor endogen, seperti kelimpahan makanan, isyarat sosial, atau kepadatan populasi dan isyarat eksogen, seperti suhu, fotoperiode, dan risiko predasi. Isyarat ini menyebabkan individu yang sedang berkembang untuk mengekspresikan jalur perkembangan alternatif, yang menghasilkan produksi fenotipe alternatif. Karena polifenisme adalah bentuk plastisitas perkembangan, kami berharap bahwa mereka tidak ‘sepenuhnya’ reversibel, dan fenotipe alternatif akan memiliki efek jangka panjang sepanjang hidup seseorang (Lafuente & Beldade, 2019 ).
Seperti plastisitas secara lebih luas, polifenisme berevolusi ketika tidak ada fenotipe tunggal yang paling menguntungkan. Keadaan seperti itu sering terjadi dari heterogenitas spasial atau temporal di lingkungan, atau ketika ada antagonisme genetik (Galipaud & Kokko, 2020 ). Hal ini dapat menyebabkan fenotipe alternatif diekspresikan dalam lingkungan yang paling cocok untuknya oleh ‘isyarat’, yang merupakan, atau terkait dengan, faktor lingkungan. Misalnya, kupu-kupu biasa Pieris napi menggunakan suhu sebagai ukuran perkiraan musim untuk mengisyaratkan polifenisme warna (Larsdotter Mellström et al., 2010 ), sementara tungau umbi akarid Rhizoglyphus robini menggunakan kondisi tubuh mereka sendiri untuk menilai daya saing mereka (Smallegange et al., 2019 ).
Mungkin, isu yang paling meluas dalam studi plastisitas adalah penekanan pada fenotipe dewasa daripada proses juvenil, yang disebut ‘adult-centrism’ (Rundle & Spicer, 2016 ). Perspektif ini menciptakan keadaan di mana fenotipe dewasa dengan kemampuan bertahan hidup dan reproduksi yang lebih tinggi akan disukai oleh ahli biologi evolusi. Namun, hal itu mengabaikan fakta bahwa kemampuan untuk bertahan hidup pada tahap kehidupan juvenil dan melakukannya dalam kondisi kompetitif sama pentingnya dalam memengaruhi dinamika ekologi seperti fenotipe dewasa itu sendiri, dan karenanya layak mendapat pertimbangan lebih besar dalam literatur ilmiah (Rundle & Spicer, 2016 ).
Bahasa Indonesia: Untuk mencapai tujuan kami, pertama-tama kami menyajikan kerangka kerja tentang bagaimana heterokairy dapat memengaruhi dinamika populasi eko-fenotipik, dan selanjutnya mengeksplorasi bukti empiris apa yang ada untuk mendukungnya. Karena populasi tidak ada dalam isolasi, kami secara singkat menyoroti implikasi dari dinamika eko-fenotipik yang didorong oleh heterokairy pada tingkat meta-populasi dan komunitas. Setelah memberikan sintesis kami, kami membahas implikasi dari kerangka kerja kami di beberapa area utama, terutama: biologi populasi dan sindrom laju kehidupan (POLS). Terakhir, karena ekspresi fenotip alternatif dalam polifenisme dapat berevolusi sebagai respons terhadap kepadatan populasi (Smallegange & Deere, 2014 ; Tomkins et al., 2011 ; Travis et al., 2013 ), kami mengakhiri perspektif kami dengan memikirkan kembali bagaimana seleksi yang bergantung pada kepadatan dapat memengaruhi dinamika populasi eko-evolusi (Smallegange, 2022 ; Travis et al., 2013 ).
2 KERANGKA KERJA UNTUK MEMPELAJARI DINAMIKA POPULASI HETEROKAIRIK DAN EKO-FENOTIPIK
2.1 Bagaimana polifenisme heterokairik dapat mendorong jalur eko-fenotipik
Dinamika populasi seperti perubahan dalam ukuran, struktur, atau pertumbuhan populasi adalah hasil dari perubahan demografi populasi: tingkat kelahiran, perkembangan, dan kematiannya. Jika fenotipe alternatif dalam polifenisme berbeda dalam waktu perkembangan, yaitu fenotipe perkembangannya, kami perkirakan bahwa perubahan dalam ekspresi fenotipenya akan memengaruhi ukuran dan struktur populasi. Misalnya, perubahan fenotipe ke arah perkembangan yang lebih cepat dan pematangan lebih awal akan mendorong reproduksi lebih awal dalam kehidupan, dan tingkat pertumbuhan populasi yang lebih cepat. Kami menyebutnya sebagai jalur fenotipe-ke-ekologi. Perubahan demografis pada gilirannya dapat memengaruhi ekspresi fenotipe alternatif dalam polifenisme heterokairik. Dampak seperti itu pada fenotipe alternatif dapat terjadi jika fenotipe tersebut sensitif terhadap kepadatan populasi (tergantung kepadatan), baik secara langsung, misalnya, sentuhan dan feromon, atau secara tidak langsung melalui, misalnya, persaingan makanan. Kami menyebutnya sebagai jalur ekologi-ke-fenotipe. Jalur eko-fenotipik ini menangkap interaksi antara fenotipe perkembangan individu dan ukuran serta struktur populasi.
Untuk menilai peran apa yang dapat dimainkan heterokairik dalam dinamika eko-fenotipik, pertama-tama kita akan mengeksplorasi jenis fenotip alternatif mana dalam polifenisme yang berbeda dalam waktu perkembangannya. Kemudian kita mengilustrasikan dengan contoh bagaimana polifenisme heterokairik tersebut dapat memengaruhi ukuran dan pertumbuhan populasi. Terakhir, kita akan menggunakan bukti apa pun yang tersedia untuk menunjukkan bagaimana ukuran populasi pada gilirannya dapat memengaruhi ekspresi fenotip alternatif suatu polifenisme, melengkapi lingkaran umpan balik eko-fenotipik (Gibert et al., 2022 ).
