
Abstrak
- Persaingan untuk mendapatkan makanan dan gangguan reproduksi (interaksi seksual antarspesies yang negatif) telah diidentifikasi sebagai pendorong utama pengecualian spesies. Namun, bagaimana interaksi biotik ini secara bersama-sama menentukan dominasi kompetitif masih belum diketahui secara luas. Kami mengatasi hal ini dengan menggabungkan model populasi dan eksperimen laboratorium dengan dua spesies tungau laba-laba yang bersaudara.
- Dengan menggunakan percobaan yang dirancang khusus untuk mengukur efek tunggal dan gabungan dari persaingan makanan dan gangguan reproduksi, pertama-tama kami menunjukkan bahwa kekuatan dan simetri gangguan reproduksi antara spesies berubah dengan adanya persaingan makanan.
- Selanjutnya, kami menunjukkan bahwa model populasi yang menggabungkan masing-masing jenis interaksi saja menghasilkan prediksi yang sangat berbeda, dari pengecualian sistematis salah satu dari dua spesies di bawah persaingan makanan hingga efek prioritas yang lebih memihak spesies yang sama ini (pesaing yang lebih rendah untuk makanan) di bawah pengaruh tunggal gangguan reproduksi. Selain itu, memperhitungkan pengurangan yang diamati dalam kekuatan gangguan reproduksi di hadapan persaingan makanan mengubah frekuensi ambang batas yang menentukan pesaing dominan ketika kedua interaksi tersebut berperan, dari peluang yang sama bagi kedua spesies untuk mengecualikan yang lain tergantung pada frekuensi awal mereka hingga lebih memihak pesaing yang lebih unggul untuk makanan kecuali ketika itu sangat jarang.
- Akhirnya, kami menunjukkan bahwa model tersebut menghasilkan prediksi akurat untuk dinamika populasi dalam percobaan kandang populasi independen, yang menunjukkan bahwa pendekatan kami menangkap proses paling relevan yang mengatur hasil interaksi antara spesies tungau laba-laba yang bersaing.
- Secara keseluruhan, hasil kami menunjukkan bahwa interaksi trofik dapat memodulasi interaksi seksual, yang berdampak signifikan pada dinamika populasi dan hasil kompetitif. Oleh karena itu, pertimbangan bersama tentang persaingan makanan dan gangguan reproduksi sangat penting untuk memprediksi dan memahami koeksistensi spesies secara akurat.
Baca Ringkasan Bahasa Sederhana gratis untuk artikel ini di blog Jurnal.
1. PENDAHULUAN
Bagaimana spesies yang bersaing untuk sumber daya umum hidup berdampingan bisa dibilang merupakan salah satu pertanyaan mendasar dalam ekologi (Chesson, 2000 ; Huston, 1994 ; Hutchinson, 1961 ; Tilman, 1980 ). Teori koeksistensi modern menyatakan bahwa dua spesies yang bersaing hidup berdampingan ketika perbedaan relung mereka, yaitu tingkat di mana persaingan intraspesifik melebihi persaingan interspesifik, lebih besar daripada perbedaan kebugaran mereka, yaitu perbedaan mereka dalam tingkat pertumbuhan intrinsik yang ditimbang oleh sensitivitas mereka secara keseluruhan terhadap persaingan (Adler et al., 2007 ; Chesson, 2000 , 2018 ; HilleRisLambers et al., 2012 ). Literatur yang kaya dan luas juga membahas bagaimana faktor abiotik dan biotik yang berbeda memengaruhi pendorong, dan akhirnya, hasil dari persaingan dan koeksistensi spesies (Chesson, 1994 ; Dunson & Travis, 1991 ; McPeek, 2014 ; Spaak et al., 2021 ). Secara khusus, persaingan untuk makanan sering kali terjadi bersamaan dengan jenis interaksi biotik lainnya, tetapi pengetahuan kita tentang efek gabungannya sebagian besar terbatas pada efek trofik, seperti predasi (Chesson & Kuang, 2008 ; Kotler & Holt, 1989 ; Shoemaker et al., 2020 ; Song et al., 2020 ) atau parasitisme (Hasik et al., 2023 ; Rovenolt & Tate, 2022 ; Terry et al., 2021 ), sedangkan dampak dari jenis interaksi lainnya, dalam tingkat trofik yang sama, telah menerima perhatian yang relatif lebih sedikit hingga saat ini.
Interaksi seksual antarspesies, karena pengenalan sinyal yang tidak sempurna, relatif umum terjadi di antara berbagai kelompok spesies yang bereproduksi secara seksual yang umumnya juga bersaing untuk mendapatkan makanan (Burns & Strauss, 2011 ; Kyogoku, 2015 ; Servedio & Hermisson, 2020 ; Webb et al., 2002 ; Weber & Strauss, 2016 ). Interaksi semacam itu umumnya disebut sebagai ‘gangguan reproduksi’ karena umumnya memiliki konsekuensi kebugaran negatif pada setidaknya satu spesies yang terlibat (Gröning & Hochkirch, 2008 ). Karena interaksi ini meliputi berbagai macam mekanisme dasar, mulai dari pengacauan sinyal, persaingan heterospesies, pacaran yang salah arah, dan kopulasi heterospesies (termasuk pemborosan gamet), hingga produksi hibrida yang tidak dapat hidup atau tidak subur (Gröning & Hochkirch, 2008 ), interaksi ini meliputi persaingan langsung (gangguan) dan tidak langsung (eksploitatif) untuk sumber daya bersama yang terbatas yang terkait dengan reproduksi spesies (yaitu ruang perkembangbiakan, pasangan, dan/atau gamet). Meskipun secara historis interferensi reproduksi sebagian besar telah dipelajari dalam penelitian spesiasi, karena peran utamanya dalam perpindahan dan penguatan karakter reproduksi (Butlin, 1987 ; Servedio & Noor, 2003 ; Yamaguchi & Iwasa, 2015 ), baru-baru ini hal ini menarik minat yang semakin besar dalam penelitian ekologi (Christie & Strauss, 2020 ; Cothran, 2015 ; Gómez-Llano et al., 2021 , 2023 ; Grether et al., 2024 ; Weber & Strauss, 2016 ).