2.2 Jalur ekologi-ke-fenotipe dalam polifenisme heterokairik
Heterokairik lazim dalam polifenisme di banyak taksa hewan (Tabel 1 ; Rundle & Spicer, 2016 ), termasuk serangga, nematoda, arakhnida, amfibi, dan krustasea. Perbedaan heterokairik antara fenotipe polifenisme yang berkembang cepat (disebut minor ) dan yang berkembang lambat (disebut mayor ) bisa sangat besar (Tabel 1 ). Misalnya, kutu daun gandum tak bersayap ( Sitobion avenae ) membutuhkan waktu 17 hari untuk dewasa, sedangkan individu bersayap membutuhkan waktu 20 hari untuk dewasa (Xu et al., 2011 ) (Tabel 1 ). Demikian pula, pada salmon Atlantik ( Salmo salar ), individu parr, yang tidak bermigrasi, membutuhkan waktu 1 tahun untuk dewasa, tetapi salmon anadromus, yang bermigrasi ke laut, membutuhkan waktu tambahan hingga 3 tahun (Thorstad et al., 2012 ).
| Kelompok taksonomi a,b , a,b | Jenis | Isyarat | Keterangan | Morf minor (waktu perkembangan) c | Morf utama (waktu pengembangan) c | Ref. |
|---|---|---|---|---|---|---|
| Amfibi | Ambystoma mavortium nebulosum | Kepadatan | Salamander harimau Arizona paedomorfosis | Pedomorf (na) | Metamorf (na) | Lackey dkk. ( 2019 ); Wakano dan Whiteman ( 2008 ) |
| Ambystoma talpoideum | Kepadatan | Salamander mol pedomorfosis | Pedomorf (na) | Metamorf (na) | Ryan dan Semlitsch ( 1998 ) | |
| Ambystoma tigrinum | Kepadatan (makanan) | Morfologi kecebong trofik salamander harimau. | Karnivora (na) | Omnivora (na) | Lannoo dan Bachmann ( 1984 ) | |
| Bunga Rana sylvatica | Predasi | Bentuk ekor katak kayu | Ekor kecil (na) | Ekor besar (na) | Denver ( 2021 ); Maher dkk. ( 2013 ) | |
| Scaphiopus multiplicatus | Kepadatan (makanan) | Morfologi kecebong katak kaki sekop | Karnivora (6 hari) | Omnivora (17 hari) | Pfennig ( 1992 ) | |
| Annelida | Bunga Polydora cornuta | Kepadatan | Poesilogeni | Adelphophage (13–23 hari) | Planktotrof (21 hari) | Mackay dan Gibson ( 1999 ); Beras dan Beras ( 2009 ) |
| Arakhnida | Rhizoglif Robini | Kondisi tubuh | Tungau umbi Acarid bentuk reproduksi jantan | Scrambler (12,6 hari) | Pejuang (14,4 hari) | Smallegange dkk. ( 2019 ); Smallegange dan Coulson ( 2011 ) |
| Kepadatan | Tungau umbi Acarid, deutonymph dispersif. | Pengembangan langsung (na) | Deutonymph (na) | Deere dan kawan-kawan ( 2015 ) | ||
| Rhizoglyphus echinopus | Kepadatan (feromon) | Morfologi reproduksi tungau jantan | Scrambler (tidak) | Pejuang (na) | Smallegange dan kawan-kawan ( 2019 ) | |
| Sancassania berlesei | Kepadatan (feremon) | Morfologi reproduksi tungau jantan | Scrambler (tidak) | Pejuang (na) | Smallegange dan kawan-kawan ( 2019 ) | |
| Crustacea | Daphnia pulex | Predasi | Gigi leher defensif | Tak berdaya (na) | ‘Morph defensif’ (na) | Riessen dan Gilbert ( 2019 ) |
| Jassa mamorata | Kondisi tubuh | Morf reproduksi jantan amphipod | Scrambler (36 hari) | Pejuang (41 hari) | Kurdziel dan Lacey Knowles ( 2002 ) | |
| Ikan | Salmo salar | Kondisi tubuh | Morfologi reproduksi/penyebaran salmon Atlantik jantan | Parr (1 tahun) | Anadromous (4 tahun) | Fleming ( 1996 ); Hutching ( 2011 ); Strom dkk. ( 2018 ); Thomas dkk. ( 1997 ); Thorstad dkk. ( 2012 ) |
| Serangga | Forficula auricularia | Kondisi tubuh | Earwig Eropa jantan reproduksi morf | Scrambler (tidak) | Pejuang (na) | Tomkins dan kawan-kawan ( 1999 ); Tomkins dan kawan-kawan ( 2005 ) |
| Onthophagus akuminatus | Kondisi tubuh | Kumbang kotoran jantan bentuk reproduksi | Scrambler (tidak) | Pejuang (na) | Emlen ( 1994 ) | |
| Onthophagus taurus | Kondisi tubuh | Kumbang kotoran bertanduk banteng bentuk reproduksi jantan | Scrambler (tidak) | Pejuang (na) | Emlen dan Nijhout ( 2001 ); Tomkins dkk. ( 2005 ) | |
| Schistocerca gregaria | Kepadatan (sentuhan) | Kebersamaan belalang | Suka berteman (30 hari) | Soliter (33,6 hari) | Ayali ( 2019 ); Cheke ( 1978 ); Maeno dkk. ( 2020 ); Maeno dan Tanaka ( 2008 ); Rogers dkk. ( 2003 ) | |
| Sitobion avenae | Kepadatan (sentuhan) | Kutu daun volancy | Non-volant (17 hari) | Volant (20 hari) | Anggap dkk. ( 2024 ); Xu dkk. ( 2011 ) | |
| Nematoda | Pristionchus pacificus | Kepadatan (makanan) | Tahapan nematoda dauer | Pengembangan langsung (na) | Dauer (na) | Fielenbach dan Antebi ( 2008 ); Renahan dkk. ( 2021 ) |
| Kepadatan (makanan) | Morfologi trofik nematoda | Omnivora (na) | Bakteriovora (na) | Casasa dkk. ( 2021 ); Dardiry dkk. ( 2023 ) |
Catatan : Polifenisme yang mewakili berbagai kelompok taksonomi dan spesies yang fenotipe alternatifnya berbeda dalam laju perkembangannya. Tercantum juga isyarat yang memicu perkembangan fenotipe alternatif dan istilah yang digunakan untuk menunjukkan fenotipe yang berkembang lebih cepat dan lebih lambat. Dalam tanda kurung di bawah ini, setiap morf minor/mayor adalah waktu perkembangannya masing-masing; perhatikan bahwa waktu-waktu ini dapat memiliki satuan yang berbeda.
a Kami telah mengecualikan ‘polifenisme musiman’, seperti yang umum terlihat pada kupu-kupu (Larsdotter Mellström et al., 2010 ) dan kutu daun (Frantz et al., 2006 ; Nespolo et al., 2009 ) karena mereka tidak mengandung jalur fenoekologi karena fenotipe tidak dapat mengubah musim.
b Yang juga dikecualikan adalah polifenisme serangga eusosial, seperti semut dan lebah, karena mereka memiliki perilaku sosial yang kuat yang membentuk kembali pertukaran umum (Negroni et al., 2016 ), yang mempersulit perbandingan dengan spesies non-eusosial.
c Jika waktu pengembangan tidak dinyatakan secara eksplisit dalam makalah yang dikutip, kami mengganti waktu dengan ‘na’.
2.3 Jalur fenotipe-ke-ekologi dalam polifenisme heterokairik
Jika fenotipe alternatif memiliki tingkat perkembangan yang berbeda, maka efeknya pada pertumbuhan populasi juga akan berbeda (Smallegange, 2022 ), tetapi sedikit bukti empiris untuk ini (Rundle & Spicer, 2016 ; tetapi lihat misalnya di S. avenae , Xu et al., 2011 , dan Rhopalosiphum padi , Nespolo et al., 2009 ). Dari beberapa spesies yang tercantum dalam Tabel 1 , kita tahu bahwa plastisitas dalam perkembangan dapat menyebabkan perubahan pada populasi. Pertama, jantan tungau umbi akarid Rhizoglyphus robini adalah ‘pejuang’ bersenjata dengan kaki berotot tebal yang memiliki ujung tajam, yang dapat mereka gunakan untuk membunuh yang lain, atau ‘pengacak’ tanpa senjata, yang tidak memiliki kaki seperti itu (Smallegange et al., 2019 ). Karena perkembangan kaki petarung mereka, petarung tumbuh lebih lambat daripada pengacak, jika hal-hal lain sama (14,4 dan 12,6 hari rata-rata untuk petarung dan pengacak yang memakan akses ad lib ke ragi, masing-masing; Smallegange, 2011 ). Oleh karena itu, tidak mengherankan bahwa variasi dalam kelimpahan relatif petarung dan pengacak memengaruhi ukuran populasi (Deere & Smallegange, 2023 ). Kedua, belalang gurun ( Schistocerca gregaria ) berkembang baik sebagai individu soliter yang tumbuh lambat, atau sebagai individu berkelompok yang berkembang cepat (Ayali, 2019 ; Cheke, 1978 ; Rogers et al., 2003 ). Peningkatan ekspresi belalang yang berkembang cepat dapat secara substansial meningkatkan pertumbuhan dan ukuran populasi, dan dalam kasus yang paling ekstrem, menciptakan wabah (Cheke, 1978 ). Akhirnya, pada kepadatan rendah salamander mol ( Ambystoma talpoideum ) dapat menjadi pedomorfosis untuk memperpendek waktu pematangan. Sementara itu pada kepadatan tinggi, proporsi individu A. talpoideum yang lebih besar memilih untuk bermetamorfosis untuk menghindari persaingan dan memperpanjang perkembangan. Semakin banyak individu bermetamorfosis, semakin rendah laju pertumbuhan populasi lokal, tidak hanya karena metamorf ini berkembang lebih lambat (Ryan & Semlitsch, 1998 ), tetapi juga karena mereka menyebar, menurunkan ukuran populasi natal. Secara keseluruhan, ada beberapa bukti yang menunjukkan bahwa perbedaan waktu perkembangan antara fenotipe alternatif dapat memiliki pengaruh yang signifikan terhadap pertumbuhan, ukuran, dan struktur populasi, seperti yang kami duga.
2.4 Umpan balik eko-fenotipik dalam polifenisme heterokairik
2.4.1 Umpan balik yang diamati
Dalam umpan balik eko-fenotipik (Gibert et al., 2022 ), variabel ekologi yang memengaruhi ekspresi fenotipe alternatif pada gilirannya dapat dipengaruhi oleh ekspresi fenotipe alternatif. Sejauh pengetahuan kami, setidaknya ada tiga sistem tempat umpan balik eko-fenotipik tersebut telah diamati. Pertama, pengacak R. robini menonjol pada kepadatan tinggi, tempat keterbatasan sumber daya mendorong individu untuk dewasa lebih awal dan terhindar dari kematian remaja (Rhebergen, 2022 ). Penurunan waktu perkembangan seperti itu dapat mempercepat pertumbuhan populasi karena individu dapat bereproduksi lebih awal dalam hidup. Hal ini, pada gilirannya, semakin membatasi ketersediaan sumber daya dan semakin meningkatkan kecenderungan individu untuk berkembang menjadi pengacak. Kedua, pada nematoda P. pacificus , Renahan et al. ( 2021 ) mengamati siklus dalam ekspresi tahap dauer . Dalam umpan balik ini, populasi besar mengonsumsi sumber daya hingga habis, mendorong juvenil memasuki tahap dauer, menghambat pertumbuhan populasi hingga titik penurunan populasi. Ketika populasi menurun dan sumber daya terisi kembali, dauer berkembang menjadi dewasa reproduktif, meningkatkan ukuran populasi dan memicu dimulainya siklus baru (Renahan et al., 2021 ). Akhirnya, volancy pada kutu daun kacang polong (Deem et al., 2024 ; Xu et al., 2011 ) menghasilkan lingkaran umpan balik seperti pada P. pacificus dauer, di mana morf yang berkembang cepat memanfaatkan makanan yang melimpah pada kepadatan rendah, menyebabkan lonjakan pertumbuhan dan ukuran populasi. Peningkatan ukuran populasi ini menghabiskan makanan dan meningkatkan persaingan, mendorong beberapa individu untuk berkembang perlahan sebagai penyebar, yang pada gilirannya menyebabkan penurunan laju pertumbuhan populasi (Deem et al., 2024 ; Xu et al., 2011 ).