Studi teoritis menyatakan bahwa interferensi reproduksi dapat menjadi pendorong utama hasil kompetitif (Kishi & Nakazawa, 2013 ; Kuno, 1992 ; Schreiber et al., 2019 ; Yamamichi et al., 2023 ; Yoshimura & Clark, 1994 ). Karena interaksi ini menyebabkan spesies lebih terdampak negatif oleh heterospesies daripada oleh konspesifik, hal ini diprediksi akan mendorong ketergantungan frekuensi positif (yakni berlawanan dengan kompetisi makanan, hal ini akan merugikan spesies yang lebih langka) dan dengan demikian tidak dapat bertindak sebagai mekanisme stabilisasi (meskipun lihat Gomez-Llano et al., 2018 ). Fenomena ini, di mana kelimpahan relatif setiap spesies menentukan spesies mana yang mengecualikan yang lain (juga dikenal sebagai efek prioritas; Grainger et al., 2019 ), menyebabkan interferensi reproduksi mendorong pengecualian lebih cepat daripada kompetisi makanan (Kuno, 1992 ; Yoshimura & Clark, 1994 ). Efek-efek ini dengan demikian dapat menghalangi koeksistensi jangka panjang antara spesies yang secara ekologis setara, sebagaimana dibuktikan oleh semakin banyaknya penelitian laboratorium dan lapangan (Hochkirch et al., 2007 ; Kishi, 2015 ; Kishi et al., 2009 ; Konishi & Takata, 2004 ; Liu et al., 2007 ; Ting & Cutter, 2018 ). Namun, seperti untuk jenis interaksi lainnya (Amarasekare, 2002 ; Chesson, 2000 ), interferensi reproduksi dapat mendorong koeksistensi melalui trade-off dengan kompetisi makanan jika kedua interaksi tersebut asimetris, sehingga spesies yang paling terpengaruh oleh kompetisi makanan kurang terpengaruh oleh interferensi reproduksi atau sebaliknya (Kishi & Nakazawa, 2013 ; Schreiber et al., 2019 ). Namun, keterbatasan dari pekerjaan sebelumnya adalah asumsi bahwa kompetisi makanan dan interferensi reproduksi beroperasi secara independen satu sama lain. Faktanya, beberapa studi empiris mengungkapkan bahwa, dalam spesies, persaingan makanan dapat mempengaruhi keberhasilan reproduksi individu (karena jantan yang lebih kenyang mungkin menjadi pesaing yang lebih kuat untuk mendapatkan pasangan, atau bahkan lebih menarik bagi betina; Fisher & Rosenthal, 2006 ; Sigurjónsdóttir, 1984 ), dan sebaliknya, strategi reproduksi yang berbeda dapat menghasilkan tingkat persaingan makanan yang berbeda (misalnya, jantan yang lebih banyak berinvestasi dalam mendapatkan pasangan mungkin menjadi pesaing yang lebih lemah untuk mendapatkan makanan dan sebaliknya; Thiel & Dennenmoser, 2007).Interaksi antara persaingan makanan dan interaksi seksual dalam spesies juga dapat terjadi antar spesies, sehingga persaingan makanan dan gangguan reproduksi dapat memengaruhi kekuatan satu sama lain jika terjadi secara bersamaan, sebagaimana yang diusulkan oleh teori terkini (Yamamichi et al., 2023 ).
Singkatnya, baik gangguan reproduksi maupun persaingan untuk mendapatkan makanan dapat berdampak besar pada koeksistensi spesies, tetapi efek gabungannya sebagian besar masih belum dieksplorasi secara eksperimental. Kesenjangan pengetahuan ini mungkin disebabkan oleh fakta bahwa, tidak seperti faktor abiotik atau biotik lainnya yang dapat dengan mudah dibedakan (misalnya suhu, salinitas, pesaing, parasit atau predator), gangguan reproduksi merupakan bagian dari interaksi antara spesies yang secara bersamaan bersaing untuk mendapatkan makanan. Oleh karena itu, merancang eksperimen yang mengukur dampak masing-masing interaksi ini secara terpisah (yang diperlukan sebagai kontrol untuk mengevaluasi efek gabungannya) merupakan tantangan yang sebenarnya. Meskipun penelitian sebelumnya berhasil memisahkan efek interaksi trofik dan reproduksi pada pertumbuhan populasi spesies yang hidup berdampingan ( capung Enallagma , Gómez-Llano et al., 2023 ; dan kumbang Callosobruchus , Kawatsu & Kishi, 2018 ) dan penelitian lain menilai efek gabungannya pada dua spesies Streptanthus jewelflower parapatrik (Christie & Strauss, 2020 ), sejauh pengetahuan kami, belum ada yang secara langsung membandingkan konsekuensi interaksi tersebut saat bertindak secara terpisah versus bersama-sama. Tungau laba-laba herbivora merupakan sistem yang ideal untuk melakukan hal tersebut, mengingat kerentanannya terhadap manipulasi eksperimental baik dalam hal persaingan makanan (Fragata et al., 2022 ; Lu et al., 2018 ; Sarmento et al., 2011 ) maupun reproduksi (termasuk gangguan reproduksi; Ben-David et al., 2009 ; Clemente et al., 2018 ; Sato & Alba, 2020 ; Smith, 1975 ).
Di sini, kami mempelajari efek tunggal dan gabungan dari gangguan reproduksi dan kompetisi untuk makanan antara dua spesies saudara tungau laba-laba, Tetranychus urticae dan T. cinnabarinus (Kuang & Cheng, 1990 ; Li et al., 2009 ), juga disebut sebagai bentuk hijau dan merah dari T. urticae (Auger et al., 2013 ). Kedua spesies ini memiliki distribusi dan rentang tanaman inang yang tumpang tindih di seluruh dunia (Migeon & Dorkeld, 2023 ), kadang-kadang ditemukan pada tanaman inang yang sama (Lu et al., 2017 ; Zélé et al., 2018 ), di mana mereka bersaing untuk mendapatkan makanan (Lu et al., 2017 , 2018 ). Spesies tungau ini juga secara alami terlibat dalam interaksi seksual heterospesifik (Smith, 1975 ) dan menderita gangguan reproduksi yang kuat karena ketidakcocokan reproduksi yang bervariasi dan hibridisasi yang tidak sesuai (mulai dari sterilitas hibrida parsial hingga lengkap dan kerusakan hibrida; Cruz et al., 2021 ; de Boer, 1982 ; Dupont, 1979 ; Sugasawa et al., 2002 ; Xue et al., 2023 ). Dengan sistem eksperimental ini, kami menggunakan kemajuan pemodelan terbaru yang berakar pada teori koeksistensi (Schreiber et al., 2019 ) dan perluasan terbarunya yang memperhitungkan efek prioritas (Ke & Letten, 2018 ) dan melakukan serangkaian eksperimen yang memanipulasi ada atau tidaknya kompetisi makanan dan gangguan reproduksi. Kombinasi teori, pemodelan, dan eksperimen ini memungkinkan kami untuk memperkirakan kekuatan kedua jenis interaksi, ketika terjadi sendiri atau bersama-sama, serta untuk memprediksi kontribusi tunggal dan gabungan mereka terhadap hasil kompetitif. Dinamika yang diprediksi dari dua spesies tungau yang berinteraksi melalui persaingan makanan dan gangguan reproduksi kemudian divalidasi menggunakan data dari percobaan populasi independen.
2 BAHAN DAN METODE
2.1 Bahan Biologis
Semua populasi yang digunakan dalam penelitian ini berasal dari satu populasi sumber dari masing-masing dua spesies tungau laba-laba, Tetranychus urticae (‘Tu’) dan T. cinnabarinus (‘Tc’), yang dikumpulkan masing-masing pada tahun 2010 dan 2013, dari lokasi yang berjarak sekitar 34 km (Cruz et al., 2021 ). Kedua populasi sumber bebas dari endosimbion yang diketahui dan sepenuhnya homozigot untuk keberadaan (Tu) atau ketiadaan (Tc) mutasi nonsinonim resesif yang mengkode resistensi pestisida, yang dapat digunakan untuk membedakan individu dari setiap spesies selain perbedaan mencolok dalam pewarnaan tubuh khusus spesies (Gambar 1 ; lihat penjelasan terperinci di Bagian S1 ). Karena isolasi pra-zigotik yang tidak lengkap (Cruz et al., 2025 ) dan isolasi pasca-zigotik yang kuat (Cruz et al., 2021 ) antara kedua populasi ini, interferensi reproduksi yang kuat diharapkan terjadi ketika mereka berbagi lingkungan yang sama (Smith, 1975 ). Memang, betina tidak menunjukkan preferensi pasangan apa pun (perkawinan non-asortatif), sedangkan jantan dari kedua spesies lebih suka kawin dengan betina Tc (Cruz et al., 2025 ). Selain itu, distorsi rasio jenis kelamin terjadi pada persilangan heterospesifik antara betina Tu dan jantan Tc (dengan produksi berlebih anak laki-laki haploid alih-alih anak perempuan hibrida, mungkin karena kegagalan pembuahan; Cruz et al., 2021 ), dan kedua arah persilangan heterospesifik menghasilkan hibrida yang sepenuhnya steril (sebagian besar betina hibrida tidak bertelur dan beberapa telur yang diletakkan tidak menetas; Cruz et al., 2021 ).