2.4.2 Umpan balik yang diharapkan
Fakta bahwa kami hanya menemukan bukti langsung umpan balik eko-fenotipik dalam tiga sistem mendorong pertanyaan tentang sistem lain apa yang dapat kita harapkan umpan balik eko-fenotipik terjadi? Banyak polifenisme, di mana fenotip alternatif berbeda dalam tingkat perkembangan dan reproduksi, bergantung pada kondisi (misalnya tungau umbi R. robini , Smallegange et al., 2019 ; kumbang kotoran Onthophagus acuminatus , Emlen, 1994 ; earwig Eropa Forficula auricularia Tomkins et al., 2005 , dan amphipoda Jassa marmorata , Kurdziel & Lacey Knowles, 2002 ). Dalam sistem seperti itu, individu dalam kondisi fisik yang baik, yaitu individu utama, mengekspresikan morfologi mahal (kaki petarung, tanduk, cerci) tetapi individu dalam kondisi fisik yang buruk, minor, tidak. Kepadatan tinggi dapat menyebabkan peningkatan persaingan atas makanan. Sebagai hasil dari kompetisi ini, proporsi individu yang lebih besar akan memiliki kondisi fisik yang buruk, yang menyebabkan peningkatan kecenderungan untuk berkembang sebagai anak di bawah umur, karena remaja tidak dapat berinvestasi dalam morfologi yang mahal. Sebaliknya, ketersediaan makanan yang tinggi dan kompetisi yang rendah meningkatkan kelimpahan (relatif) dari individu dengan kondisi fisik yang baik. Ketika perbedaan dalam kondisi tubuh karena fluktuasi kepadatan populasi dan kompetisi makanan menyebabkan perbedaan ekspresi individu dengan kondisi fisik yang baik, kita akan mengharapkan dinamika eko-fenotipik terjadi, seperti pada R. robini (Deere & Smallegange, 2023 ).
Menariknya, pada ikan trout dan salmon, juvenil dalam kondisi fisik yang baik berkembang menjadi minor sementara juvenil dalam kondisi fisik yang buruk berkembang menjadi mayor. Misalnya, pada salmon Coho Oncorhynchus kisutch , jantan dengan kondisi tubuh yang baik menahan diri untuk tidak mengembangkan persenjataan (rahang ‘berkait’, atau hidung) dan lebih cepat dewasa dalam hidup sebagai ‘jack’ untuk mengadopsi strategi sneaker untuk mendapatkan akses ke betina. Jantan dengan kondisi tubuh yang buruk, sebaliknya, bermigrasi dan dewasa pada usia yang jauh lebih tua dalam hidup sebagai pejuang anadrom (‘hooknosis’). Hooknose seperti itu memonopoli akses ke betina setelah mereka bermigrasi kembali ke tempat perkembangbiakan mereka (Koseki & Fleming, 2006 ). Dalam sistem ini, pematangan awal dan reproduksi dalam hidup jack dapat memicu pertumbuhan populasi lokal. Pertumbuhan populasi lokal ini kemudian dapat, pada gilirannya, membatasi ketersediaan sumber daya karena persaingan di antara rekrutan, yang berpotensi menyebabkan lebih banyak rekrutan berkembang menjadi hooknose. Jika semakin banyak jantan berkembang perlahan hingga dewasa sebagai hooknose, jika hal lain sama, laju pertumbuhan populasi akan menurun. Pada salmonid, migrasi anadrom ini dapat memiliki laju kematian yang tinggi (misalnya laju kematian 8%–71% pada anadrom S. salar , Thorstad et al., 2012 ) yang dapat memengaruhi dinamika eko-fenotipik dengan mengurangi ukuran populasi. Oleh karena itu, mekanisme pasti dari dinamika eko-fenotipik khusus ini masih harus diurai. Namun, kami perkirakan bahwa pergerakan migrasi O. kisutch dan salmonid lainnya akan mendorong umpan balik eko-fenotipik, karena kembalinya individu yang bermigrasi ke suatu populasi akan memengaruhi kepadatannya dan dengan demikian isyarat yang mendorong migrasi untuk memulai.
3 RINGKASAN POLA UMUM NAMUN BERBEDA
3.1 Polifenisme heterokairik pada hewan ektotermik versus endotermik
Menariknya, penelusuran kami hanya mengungkap polifenisme heterokairik pada ektoterm (Tabel 1 ). Mungkin, kami tidak dapat menemukan bukti empiris, atau sebagai alternatif, polifenisme heterokairik tidak ada pada endoterm. Bisa jadi juga polifenisme endoterm terutama bersifat perilaku dan dengan demikian lebih sulit dibedakan. Di sisi lain, bagaimanapun, mungkin ada alasan fungsional mengapa polifenisme heterokairik dominan, atau bahkan secara eksklusif, terjadi pada ektoterm. Misalnya, ektoterm memiliki pertumbuhan ‘didorong oleh pasokan’, di mana pertumbuhan individu, di antara proses perkembangan lainnya, bervariasi dengan pasokan makanan dan sangat fleksibel (Kooijman, 2010 ). Endoterm, di sisi lain, jauh kurang fleksibel dan menunjukkan pertumbuhan ‘didorong oleh permintaan’, di mana mereka membutuhkan pasokan makanan dalam jumlah minimal untuk bertahan hidup dan tumbuh. Terkait dengan pertumbuhan yang didorong oleh pasokan, ektoterm memiliki kemampuan yang lebih buruk untuk mempertahankan homeostasis daripada endoterm (Clarke, 2006 ), yang berpotensi memicu evolusi polifenisme heterokairik, khususnya di lingkungan yang bervariasi. Terlebih lagi, endoterm memiliki mekanisme fisiologis untuk mengatasi stres termal, seperti berkeringat, terengah-engah, atau menggigil (Huey et al., 2002 ). Oleh karena itu, polifenisme heterokairik mungkin lebih mungkin berevolusi pada ektoterm untuk mencocokkan peristiwa siklus hidup mereka dengan suhu optimal untuk pertumbuhan dan reproduksi. Endoterm mungkin kurang membutuhkan plastisitas seperti itu, karena mereka dapat menahan efek fluktuasi suhu pada perkembangan mereka (Reece, 2011 ). Bisa jadi juga karena alasan-alasan ini, polifenisme endoterm terutama bersifat perilaku dan dengan demikian lebih sulit dibedakan. Dengan demikian, kami mengusulkan bahwa umpan balik eko-fenotipik lebih lazim terjadi pada ektoterm daripada pada endoterm.
3.2 Sistem Minor pada Kepadatan Rendah dan Minor pada Kepadatan Tinggi
Jika plastisitas disebabkan oleh faktor endogen, umpan balik eko-fenotipik kemungkinan tersebar luas dan heterokairy dapat memainkan peran penting di dalamnya (Gambar 1 ). Efek pasti heterokairy dalam membentuk umpan balik eko-fenotipik akan bergantung pada apakah fenotip minor paling sering diproduksi pada kepadatan rendah dan fenotip mayor pada kepadatan tinggi (Gambar 1a ) atau sebaliknya (Gambar 1b ). Sistem minor-pada-kepadatan-rendah meliputi paedomorfosis salamander (Ryan & Semlitsch, 1998 ), volansi kutu daun (Xu et al., 2011 ), pembentukan nematoda dauer (Renahan et al., 2021 ) dan migrasi salmon (Hutchings, 2011 ). Tetapi banyak polifenisme adalah sistem minor-pada-kepadatan-tinggi, di mana minor sebagian besar terjadi pada kepadatan populasi tinggi ketika persaingan makanan tinggi, dan fenotipe mayor yang matang terlambat pada kepadatan rendah ketika persaingan rendah. Contohnya termasuk polifenisme reproduksi invertebrata (Emlen & Nijhout, 2001 ; Kurdziel & Lacey Knowles, 2002 ; Smallegange et al., 2019 ; Tomkins et al., 2005 ), polifenisme trofik (Dardiry et al., 2023 ; Lannoo & Bachmann, 1984 ; Pfennig, 1992 ), dan gregariousness belalang (Ayali, 2019 ; Cheke, 1978 ; Rogers et al., 2003 ).
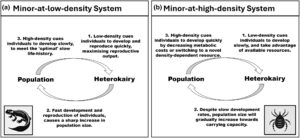
Kami berharap sistem minor-pada-kepadatan-rendah dan minor-pada-kepadatan-tinggi untuk keduanya menjadi penyangga individu terhadap variasi lingkungan, tetapi dengan cara yang berbeda. Secara khusus, sistem minor-pada-kepadatan-rendah mencerminkan prediksi teori riwayat hidup klasik bahwa dalam populasi yang berkembang (yaitu kepadatan populasi rendah), waktu perkembangan harus diminimalkan (Stearns, 1976 ). Dengan demikian, kami akan berharap lebih banyak minor terjadi pada kepadatan rendah, tetapi relatif lebih banyak mayor pada kepadatan tinggi. Salamander, sekali lagi, memberikan contoh buku teks karena mengoptimalkan penggunaan badan air sementara dengan mempercepat pematangan pada kepadatan rendah, tetapi menundanya pada kepadatan tinggi (Ryan & Semlitsch, 1998 ). Sistem minor-pada-kepadatan-tinggi juga menjadi penyangga terhadap kondisi lingkungan yang bervariasi dengan memungkinkan juvenil yang tertekan sumber daya untuk tetap matang tetapi sebagai minor. Hal ini dilakukan dengan mengurangi biaya pematangan dan metabolisme (misalnya R. robini , Croll et al., 2019 ), atau mengakses sumber daya yang bergantung pada kepadatan (misalnya S. multiplicatus , Pfennig, 1992 ). Dengan mengurangi biaya metabolisme atau mengakses sumber daya alternatif, individu dengan fenotipe alternatif mengurangi paparan mereka terhadap persaingan intraspesifik untuk sumber daya, yang disebut pelepasan kompetitif (Chapman et al., 2011 ; Figueredo et al., 2009 ). Namun, sementara sistem minor-pada-kepadatan-rendah dan minor-pada-kepadatan-tinggi dapat melindungi individu terhadap perubahan lingkungan dan mengoptimalkan kebugaran individu, efeknya pada populasi mungkin berbeda secara signifikan tergantung pada jenis umpan balik yang mereka hasilkan (DeAngelis et al., 2012 ). Sebagai kesimpulan, proses alternatif dan mungkin tersebar luas ada di mana organisme dapat melindungi diri terhadap perubahan lingkungan, yang masing-masing dapat memacu dinamika eko-fenotipik.
3.3 Sistem kepadatan fenotipe yang berbeda terkait dengan polifenisme yang berbeda
Dalam sistem minor-pada-kepadatan-rendah dan minor-pada-kepadatan-tinggi, terdapat pengelompokan signifikan ‘tipe’ polifenik dalam setiap sistem. Secara khusus, dalam sistem minor-pada-kepadatan-rendah, kita melihat polifenisme yang dominan dispersif (misalnya tungau umbi acarid deutonymph, Edwards, pengamatan pribadi; nematoda dauer, Renahan et al., 2021 ; migrasi salmonid, Koseki & Fleming, 2006 ) dan defensif (misalnya morfologi ekor kecebong, Denver, 2021 ; Maher et al., 2013 ; gigi leher daphnia, Riessen & Gilbert, 2019 ; lihat Bagian 3.4 ). Di sisi lain, sistem minor-pada-kepadatan-tinggi sebagian besar bersifat trofik (misalnya kodok kaki sekop, nematoda, belalang), atau reproduktif (tungau umbi, kumbang kotoran, amphipoda). Tetapi proses apa yang akan mendorong pengelompokan tipe polifenik tersebut dalam sistem kepadatan fenotipe yang berbeda? Satu penjelasannya adalah bahwa dalam populasi tempat mayor diekspresikan pada kepadatan rendah, tekanan seleksi selain yang mendukung waktu perkembangan cepat sedang berperan. Contohnya adalah persaingan jantan-jantan dewasa seperti yang terlihat pada tungau dan kumbang kotoran di atas. Atau, seleksi dapat mendukung waktu perkembangan yang lebih pendek untuk meningkatkan keberhasilan predasi, misalnya, dalam polifenisme trofik seperti pada contoh nematoda dan kodok kaki sekop di atas. Keberagaman belalang adalah kasus menarik di mana, meskipun spesies tersebut menunjukkan polifenisme dispersif, itu adalah sistem minor-pada-kepadatan tinggi dan bukan yang minor-pada-kepadatan rendah. Mungkin, perkembangan cepat disukai karena individu yang suka berkelompok memakan juvenil, memilih perkembangan cepat untuk meminimalkan risiko kanibalisme (Hansen et al., 2011 ). Pekerjaan di masa depan dapat mengungkap bagaimana tekanan seleksi alternatif dapat mengganggu pola riwayat kehidupan yang biasanya kita harapkan untuk diamati pada kepadatan tinggi versus kepadatan rendah.