Sebelum penelitian ini, lima replikasi independen diperoleh dari masing-masing populasi sumber Tu dan Tc dengan memindahkan 200 betina dewasa ke kandang populasi yang berisi dua tanaman kacang berumur 14 hari ( Phaseolus vulgaris , cv. Bibit Contender yang diperoleh dari Germisem, Oliveira do Hospital, Portugal). Semua populasi replikasi kemudian dipelihara di bawah generasi diskret pada ukuran populasi yang konstan, dengan memindahkan 200 betina dewasa muda ke kandang populasi baru setiap 14 hari. Sebelum melakukan setiap percobaan, kelompok usia ditetapkan dari masing-masing populasi replikasi ini untuk mendapatkan jumlah individu yang cukup dengan usia yang sama (lihat rincian di Bagian S2 ). Individu dari populasi replikasi tertentu selalu diuji terhadap individu dari populasi replikasi yang sama.
2.2 Mengukur persaingan pangan, gangguan reproduksi dan dampak gabungannya
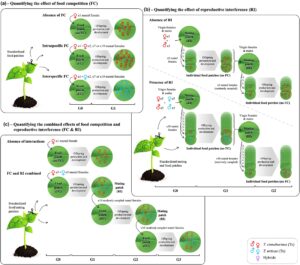
Bahasa Indonesia: Untuk memisahkan efek kompetisi makanan dan interferensi reproduksi pada pertumbuhan spesies yang berinteraksi, serta untuk mengukur efek interferensi reproduksi di hadapan kompetisi makanan, kami melakukan serangkaian tiga eksperimen berbeda, yang semuanya dirinci sepenuhnya di Bagian S2 . Pertama, untuk mengukur bagaimana kompetisi makanan memengaruhi laju pertumbuhan per kapita kedua spesies tanpa adanya interferensi reproduksi (Eksperimen 1; Gambar 2a ), betina fokus yang telah kawin dengan jantan konspesifik (sehingga tidak terpapar pada interferensi reproduksi) terpapar pada gradien kepadatan pesaing konspesifik dan heterospesifik untuk makanan, dan jumlah keturunan betina mereka dihitung untuk memperkirakan laju pertumbuhan per kapita mereka (Hart et al., 2018 ). Kedua, untuk mengukur kekuatan interferensi reproduksi sambil membatasi kompetisi untuk makanan (Eksperimen 2, Gambar 2b ), kami menghitung jumlah keturunan yang dihasilkan oleh betina yang terisolasi pada petak makanan individu (sehingga tidak terpapar pesaing sesama spesies dan/atau heterospesifik untuk makanan) setelah mereka berinteraksi secara seksual dengan jantan sesama spesies dan heterospesifik. Ini dilakukan selama 2 generasi untuk mencakup biaya yang dihasilkan dari produksi hibrida F1 steril. Ketiga, untuk mengukur efek gabungan dari kompetisi makanan dan interferensi reproduksi pada laju pertumbuhan setiap spesies (Eksperimen 3; Gambar 2c ), kami menilai produksi per kapita anak perempuan oleh betina dari setiap spesies ketika mereka berinteraksi secara seksual dan berbagi sumber daya umum selama 3 generasi. Dalam ketiga percobaan, kontrol tanpa pesaing untuk makanan maupun pasangan heterospesifik juga digunakan untuk memperkirakan laju pertumbuhan setiap spesies tanpa adanya interaksi. Analisis statistik dan data produksi keturunan yang diperoleh dalam setiap percobaan dilaporkan dalam Bagian S3 .
2.3 Pendekatan pemodelan untuk memperkirakan kekuatan interaksi dari data eksperimen dan untuk memprediksi hasil jangka panjang dari interaksi spesies
Untuk memperkirakan koefisien interaksi utama dari data yang diperoleh di setiap percobaan, kami mengadaptasi model populasi waktu-diskrit yang dikembangkan oleh Schreiber et al. ( 2019 ), di mana komponen interferensi reproduksi ditambahkan ke fungsi Beverton-Holt yang digunakan untuk memprediksi dinamika populasi di bawah kompetisi pangan (Godoy & Levine, 2014 ). Dalam model ini, jumlah individu spesies i ( N i ) diprediksi pada setiap generasi t , yang memperhitungkan interaksi dengan individu spesies j ( N j ), dengan persamaan berikut:

Akhirnya, nilai parameter yang diestimasikan digunakan untuk memprediksi hasil jangka panjang dari interaksi trofik dan/atau seksual antara populasi kami, sebagaimana dijelaskan secara lengkap di Bagian S5 . Singkatnya, karena solusi analitis belum diturunkan untuk model lengkap (Schreiber et al., 2019 ), kami malah berfokus pada tiga kondisi utama yang menjadi perhatian dalam penelitian ini:
- Jika tidak ada interferensi reproduksi (maka β ij = β ji = 0), model disederhanakan menjadi fungsi Beverton-Holt, dan koeksistensi dapat diprediksi menggunakan dua metrik utama (Chesson, 2000 ): perbedaan kebugaran rata-rata, yang merujuk pada keunggulan kompetitif satu spesies atas spesies lain; dan perbedaan relung penstabil, yang menggambarkan derajat pembatasan pertumbuhan masing-masing spesies terhadap pertumbuhan spesies pesaingnya (lihat Bagian S5 ).
- Bila tidak ada kompetisi makanan (sehingga α ii = α ij = α jj = α ji = 0), solusi analitis dapat diturunkan berdasarkan metrik kunci ketiga: kekuatan relatif gangguan reproduksi, yang berhubungan dengan tingkat sensitivitas satu spesies terhadap gangguan reproduksi dibandingkan spesies lainnya (seperti dalam Schreiber et al., 2019 ; lihat Bagian S5 ).
- Bahasa Indonesia: Ketika kedua interaksi terjadi secara bersamaan, solusi analitis hanya ada untuk kasus spesifik di mana kedua spesies sama-sama dipengaruhi oleh persaingan makanan, yaitu ketika interaksi ini simetris (Schreiber et al., 2019 ). Namun, hasil kami (lihat di bawah) mengungkapkan bahwa ini bukan kasus dalam sistem kami (seperti dalam banyak sistem lain seperti komunitas tanaman tahunan; Allen-Perkins et al., 2023 ; Godoy & Levine, 2014 ). Oleh karena itu, dalam kasus ini, untuk menentukan konsekuensi jangka panjang dari kekuatan interaksi yang diukur secara eksperimental, kami melakukan analisis numerik di mana kami memvariasikan frekuensi relatif awal dan kepadatan total individu dari setiap spesies (lihat prosedur terperinci di Bagian S5 ).