3.4 Di luar tingkat populasi
Jika dinamika eko-fenotipik terjadi pada tingkat populasi, dinamika tersebut kemungkinan besar juga terjadi pada tingkat meta-populasi dan komunitas. Dalam meta-populasi, polifenisme dispersif dapat menciptakan umpan balik eko-fenotipik negatif dan positif dari dispersi (Tabel 1 , misalnya P. pacificus dauer, Renahan et al., 2021 ; S. avenae volancy, Xu et al., 2011 ; A talpoidium metamorphosis, Ryan & Semlitsch, 1998 ; annelid poecilogeny, Mackay & Gibson, 1999 ; R. robini deutonymph, Deere et al., 2015 ) dan migrasi parsial (misalnya O. kisutch , Koseki & Fleming, 2006 ; Salmo salar , Hutchings, 2011 ). Perbedaan utama antara polifenisme dispersif dan parsial-migran adalah bahwa, sementara penyebar meninggalkan lingkungan mereka dan menciptakan keturunan non-dispersif, migran akan kembali ke lingkungan kelahiran mereka untuk bereproduksi. Kembalinya untuk berkembang biak ini meningkatkan kepadatan dan membuat migran lebih mungkin menghasilkan keturunan dispersif. Dengan demikian, polifenisme dispersif dan parsial-migran ini masing-masing menciptakan umpan balik eko-fenotipik negatif dan positif.
Demikian pula, pada tingkat komunitas, polifenisme defensif dapat menciptakan umpan balik negatif dengan mengurangi tingkat predasi dan mendorong pergantian mangsa (Kishida et al., 2009 ; Prokopenko et al., 2023 ; Riessen & Gilbert, 2019 ; Tollrian, 1995 ). Pemberian makan alternatif oleh predator kemudian dapat mengurangi ekspresi morfologi yang diinduksi. Umpan balik negatif tersebut untuk morfologi defensif bahkan mungkin telah memilih untuk reversibilitas fitur plastik ini (Gabriel, 1999 ; Orizaola et al., 2012 ). Ketika mengembalikan fokus kita ke heterokairy sebagai agen utama dalam dinamika eko-fenotipik, ekspresi morfologi defensif mengurangi tingkat pertumbuhan dan pematangan, berdampak negatif pada ukuran dan pertumbuhan populasi (Maher et al., 2013 ; Riessen & Gilbert, 2019 ). Oleh karena itu, masuk akal jika pengurangan ukuran populasi bersamaan dengan ciri-ciri morfologi itu sendiri mengisyaratkan predator untuk menggunakan petak alternatif (Vijayan et al., 2019 ) atau menargetkan mangsa alternatif (Kishida et al., 2009 ; Prokopenko et al., 2023 ).
3.5 Arah masa depan
Dalam perspektif ini, kami telah mengeksplorasi peran potensial yang dimainkan oleh heterokairy dalam dinamika populasi eko-fenotipik. Secara khusus, kami telah menyediakan kerangka kerja konseptual yang menyoroti bagaimana polifenisme heterokairik dapat mendorong dinamika populasi eko-fenotipik dalam sistem minor-pada-kepadatan-rendah atau minor-pada-kepadatan-tinggi. Kami juga menyoroti bagaimana masing-masing sistem ini dapat menahan lingkungan yang penuh tekanan dengan cara yang berbeda. Sayangnya, rentang taksa yang kami bahas kecil, sebagian karena kurangnya studi empiris tentang konsekuensi riwayat hidup polifenisme. Menerapkan kerangka kerja kami pada bentuk-bentuk plastisitas yang berkelanjutan dapat meningkatkan keragaman taksa, termasuk yang tidak terwakili. Mungkin, dengan demikian dimungkinkan untuk menjawab pertanyaan-pertanyaan penting, termasuk: Apakah sistem eko-fenotipik yang berbeda memicu dinamika populasi yang berbeda? Bagaimana dinamika eko-fenotipik memengaruhi seleksi yang bergantung pada kepadatan? Apakah ada ruang untuk ketidaksesuaian antara isyarat dan tekanan seleksi pada fenotipe plastik?
3.5.1 Apakah sistem eko-fenotipik yang berbeda memicu dinamika populasi yang berbeda?
Meskipun keduanya berkontribusi terhadap ketahanan individu terhadap perubahan lingkungan, sistem minor-pada-kepadatan-rendah dan minor-pada-kepadatan-tinggi beroperasi melalui mekanisme yang berbeda. Apakah mekanisme ini eksplisit atau implisit, perbedaan tersebut dapat berjenjang untuk memiliki konsekuensi populasi yang lebih besar. Sistem minor-pada-kepadatan-rendah dan minor-pada-kepadatan-tinggi keduanya dapat menghasilkan lingkaran umpan balik positif atau negatif, dan dengan demikian dapat menjadi tidak stabil atau menstabilkan dalam hal persistensi populasi. Misalnya, polifenisme trofik dapat menstabilkan populasi dengan mendiversifikasi penggunaan sumber daya dan mempromosikan populasi campuran ukuran dan usia (Claessen et al., 2000 ) atau mengacaukannya dengan memakan juvenil (Buonomo & Lacitignola, 2006 ). Selain itu, polifenisme reproduksi yang bergantung pada kondisi dapat mengakibatkan pergeseran rasio jenis kelamin, yang berpotensi memiliki konsekuensi populasi yang tidak stabil secara signifikan (Lindström & Kokko, 2002 ). Dalam sistem minor-pada-kepadatan-rendah, kemampuan untuk beralih ke laju perkembangan optimal untuk kepadatan tinggi dan rendah kemungkinan akan menstabilkan populasi dengan mengurangi seleksi yang bergantung pada kepadatan. Polifenisme yang melibatkan fenotipe penyebaran juga dapat meningkatkan stabilitas populasi dalam banyak kasus (Vance, 1984 ), kecuali dalam kasus-kasus ketika ia menghambat pertumbuhan atau mendorong pertumbuhan populasi negatif. Perbedaan antara sistem minor-pada-kepadatan-rendah dan minor-pada-kepadatan-tinggi juga dapat muncul karena perbedaan sistematis dalam penggunaan isyarat; misalnya, jika individu merespons secara langsung atau untuk mengantisipasi stresor (Frankenhuis & Walasek, 2020 ; Smallegange, 2022 ). Mengembangkan fenotipe untuk mengantisipasi stresor dapat meningkatkan ruang lingkup ketidaksesuaian lingkungan. Hal ini terutama terjadi jika isyarat dan stresor menjadi tidak sinkron, yang mengarah pada plastisitas nonadaptif atau maladaptif (Nettle & Bateson, 2015 ). Meskipun beberapa pihak telah mengusulkan bahwa plastisitas nonadaptif tersebut dapat memberikan kontribusi positif terhadap evolusi sifat adaptif (Ghalambor et al., 2007 , 2015 ), efek tersebut dapat memiliki dampak negatif yang signifikan terhadap pertumbuhan dan stabilitas populasi. Pekerjaan di masa mendatang dapat berupaya untuk memperluas kerangka kerja kami dengan menyelidiki perbedaan mekanistik antara sistem eko-fenotipik, karena perbedaan sistematis tersebut dapat berdampak pada tingkat populasi atau lebih tinggi.