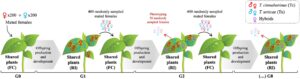
2.4 Uji empiris dinamika populasi yang diprediksi
Bahasa Indonesia: Untuk menentukan apakah koefisien interaksi yang kami estimasi memungkinkan prediksi akurat dari dinamika populasi jangka pendek, kami melakukan eksperimen skala besar (Eksperimen 4; Gambar 3 ) di mana kami mengikuti frekuensi relatif betina dari setiap spesies, serta betina hibrida, dalam populasi sangkar selama delapan generasi diskrit, sebagaimana dirinci sepenuhnya di Bagian S4 . Selain itu, kami mengadaptasi model matematika yang digunakan sebelumnya (Persamaan 1 ) agar sesuai dengan prosedur eksperimen sangkar populasi, yaitu, untuk memodelkan pengambilan sampel acak betina yang digunakan untuk menyemai setiap generasi baru, serta produksi asimetris betina hibrida steril karena kegagalan pembuahan yang terjadi dalam sistem kami (Cruz et al., 2021 ). Ini dilakukan dengan menerapkan langkah-langkah rekursif tambahan dalam model, dan parameter tambahan θ , yang sesuai dengan distorsi rasio jenis kelamin dalam persilangan heterospesifik, sebagaimana dijelaskan sepenuhnya di Bagian S6 .
Selanjutnya, kami membandingkan proporsi yang diamati dari setiap jenis betina dengan yang diprediksi oleh model, dengan mengasumsikan efek independen atau gabungan dari persaingan makanan dan gangguan reproduksi (dengan meparameterisasi model dengan estimasi dari Eksperimen 1 dan 2, atau dari Eksperimen 3, masing-masing). Karena adanya perbedaan antara proporsi betina hibrida yang diprediksi dan yang diamati (lihat Gambar S4 ), kami juga membuat dan membandingkan prediksi tambahan, dengan mengasumsikan bahwa hibrida kurang terpengaruh oleh persaingan makanan daripada betina parental oleh berbagai faktor skala parameter α dalam persamaan yang digunakan untuk memprediksi produksi hibrida (lihat Bagian S6 ). Akhirnya, kami menggunakan regresi linear antara proporsi yang diamati dan yang diprediksi dari setiap jenis betina, untuk setiap skenario, untuk menentukan parameterisasi model mana yang mengarah pada dinamika prediksi yang paling akurat (lihat detail di Bagian S6 ).
2.5 Pernyataan replikasi
Jumlah replikasi yang dilakukan di setiap percobaan yang berbeda dalam studi ini disediakan dalam Tabel 1 , dan dirinci lebih lanjut di Bagian S2 . Skala inferensi untuk semua percobaan berada pada tingkat spesies (meskipun perlu dicatat bahwa populasi sumber tunggal digunakan untuk menetapkan populasi replikasi setiap spesies). Skala di mana faktor-faktor yang diinginkan diterapkan adalah cakram daun untuk tiga percobaan pertama, dan populasi kandang untuk percobaan keempat. Studi ini tidak memerlukan persetujuan etis.
| Percobaan | Jumlah replikasi pada skala yang sesuai |
|---|---|
| 1 – Hanya kompetisi makanan | 10 replikasi a × 5 populasi b × 2 spesies fokus × 7 perlakuan = total 700 cakram daun |
| 2 – Hanya gangguan reproduksi | 10 replikasi a × 5 populasi b × 3 perlakuan (termasuk 2 spesies fokus) = total 150 cakram daun |
| 3 – Persaingan pangan dan gangguan reproduksi | 10 replikasi a × 5 populasi b × 2 spesies fokus untuk perlakuan “Interaksi” |
| 30 replikasi a × 5 populasi b × 2 spesies fokus untuk perlakuan “Tanpa interaksi” = 100 + 300 total cakram daun | |
| 4 – Dinamika populasi kandang | 5 populasi kandang b dengan 2 spesies yang dicampur (total ukuran populasi 400 dalam setiap kandang) |
Replikasi eksperimental . b Replikasi populasi.
3 HASIL
3.1 Persaingan pangan mempengaruhi kekuatan dan simetri interferensi reproduksi
Secara keseluruhan, ketiga percobaan, di mana betina dari setiap spesies dihadapkan pada kompetisi untuk makanan saja (Percobaan 1), hanya gangguan reproduksi (Percobaan 2), atau keduanya (Percobaan 3), mengungkapkan perbedaan halus dalam pertumbuhan populasi (Bagian S3 ), tetapi asimetri besar dalam tingkat pertumbuhan intrinsik spesies dan koefisien interaksi. Pertama, tingkat pertumbuhan intrinsik secara konsisten berbeda antara kedua spesies dalam ketiga percobaan: dengan tidak adanya pesaing untuk makanan dan jantan heterospesifik, betina Tc menghasilkan, rata-rata, lebih banyak keturunan daripada betina Tu (koefisien yang diperkirakan λ Tc > λ Tu sekitar 25%; Gambar 4a ; Tabel S1 dan S2 ). Kedua, ketika spesies berkompetisi hanya untuk makanan (Eksperimen 1), kedua spesies lebih sensitif terhadap pesaing heterospesifik daripada pesaing konspesifik (koefisien kompetisi yang diperkirakan α TcTu > α TcTc sekitar 8%, dan α TuTc > α TuTu sekitar 40%), tetapi betina Tc secara umum lebih sensitif terhadap kompetisi daripada betina Tu, terlepas dari identitas pesaing ( α TcTc > α TuTu sekitar 140% dan α TcTu > α TuTc sekitar 86%; Gambar 4b ; Tabel S1 ). Sebaliknya, ketika kedua spesies hanya mengalami gangguan reproduksi (Eksperimen 2), betina Tu lebih menderita akibat kehadiran heterospesifik daripada betina Tc (perkiraan koefisien gangguan reproduksi β TuTc > β TcTu sekitar 276%; Gambar 4c ; Tabel S1 ), yang menunjukkan bahwa pertukaran antara kedua jenis interaksi dapat terjadi ketika keduanya berperan.

Akhirnya, ketika kedua interaksi terjadi secara bersamaan (Eksperimen 3), kami menemukan tingkat pertumbuhan intrinsik yang lebih rendah ( λ Tc dan λ Tu ) dan kekuatan kompetisi makanan ( α TcTc , α TcTu , α TuTu , dan α TuTc ) daripada dalam eksperimen sebelumnya, tetapi kedua spesies tersebut terpengaruh oleh perubahan ini dengan cara yang sama: Betina Tc masih menghasilkan lebih banyak keturunan ( λ Tc > λ Tu sekitar 29%) dan lebih terpengaruh oleh kompetisi makanan ( α TcTc > α TuTu sekitar 200% dan α TcTu > α TuTc sekitar 167%) daripada betina Tu (Gambar 4a,b ; lihat juga Tabel S2 vs. S1 ). Namun, kami menemukan perubahan drastis dalam kekuatan dan simetri interferensi reproduksi ( β TcTu dan β TuTc ): sensitivitas betina Tc terhadap keberadaan heterospesifik sedikit meningkat dan betina Tu menurun drastis, sehingga betina Tu beralih dari lebih menderita menjadi lebih sedikit menderita dari interferensi reproduksi daripada betina Tc ( β TuTc < β TcTu sekitar 40%; Gambar 4c ). Oleh karena itu, ketika terlibat dalam kompetisi makanan dan interferensi reproduksi secara bersamaan, Tu menderita lebih sedikit daripada Tc dari kedua interaksi ini, dan pertumbuhan populasinya secara keseluruhan menjadi lebih tinggi daripada Tc sejak generasi kedua dan seterusnya (lihat Bagian S3 ).
3.2 Perubahan kekuatan gangguan reproduksi akibat kompetisi pangan berdampak pada prediksi teoritis untuk koeksistensi
Ketika kami meparameterisasi model populasi dengan koefisien yang diestimasikan dari percobaan skala kecil di mana kompetisi makanan bertindak sendiri (Percobaan 1), kami menemukan perbedaan relung negatif dalam dua populasi replikasi, karena kedua spesies membatasi pertumbuhan pesaing mereka lebih dari pertumbuhan mereka sendiri (yaitu setiap spesies lebih dipengaruhi oleh heterospesifik daripada oleh konspesifik). Ini harus mempromosikan efek dependen kepadatan positif (yaitu efek prioritas), di mana spesies dengan kelimpahan relatif yang lebih tinggi mengecualikan spesies dengan kelimpahan yang lebih rendah. Namun, untuk tiga replikasi dan rata-rata, kami juga mengamati perbedaan kebugaran yang besar antara kedua spesies, yang mengarah pada prediksi Tu mengecualikan Tc terlepas dari kelimpahan relatif kedua spesies (Gambar 5a ).
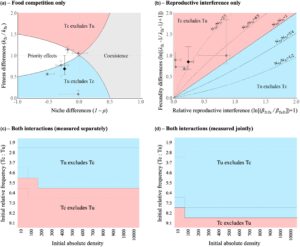
Bahasa Indonesia: Jika tidak ada kompetisi makanan, interferensi reproduksi diperkirakan akan mengarah pada efek prioritas (Schreiber et al., 2019 ), sehingga hasilnya harus bergantung tidak hanya pada kombinasi kekuatan relatif interferensi reproduksi antara dua spesies dan ‘perbedaan kesuburan’ relatif mereka, tetapi juga pada frekuensi relatif awal mereka (yaitu rasio antara kelimpahan dua spesies). Memang, kombinasi yang berbeda dari interferensi reproduksi relatif dan perbedaan kesuburan menentukan frekuensi ambang batas yang berbeda di atas mana satu spesies lebih disukai daripada yang lain (Persamaan S9 ; Gambar 5b ). Dalam sistem kami (yaitu dengan nilai parameter yang kami ukur secara eksperimental), kami memperkirakan bahwa Tc harus disukai tidak hanya ketika kedua spesies awalnya berada pada frekuensi relatif yang sama seperti dalam kondisi eksperimental kami, tetapi juga ketika Tc kurang melimpah daripada Tu (yaitu untuk semua data dan 3 populasi replikasi, itu harus dikecualikan hanya ketika frekuensinya turun di bawah 33%; dan di bawah 50% untuk replikasi lain; Gambar 5b ). Dengan demikian, persaingan makanan dan gangguan reproduksi diperkirakan akan mengarah pada hasil yang berlawanan bila dilakukan secara terpisah dalam sistem kita.
Kemudian, untuk menentukan bagaimana kedua interaksi tersebut akan secara bersama-sama memengaruhi dinamika populasi jangka panjang, kami melakukan simulasi dengan kekuatan interaksi yang diukur baik ketika kompetisi pangan dan interferensi reproduksi bertindak secara terpisah (menggunakan parameter yang diestimasikan dari Eksperimen 1 dan 2) atau bersama-sama, oleh karena itu ketika kekuatan dan simetri interferensi reproduksi berubah di hadapan kompetisi pangan (menggunakan parameter yang diestimasikan dari Eksperimen 3). Dalam kedua kasus tersebut, dan serupa dengan prediksi kami ketika interferensi reproduksi bertindak sendiri, kami menemukan efek prioritas, dengan identitas spesies yang mengecualikan yang lain bergantung pada kondisi awal (baik frekuensi relatif maupun kepadatan absolut dalam kasus ini; Gambar 5c,d ). Namun, memperhitungkan perubahan dalam kekuatan dan simetri interferensi reproduksi di bawah kompetisi pangan secara drastis mengubah frekuensi ambang batas yang menentukan spesies mana dari keduanya yang akan dominan (lihat Gambar 5d vs. Gambar 5c ). Ketika mempertimbangkan efek independen dari dua interaksi, kami menemukan bahwa interferensi reproduksi harus mengimbangi asimetri dalam kompetisi pangan, dengan pesaing terlemah untuk pangan (Tc) menyingkirkan pesaing terkuat (Tu) ketika lebih berlimpah (ambang mendekati 50% rata-rata, Gambar 5c ; lihat juga Gambar S1 dan S2 untuk variasi di antara populasi replikasi). Sebaliknya, ketika kekuatan interferensi reproduksi dimodulasi oleh kompetisi pangan, efek penyangganya sangat menurun. Dalam skenario ini, kami menemukan bahwa Tu harus menyingkirkan Tc kecuali jika frekuensinya turun di bawah 20% rata-rata (Gambar 5d ; lihat Gambar S1 dan S3 untuk variasi di antara populasi replikasi).
3.3 Model ini secara akurat memprediksi dinamika populasi kandang
Untuk memvalidasi hipotesis kami bahwa akuntansi untuk perubahan dalam kekuatan interferensi reproduksi di bawah kompetisi makanan meningkatkan akurasi prediksi model, kami membandingkan dinamika yang diamati dalam percobaan kandang populasi (Eksperimen 4; Gambar 6a ) dengan yang diprediksi oleh berbagai pemasangan model populasi. Dalam percobaan, semua populasi replikasi menunjukkan dinamika yang konsisten, dengan semua betina Tc dan hibrida dikecualikan oleh generasi ke-6 dan ke-8, masing-masing (Gambar 6b ; Tabel S3 ). Pola yang sama juga diprediksi oleh model, baik ketika memperhitungkan efek independen dari kompetisi makanan dan interferensi reproduksi dan ketika memperhitungkan pengurangan asimetris dalam interferensi reproduksi di bawah kompetisi makanan. Namun, dinamika populasi yang disimulasikan dengan mempertimbangkan kekuatan interaksi yang diukur secara independen menunjukkan varians yang sangat besar, yang menunjukkan ketidakpastian yang kuat mengenai spesies mana yang harus mengecualikan yang lain, sedangkan prediksi menunjukkan varians yang jauh lebih rendah ketika didasarkan pada kekuatan interaksi yang diukur ketika kompetisi makanan dan interferensi reproduksi bertindak bersama-sama, dengan Tu selalu mengecualikan Tc ( R2 = 0,89; Gambar S4 ). Hasil-hasil ini dengan demikian menyoroti pentingnya menggabungkan interaksi antara efek-efek dari dua interaksi untuk memprediksi dinamika populasi dengan lebih baik. Meskipun demikian, bahkan ketika kompetisi pangan dan interferensi reproduksi bertindak secara bersamaan, model tersebut memprediksi proporsi hibrida yang lebih rendah (dari generasi 2 hingga 6), dan pengecualian Tc yang lebih cepat dibandingkan dengan pengamatan eksperimental (Gambar S4B vs. Gambar 6b ). Simulasi tambahan di mana kami memvariasikan sensitivitas betina hibrida terhadap kompetisi pangan dibandingkan dengan betina ras murni (Tabel S4 ) mengungkapkan bahwa betina hibrida mungkin setidaknya 15 kali kurang sensitif terhadap kompetisi pangan daripada yang ras murni (Gambar 6c ) untuk mendapatkan kesesuaian terbaik antara dinamika yang diamati dan diprediksi ( R2 = 0,97; Gambar 6d ) sementara hanya sedikit meningkatkan AIC dari model regresi (sebesar 2,55; lihat Tabel S4 ).