3.5.2 Dapatkah sistem eko-fenotipik yang berbeda menjelaskan mengapa kita melihat berbagai respons laju kehidupan terhadap variabilitas lingkungan?
Polifenisme analog dengan sindrom multi-sifat (Smallegange & Guenther, 2024 ), dan kesimpulan kami tentang polifenisme heterokairik paling mirip dengan POLS (Wright et al., 2019 ). Karena teori POLS saat ini dibangun atas trade-off antara reproduksi saat ini dan masa depan (Wright et al., 2019 ), teori ini berfokus pada hasil perkembangan, yaitu fenotipe dewasa. Namun, premis kami berpusat pada bagaimana proses perkembangan membangun fenotipe selama ontogeni. Mendamaikan ini melalui perspektif perkembangan yang lebih berarti, misalnya, memperkenalkan trade-off yang dihadapi selama perkembangan. Selama perkembangan, organisme dapat berbeda dalam perolehan energinya karena kondisi lingkungan, termasuk kepadatan populasi, berfluktuasi. Tetapi individu juga dapat berbeda dalam cara mereka mengalokasikan energi antara pemeliharaan dan pertumbuhan versus pematangan dan reproduksi. Faktanya, studi terbaru menunjukkan bahwa heterogenitas dalam perolehan versus alokasi energi bertanggung jawab atas dukungan empiris campuran dari hipotesis POLS (Haave-Audet et al., 2022 ; Prabh et al., 2023 ). Dengan demikian, kita harus mempertimbangkan keseimbangan relatif variasi dalam alokasi energi dibandingkan dengan perolehan energi antara individu dan memasukkan dampak yang dapat dimiliki individu terhadap diri mereka sendiri. Misalnya, pemberian makan akan memengaruhi perolehan energi individu tetapi secara berbeda pada kepadatan populasi yang berbeda (Gambar 1 ). Hal ini pada gilirannya akan memengaruhi lintasan perkembangan individu dan ekspresi fenotipe. Kerangka konseptual kami tentang sistem minor-pada-kepadatan-rendah dan minor-pada-kepadatan-tinggi dapat diterapkan pada teori POLS tersebut untuk memberikan latar belakang teoritis mengapa kita mengamati keragaman respons laju kehidupan terhadap tekanan tertentu, seperti kepadatan. Aplikasi semacam itu akan menjadikan perbedaan dalam respons POLS, baik terhadap kecepatan hidup yang cepat maupun lambat, sebagai bagian integral dari teori, dan menghubungkan perbedaan tersebut dalam respons terhadap lingkungan yang memicunya (Hämäläinen et al., 2021 ; Smallegange & Guenther, 2024 ). Dengan demikian, perspektif perkembangan eko-fenotipik dapat memperkenalkan dasar mekanistik pada proses (alih-alih hasil) perkembangan fenotipe. Hal ini dapat membantu mengubah kerangka kerja POLS modern menjadi kerangka kerja yang lebih prediktif tentang bagaimana riwayat hidup, ciri fisiologis dan perilaku bervariasi secara konsisten di seluruh populasi dan spesies (Smallegange & Guenther, 2024)). Perspektif ini akan mencakup dinamika eko-fenotipik karena menggabungkan umpan balik populasi melalui kompetisi. Namun, perspektif ini juga menggabungkan bagaimana fluktuasi lingkungan yang kontras dan umpan balik populasi menciptakan isyarat populasi yang berubah pada ekspresi fenotip pada skala waktu kontemporer. Perubahan isyarat tersebut dapat meningkatkan keragaman POLS dalam suatu lingkungan, dan dapat memacu dinamika populasi eko-fenotipik lebih lanjut.
3.5.3 Bagaimana dinamika eko-fenotipik memengaruhi seleksi yang bergantung pada kepadatan?
Jika fenotipe alternatif dalam polifenisme heterokairik sensitif secara diferensial terhadap kepadatan populasi (baik secara langsung melalui misalnya sentuhan/feromon atau secara tidak langsung melalui kompetisi makanan; Tabel 1 ), maka periode kepadatan tinggi atau kepadatan rendah yang berkepanjangan dapat memaksakan seleksi pada ekspresinya (Deere & Smallegange, 2023 ; Smallegange & Deere, 2014 ; Tomkins et al., 2011 ; Travis et al., 2013 ). Secara khusus, seleksi yang dipaksakan oleh kepadatan populasi tinggi atau rendah yang berkepanjangan dapat mengembangkan ambang isyarat (di atas dan di bawahnya individu berkembang menjadi fenotipe minor atau mayor). Evolusi ambang ini akan memfasilitasi lebih banyak individu untuk berkembang menjadi fenotipe berkinerja terbaik pada kepadatan itu (Tomkins et al., 2011 ). Untuk memprediksi ke arah mana ambang isyarat berevolusi, misalnya, menuju peningkatan ekspresi tipe mayor atau minor, perlu diketahui kebugaran fenotipe alternatif di bawah kepadatan populasi yang berbeda (Deere & Smallegange, 2023 ; Nettle & Bateson, 2015). Dalam contoh yang kita bahas, kita menemukan bahwa pada beberapa spesies, kepadatan populasi yang tinggi mendukung mayor (misalnya salamander mol, Ryan & Semlitsch, 1998), tetapi untuk spesies lain, mendukung minor (misalnya tungau umbi; Smallegange et al., 2019 ). Dengan asumsi individu polifenik akan menghasilkan fenotipe yang mengembalikan kebugaran tertinggi di lingkungan mereka saat ini (Chevin et al., 2013 ; Ghalambor et al., 2007 ), seleksi bergantung kepadatan yang terus-menerus akan mendukung evolusi ambang isyarat menuju ekspresi fenotipe yang paling optimal (Ghalambor et al., 2007 ). Dalam kasus yang paling ekstrim, akomodasi genetik akan memperbaiki fenotipe yang menguntungkan secara genetik, dan plastisitas akan hilang (Lafuente & Beldade, 2019 ).