4 DISKUSI
Studi ini memberikan bukti empiris pertama bahwa kekuatan gangguan reproduksi dapat dipengaruhi oleh persaingan untuk mendapatkan makanan, sehingga efek gabungan dari dua jenis interaksi pada dinamika populasi dan hasil persaingan tidak dapat diprediksi oleh tindakan independen mereka. Secara khusus, kami menunjukkan bahwa, jika tidak ada gangguan reproduksi, T. urticae (Tu) adalah pesaing yang lebih kuat untuk mendapatkan makanan dan karenanya diprediksi akan selalu menyingkirkan T. cinnabarinus (Tc). Sebaliknya, jika tidak ada persaingan untuk mendapatkan makanan, Tu lebih menderita daripada Tc dari gangguan reproduksi dan karenanya diprediksi akan disingkirkan, kecuali jika jumlahnya setidaknya dua kali lebih banyak pada awalnya (karena ketergantungan frekuensi positif). Padahal secara teoritis, trade-off antara persaingan untuk makanan dan gangguan reproduksi dapat memungkinkan koeksistensi yang stabil (Kishi & Nakazawa, 2013 ; Schreiber et al., 2019 ), simulasi kami malah menunjukkan bahwa ketergantungan frekuensi positif yang didorong oleh gangguan reproduksi harus tetap ada ketika kedua jenis interaksi tersebut terjadi secara bersamaan (identitas spesies yang bertahan bergantung pada frekuensinya), baik dengan efek independen maupun yang berinteraksi. Ketika kedua jenis interaksi diasumsikan memiliki efek independen, persaingan makanan harus menyeimbangkan keuntungan yang diberikan kepada Tc oleh gangguan reproduksi, dengan kedua spesies memiliki kemungkinan yang sama untuk bertahan pada frekuensi awal yang genap. Namun, ketika kekuatan gangguan reproduksi dapat dimodulasi oleh persaingan makanan, pesaing yang lebih unggul untuk makanan, Tu, menjadi yang paling tidak sensitif terhadap gangguan reproduksi. Prediksi kami yang memperhitungkan perubahan tersebut menunjukkan pengecualian Tc dalam semua skenario yang diuji, kecuali ketika awalnya sangat melimpah dibandingkan dengan pesaingnya, Tu. Hasil yang diperoleh dari eksperimen kandang populasi independen sebagian besar sesuai dengan prediksi ini, dengan Tc secara sistematis dikecualikan ketika kedua spesies memulai pada frekuensi awal yang genap. Oleh karena itu, hasil kami memberikan dua pelajaran langsung: pertama, persaingan makanan memodulasi gangguan reproduksi, yang menunjukkan bahwa interaksi ini adalah proses yang saling terkait dengan efek non-independen pada koeksistensi spesies, dan kedua, teori koeksistensi modern (Schreiber et al., 2019 ; Yamamichi et al., 2023 ) adalah kerangka kerja yang cocok untuk memprediksi efeknya pada dinamika populasi dan hasil kompetitif spesies yang berkerabat dekat.
Banyak penelitian telah menyelidiki kekuatan isolasi reproduksi antara Tu dan Tc (Cruz et al., 2021 , 2025 ; Murtaugh & Wrensch, 1978 ; Sugasawa et al., 2002 ; Xue et al., 2023 ). Namun, interferensi reproduksi dan konsekuensinya terhadap dinamika populasi hampir tidak dipertimbangkan (tetapi lihat Murtaugh & Wrensch, 1978 ). Di sini, dengan tidak adanya persaingan untuk makanan, kami menemukan interferensi reproduksi asimetris antara Tc dan Tu, hasil yang kongruen dengan temuan sebelumnya tentang isolasi reproduksi pra dan pasca perkawinan antara kedua populasi yang digunakan dalam penelitian ini (Cruz et al., 2021 , 2025 ). Perhatikan, bagaimanapun, bahwa arah asimetri dalam interferensi reproduksi yang ditemukan di sini tidak intuitif mengingat hambatan reproduksi yang diidentifikasi dalam studi sebelumnya tentang sistem ini, di mana persilangan antara betina Tu dan jantan Tc mengakibatkan produksi berlebih dari keturunan jantan (Tu) karena kegagalan pembuahan, sedangkan jantan dari kedua spesies lebih suka kawin dengan betina Tc (Cruz et al., 2021 , 2025 ). Dengan demikian, ini akan meningkatkan risiko perkawinan heterospesifik untuk betina Tc dibandingkan dengan betina Tu, sehingga membuat mereka lebih mungkin mengalami interferensi reproduksi. Fakta bahwa kami mengamati pola yang berlawanan di sini menyoroti bahwa hubungan antara hambatan reproduksi dan interferensi reproduksi tidak sesederhana yang mungkin dipikirkan dan bahwa studi lebih lanjut diperlukan untuk memahami interaksi kompleks antara mekanisme yang berbeda yang mendasari interferensi reproduksi (seperti dalam Gómez-Llano et al., 2023 ). Namun, hasil studi saat ini sesuai dengan studi sebelumnya yang menggunakan populasi Tiongkok dari spesies yang sama, baik di laboratorium (Lu et al., 2017 ) dan di lapangan (Lu et al., 2018 ). Memang, studi-studi ini mengungkapkan bahwa Tc secara konsisten menggantikan Tu ketika pada frekuensi awal yang genap, meskipun hasil ini terbalik dengan adanya pestisida karena resistensi pestisida yang lebih kuat pada Tu. Faktanya, setiap faktor abiotik atau biotik yang memengaruhi kebugaran relatif kedua spesies ini, seperti suhu (Gotoh et al., 2015 ; Lu et al., 2017 ; Riahi et al., 2013 ; Zou et al., 2018 ), tanaman inang (Huo et al., 2021 ; Tomczyk et al., 1995 ; Witul & Kielkiewicz, 1999 ), atau musuh alami (Takabayashi et al., 2000 ) dapat secara signifikan menggantikan Tu dalam populasi Tiongkok.), juga harus berperan dalam menentukan ketahanan mereka dalam lingkungan alami. Namun, sejauh ini belum ada penelitian yang bertujuan untuk mengurai peran relatif faktor-faktor tersebut dalam membentuk hasil kompetitif, apalagi peran berbagai faktor intrinsik.
Pekerjaan teoritis sebelumnya meramalkan bahwa persaingan makanan dan interferensi reproduksi yang bertindak dalam arah yang berlawanan (yaitu di bawah trade-off) dapat mendukung koeksistensi yang stabil kecuali ketika kekuatan interferensi reproduksi sangat tinggi (Kishi & Nakazawa, 2013 ). Meskipun kami menemukan bahwa persaingan untuk makanan menyeimbangkan keuntungan yang diberikan oleh interferensi reproduksi kepada Tc dalam penelitian saat ini, koeksistensi tidak diprediksi dalam skenario apa pun. Sebaliknya, dinamika populasi yang disimulasikan mengungkapkan bahwa ketergantungan frekuensi positif yang didorong oleh interferensi reproduksi harus terjadi bahkan di hadapan persaingan makanan. Ini mungkin karena fakta bahwa kami menemukan perbedaan relung negatif dalam sistem kami, sebuah skenario yang belum dieksplorasi meskipun beberapa studi teoritis dan empiris dalam literatur koeksistensi menunjukkan bahwa efek prioritas harus menjadi hasil umum dari interaksi ekologis (Fragata et al., 2022 ; Ke & Letten, 2018 ; Song et al., 2021 ; Spaak et al., 2021 ). Perbedaan relung negatif dalam studi kami, bagaimanapun, bisa jadi merupakan konsekuensi dari desain eksperimen kami, di mana hanya satu jenis sumber daya dan tidak ada heterogenitas spasial yang tersedia, sehingga sangat menghalangi penghindaran pesaing. Sebaliknya, dalam populasi alami, kedua spesies memiliki rentang tanaman inang yang serupa tetapi sangat luas (Migeon & Dorkeld, 2023 ), dan persaingan makanan dapat menggeser rentang ini di lingkungan dengan lebih dari satu spesies tanaman (Ferragut et al., 2013 ). Dengan demikian, penghindaran pesaing dalam populasi alami dapat mendorong tumpang tindih relung yang lebih rendah antara spesies, yang mendorong koeksistensi dalam skala yang lebih luas (Wittmann & Fukami, 2018 ). Namun, bukti tungau laba-laba yang menghindari pesaing interspesifik beragam (Godinho et al., 2024 ; Pallini et al., 1997 ; Sato et al., 2016 ), oleh karena itu tidak jelas apakah mereka akan menampilkan perilaku ini dalam semua keadaan. Selain itu, sebagai hama tanaman, mereka sering kali terpapar pada lingkungan yang relatif homogen, di mana hasil yang diuraikan di sini bermakna.
Percobaan kami secara khusus dirancang untuk menguji perubahan kekuatan interferensi reproduksi dalam menanggapi kompetisi pangan. Kemungkinan ini belum diselidiki, baik secara teoritis maupun empiris. Namun, ada bentuk lain di mana kedua interaksi ini dapat memengaruhi satu sama lain. Memang, karya teoritis baru-baru ini juga menunjukkan bahwa perubahan intensitas kompetisi pangan yang dimediasi oleh interferensi reproduksi dapat mendorong koeksistensi dengan mengubah perbedaan relung yang awalnya negatif menjadi positif (Yamamichi et al., 2023 ) bahkan tanpa adanya sumber daya alternatif (yaitu karena perubahan perilaku dan/atau evolusi; Kishi & Tsubaki, 2014 ; Noriyuki & Osawa, 2016 ; Ruokolainen & Hanski, 2016 ). Namun, analisis menyeluruh dan sistematis tentang efek gabungan dari kedua interaksi ini masih dalam tahap awal.
Uji eksperimental dari efek gabungan dari gangguan reproduksi dan kompetisi untuk makanan mengungkapkan bahwa ini tidak setara dengan kombinasi efek independennya. Sementara laju pertumbuhan intrinsik kedua spesies dan sensitivitas mereka terhadap kompetisi secara keseluruhan secara konsisten lebih rendah karena efek lingkungan yang tidak diketahui (Eksperimen 3), intensitas gangguan reproduksi menjadi serupa untuk kedua spesies di hadapan kompetisi makanan, sementara itu lebih kuat untuk Tu daripada untuk Tc tanpanya (Eksperimen 2). Ini dapat dijelaskan oleh korelasi positif antara sifat fenotipik yang terlibat dalam masing-masing interaksi ini, atau oleh sifat yang sama yang terlibat dalam (dan dipengaruhi oleh) kedua interaksi (Maan & Seehausen, 2011 ). Misalnya, ukuran tubuh biasanya merupakan sifat utama untuk interaksi trofik dan seksual (Okuzaki et al., 2010 ). Pada tungau laba-laba, berkurangnya ketersediaan makanan dapat berdampak negatif pada ukuran tubuh (Li & Zhang, 2018 ), sedangkan jantan yang lebih besar umumnya merupakan pesaing yang lebih unggul untuk pasangan (Enders, 1993 ; Potter et al., 1976 ), dan betina yang lebih besar lebih disukai daripada yang lebih kecil (Edward & Chapman, 2011 ; Zahradnik et al., 2008 ). Oleh karena itu, karena individu Tu (pesaing unggul untuk makanan) kurang terpengaruh oleh persaingan makanan, mereka dapat mempertahankan ukuran tubuh yang lebih besar pada generasi berikutnya. Ini akan meningkatkan frekuensi perkawinan antara kedua jenis betina dan jantan Tu (jika ini lebih besar dari jantan Tc, mereka akan menjadi lebih kompetitif), sehingga mengurangi kekuatan gangguan reproduksi untuk betina Tu, tetapi meningkatkannya untuk yang Tc. Alternatifnya, atau sebagai tambahan, berkurangnya ketersediaan makanan juga dapat menyebabkan perkembangan keturunan yang lebih lambat pada tungau laba-laba (Wilson, 1994 ), seperti pada sistem artropoda lainnya (misalnya Teng & Apperson, 2000 ), sehingga jantan Tu, yang merupakan pesaing unggul untuk makanan, dapat berkembang lebih cepat dan mendapatkan pasangan sejenis sebelum jantan Tc menjadi dewasa secara seksual, dengan konsekuensi serupa seperti yang baru saja dijelaskan di atas. Mekanisme lain mungkin juga melibatkan produksi sinyal dengan kualitas lebih rendah, atau produksi gamet yang lebih sedikit/lebih kecil/kurang bergerak oleh pesaing yang lebih rendah. Secara keseluruhan, persaingan makanan dapat memengaruhi nilai sifat lebih parah pada keturunan pesaing yang lebih rendah, yang pada gilirannya menjadi lebih terpengaruh oleh gangguan reproduksi. Alternatifnya, perubahan kekuatan gangguan reproduksi dengan adanya persaingan makanan mungkin hanya muncul melalui efek yang terakhir miliki pada kepadatan populasi, dengan asumsi bahwa kekuatan gangguan reproduksi ( β) bervariasi dengan kepadatan populasi total (dengan cara yang sama seperti kekuatan persaingan pangan dapat berubah dengan adanya gangguan reproduksi jika nilai α bergantung pada kepadatan, seperti yang diusulkan oleh Yamamichi et al., 2023 ). Namun, agar hipotesis ini dapat menjelaskan hasil kami, nilai β harus memiliki sensitivitas terhadap kepadatan yang spesifik terhadap spesies.