Apakah dinamika eko-fenotipik dapat memfasilitasi seleksi yang bergantung pada kepadatan, akan bergantung pada apakah respons ambang isyarat itu sendiri bersifat plastis, yaitu, apakah respons tersebut menunjukkan plastisitas sebagai respons terhadap kondisi lingkungan lokal, seperti variasi makanan (Emlen, 1994 ; Moczek & Emlen, 2000 ). Misalnya, variasi makanan tidak hanya dapat menyebabkan variasi kondisi tubuh, dan karenanya dalam morfologi (mayor/minor), tetapi juga dalam cara organisme peka terhadap isyarat kondisi untuk mengembangkan sifat morfologi ini. Jika arah plastisitas alometrik yang bergantung pada makanan tersebut bertepatan dengan pergeseran evolusi dalam alometri, hal ini dapat memfasilitasi adaptasi lokal yang cepat, divergensi adaptif, dan bahkan spesiasi (Rhebergen, 2022 ). Hal ini mendorong pertanyaan: apa yang akan terjadi jika ekspresi fenotipe yang bergantung pada kepadatan terjadi secara diametris berlawanan dengan plastisitas alometrik? Misalnya, jantan dalam populasi R. robini yang dipelihara pada sumber makanan yang sangat buruk (kertas saring), awalnya hanya mengekspresikan pengacak. Namun, akhirnya, meskipun kondisi gizi buruk, setelah puluhan generasi, jantan mengembangkan kapasitas untuk berkembang menjadi petarung, meskipun dalam ukuran yang sangat kecil (Smallegange, pengamatan pribadi). Pengamatan ini menunjukkan bahwa ada seleksi kuat untuk ekspresi petarung dalam spesies ini, tetapi plastisitas yang bergantung pada gizi dalam ekspresi morf jantan dapat menangkal seleksi ini, setidaknya pada awalnya, dalam kondisi makanan yang buruk. Apakah plastisitas alometrik menyediakan ‘waktu pembelian’ untuk memfasilitasi efisiensi perkembangan yang lebih besar (Efek Baldwin; Badyaev, 2009 ) atau bahwa plastisitas non-adaptif memacu evolusi (Ghalambor et al., 2015 ) dalam konteks seperti itu, memerlukan penelitian lebih lanjut.
Penyebaran dapat bergantung pada kepadatan (Tabel 1 ), dan sifat-sifat penyebaran umumnya berevolusi di bawah heterogenitas spasial atau temporal dalam kondisi lingkungan (Buoro & Carlson, 2014 ). Ekspresi penyebaran umumnya tinggi ketika kepadatan populasi lokal tinggi, dan individu menyebar ke lokasi yang lebih baik dan berdensitas rendah (Buoro & Carlson, 2014 ). Namun, jika kondisi menjadi lebih konstan, misalnya, jika kepadatan tetap tinggi di seluruh metapopulasi, kami perkirakan kebugaran individu yang dispersif menurun, karena mereka menjadi maladaptasi karena ketidaksesuaian lingkungan (Simmons & Thomas, 2004 ). Ekspresi fenotipe yang tidak menyebar akan lebih disukai (Simmons & Thomas, 2004 ), yang memicu evolusi ambang isyarat penyebaran menuju penurunan ekspresi penyebaran. Meskipun skenario ini mungkin tidak begitu mungkin karena populasi alami hampir selalu mengalami variasi dalam kondisi lingkungan, setiap evolusi ambang batas isyarat penyebaran akan memengaruhi dinamika eko-evolusi (meta-)populasi (Deere & Smallegange, 2023 ).
4 KESIMPULAN
Premis kami adalah bahwa heterokairy dapat memainkan peran penting dalam dinamika eko-fenotipik, setidaknya pada ektoterm. Namun, cara heterokairy mendorong dinamika eko-fenotipik akan bergantung pada bagaimana perkembangan berbeda antara fenotipe alternatif dalam polifenisme (misalnya minor vs. mayor) dan bagaimana ekspresinya bervariasi dengan kepadatan populasi (misalnya minor-pada-kepadatan-rendah atau minor-pada-kepadatan-tinggi). Dinamika eko-fenotipik cenderung mendorong seleksi yang bergantung pada kepadatan, mengembangkan ambang isyarat yang mendukung fenotipe alternatif yang paling optimal. Ini akan memiliki dampak potensial bagi pemahaman kita tentang evolusi dan pemeliharaan sindrom sifat berkorelasi, seperti POLS. Tantangan berikutnya adalah menilai secara empiris seberapa luas dinamika eko-fenotipik di seluruh ektoterm, apa pendorongnya, dan sejauh mana mereka berbeda dari endoterm. Secara lebih luas, penelitian kami menyoroti peran penting yang dimainkan oleh perkembangan dalam pembentukan fenotipe, dan konsekuensi yang dapat ditimbulkannya bagi dinamika (meta)populasi dan evolusi fenotipe. Oleh karena itu, kami menyerukan pergeseran dari fokus pada hasil perkembangan di masa dewasa ke proses perkembangan yang membentuk fenotipe. Pada akhirnya, hal ini dapat mengarah pada pendekatan yang lebih mekanistik, dan dengan demikian prediktif, untuk memahami peran adaptif dari berbagai laju perkembangan dalam konteks ekologi dan evolusi.