Terlepas dari mekanisme yang mendasari interaksi antara interaksi kompetitif dan seksual, simulasi kami mengungkapkan bahwa perubahan dalam kekuatan interferensi reproduksi sebagai respons terhadap kompetisi makanan (dibandingkan dengan ketidakhadirannya) menyebabkan peningkatan yang kuat dalam frekuensi awal ambang batas di mana pesaing yang lebih rendah untuk makanan (Tc) tidak dapat bertahan. Kemudian, dengan menggunakan data independen untuk parameterisasi dan validasi model, kami menunjukkan pentingnya memperhitungkan efek ini untuk secara akurat memprediksi dinamika populasi, dengan demikian lebih jauh menyoroti pentingnya memahami bagaimana interaksi trofik dan seksual saling memengaruhi. Karya terakhir ini juga menunjukkan bahwa model sederhana yang menangkap efek demografis dari interferensi reproduksi hanya dengan dua parameter ( β dan θ ; lih. Metode), karenanya tidak secara eksplisit memodelkan masing-masing mekanisme yang mendasarinya (atau ‘komponen kebugaran’; Kyogoku, 2015 ), dapat menghasilkan prediksi yang sangat akurat untuk dinamika dua spesies dalam populasi kandang eksperimental. Kemampuan untuk memprediksi dinamika sistem ini lebih ditingkatkan ketika memperhitungkan produksi betina hibrida yang kurang sensitif terhadap kompetisi makanan. Sementara dampak ekologis hibrida dipelajari secara luas dalam konteks adaptasi (Gow et al., 2007 ; Seehausen, 2004 ), dampak demografis hibrida yang ‘tidak cocok’ pada spesies induk sebagian besar diabaikan. Faktanya, peningkatan frekuensi hibrida dalam populasi induk (seperti yang diprediksi dan diamati di sini) dapat menghasilkan perubahan yang tidak terduga dalam dinamika populasi, terutama jika mereka bersaing ketat untuk mendapatkan makanan dan/atau jika mereka sangat menarik bagi pejantan, terlepas dari kesuburannya. Namun, sepengetahuan kami, belum diketahui apakah betina hibrida dalam sistem ini dikenali atau bahkan lebih disukai sebagai calon pasangan oleh pejantan dari salah satu spesies atau apakah mereka merupakan pesaing kuat untuk mendapatkan makanan. Namun demikian, konsisten dengan pengamatan yang dilakukan di laboratorium kami, analisis kami mengungkapkan bahwa mereka mungkin membutuhkan sumber daya yang jauh lebih sedikit daripada betina ras murni, mungkin karena kebanyakan dari mereka tidak menghasilkan telur (Cruz et al., 2021 ).
Studi kami merupakan upaya pertama namun ketat untuk menyelidiki kompleksitas evaluasi interaksi ekologis secara eksperimental, sementara secara bersamaan memperhitungkan persaingan untuk makanan dan pasangan. Kami telah melakukan yang terbaik yang dapat kami lakukan mengingat keterbatasan logistik kami, tetapi studi yang lebih besar dapat membahas aspek-aspek tambahan yang tidak tercakup dalam studi kami untuk sepenuhnya memahami interaksi kompleks mereka. Misalnya, kami dapat merancang eksperimen untuk menentukan apakah hubungan antara kepekaan setiap spesies terhadap persaingan intraspesifik dan interspesifik juga berubah dengan adanya gangguan reproduksi atau apakah dan bagaimana gangguan reproduksi memengaruhi intensitas persaingan makanan (yaitu kebalikan dari apa yang kami uji di sini) seperti yang diusulkan oleh Yamamichi et al. ( 2023 ). Misalnya, pelecehan kawin jantan dalam spesies (Oku, 2009 ) dapat menurunkan atau meningkatkan intensitas persaingan makanan, masing-masing dengan mengurangi waktu makan betina karena mereka menginvestasikan lebih banyak waktu untuk menghindari upaya kawin jantan (Bancroft & Margolies, 1996 ), atau dengan meningkatkan agregasi spasial betina sebagai perlindungan dari jantan (Yamamichi et al., 2023 ). Lebih jauh lagi, model tersebut dapat dikembangkan untuk memperhitungkan interaksi reproduksi negatif dalam spesies, seperti pelecehan jantan, yang mungkin juga memainkan peran penting dalam menentukan kekuatan interferensi reproduksi antarpopulasi (Kyogoku & Sota, 2017 ). Interaksi antara jantan dan betina konspesifik bahkan dapat diubah oleh kehadiran heterospesifik, yang berpotensi menghasilkan fasilitasi daripada interferensi (Gomez-Llano et al., 2018 ). Perkembangan teoritis seperti itu pada akhirnya dapat memungkinkan perluasan konsep perbedaan ceruk tidak hanya pada interaksi trofik tetapi juga interaksi seksual, yang mengarah pada integrasi yang lebih kuat ke dalam kerangka koeksistensi umum (Gómez-Llano et al., 2021 ).
Kesimpulannya, studi kami mengungkap bahwa kompetisi pangan dapat memengaruhi kekuatan interferensi reproduksi, dengan konsekuensi signifikan bagi dinamika spesies yang bersaing. Mengingat bahwa interferensi reproduksi diperkirakan sering terjadi antara spesies yang bersaing untuk mendapatkan makanan (Servedio & Hermisson, 2020 ), hasil seperti itu kemungkinan merupakan fitur umum dari banyak sistem ekologi yang melibatkan taksa yang secara kebetulan terlibat dalam interaksi seksual (Weber & Strauss, 2016 ), dan mungkin memiliki konsekuensi besar bagi studi koeksistensi ekologis (Germain et al., 2018 ; Gómez-Llano et al., 2021 ). Memang, kami menunjukkan bahwa mengatasi efek dari setiap jenis interaksi saja mungkin tidak cukup untuk memprediksi dinamika populasi secara akurat. Ini karena ko-kemunculan kedua jenis interaksi dapat mengubah bentuk trade-off antara interferensi reproduksi dan kompetisi untuk mendapatkan makanan, yang menyebabkan ketidakseimbangan prioritas satu spesies atas spesies lain dalam sistem dengan perbedaan relung negatif (sementara berpotensi menghambat koeksistensi dalam sistem dengan perbedaan relung positif). Memahami interaksi antara makan dan interaksi seksual ini juga penting untuk penelitian spesiasi, karena kemungkinan dan durasi koeksistensi antara spesies yang terkait erat akan menentukan peluang untuk penguatan isolasi reproduksi pra-zigotik. Temuan kami dengan demikian mungkin memiliki konsekuensi yang luas untuk bidang yang baru-baru ini berkembang di antarmuka antara teori spesiasi dan koeksistensi (Boussens-Dumon & Llaurens, 2021 ; Germain et al., 2018 ; Grether et al., 2020 ; Kyogoku & Kokko, 2020 ; Kyogoku & Wheatcroft, 2020 ). Akhirnya, meskipun interaksi spesies dipelajari secara intensif selama lebih dari satu abad sebagai penentu utama distribusi spesies dan hasil kompetitif, hasil kami secara kolektif menunjukkan bahwa kita masih dalam tahap awal dalam memahami bagaimana mekanisme yang berbeda berinteraksi untuk menentukan dinamika populasi spesies yang berinteraksi. Menggabungkan pendekatan teoritis dan empiris adalah kunci untuk mengungkap bagaimana berbagai jenis interaksi secara bersama-sama memengaruhi koeksistensi atau pengecualian spesies.


