Abstrak
- Ekosistem perkotaan berkembang pesat, secara signifikan mengubah lanskap alam dan memengaruhi keanekaragaman hayati.
- Di sini kami mengeksplorasi variasi musiman dalam keanekaragaman mamalia menggunakan DNA lingkungan (eDNA) dari sampel tanah yang dikumpulkan selama musim dingin dan musim panas di 21 taman kota di Detroit, Michigan. Kami memperkirakan keanekaragaman gamma (regional), alfa (lokal) dan beta (perubahan komposisi) untuk menentukan apakah pergeseran musiman, yang mencerminkan kelangkaan musim dingin dan kelimpahan musim panas dalam komposisi komunitas mamalia dan aktivitas manusia, dapat dideteksi menggunakan eDNA. Kami berharap bahwa taman yang lebih besar akan menunjukkan keanekaragaman yang lebih besar dan pergantian musiman yang lebih tinggi, konsisten dengan hubungan spesies-area (SAR) dan berhipotesis bahwa peningkatan ketersediaan sumber daya musim panas akan menyebabkan penurunan kepadatan jaringan karena spesies menyebar lebih luas.
- Kami menemukan bahwa taman kota menunjukkan perubahan halus yang spesifik pada komposisi komunitas taman yang dipengaruhi oleh faktor ekologi dan antropogenik, dengan spesies termasuk sigung belang, tikus cokelat, dan babi tanah yang bertanggung jawab atas variasi musiman yang diamati. Konsisten dengan SAR, taman yang lebih besar mendukung kekayaan dan keanekaragaman spesies yang lebih tinggi. Analisis jaringan ekologi, yang berfokus pada metrik seperti koefisien pengelompokan dan kepadatan jaringan, mengungkapkan penurunan konektivitas dan kohesivitas interaksi spesies secara keseluruhan dari musim dingin ke musim panas, yang mendukung hipotesis kami tentang penyebaran spesies yang lebih luas selama periode kaya sumber daya. Khususnya, DNA manusia lazim di semua taman, di samping deteksi eDNA babi dan sapi, yang berpotensi mencerminkan gangguan manusia dan masukan makanan antropogenik.
- Temuan kami menggarisbawahi kemanjuran analisis eDNA dalam menangkap dinamika komunitas mamalia perkotaan, dampak aktivitas manusia terhadap keanekaragaman hayati, dan potensinya sebagai alat yang berharga untuk penelitian ekologi perkotaan. Pada akhirnya, peningkatan kapasitas pemantauan membantu upaya konservasi dan perencanaan perkotaan yang akan mendorong koeksistensi manusia-satwa liar dan melestarikan manfaat sosial-ekologis yang berasal dari keanekaragaman hayati di seluruh lanskap kota.
1. PENDAHULUAN
Urbanisasi adalah tren global yang dominan, dengan lebih dari setengah populasi dunia sekarang tinggal di kota-kota (Derby Lewis et al., 2019 ). Sementara daerah perkotaan mencakup 2% lahan di Amerika Utara, kota-kota diproyeksikan akan terus berkembang secara signifikan dalam beberapa dekade mendatang (Simkin et al., 2022 ). Pada tahun 2030, tutupan lahan perkotaan diperkirakan akan meningkat sekitar 1,2 juta kilometer persegi secara global, dengan Amerika Utara berkontribusi pada sebagian besar pertumbuhan ini (Mahtta et al., 2022 ). Transformasi ini menggantikan lanskap alam dengan lingkungan yang dibangun, mengubah keanekaragaman hayati dan proses ekosistem melalui fragmentasi habitat, pulau panas, dan jaringan jalan (Chen et al., 2020 ; Li et al., 2022 ). Bagi spesies bergerak, area perkotaan dapat berfungsi sebagai tempat berlindung sekaligus bahaya, dikotomi yang dipengaruhi oleh peningkatan ketersediaan makanan, pelepasan predator, dan peningkatan risiko kematian (Bateman & Fleming, 2012 ; Fehlmann et al., 2021 ). Lingkungan binaan sering kali mendorong pergantian spesies, atau penggantian spesies dalam suatu komunitas dari waktu ke waktu atau ruang, yang menguntungkan spesies yang beradaptasi dengan perkotaan sambil memindahkan spesies spesialis ke area pinggiran yang kurang terganggu (Gallo et al., 2017 ). Seiring waktu, ekosistem perkotaan dapat mengalami homogenisasi biotik, di mana spesies generalis menjadi dominan dengan mengorbankan keanekaragaman komunitas (Alberti et al., 2020 ; Haight et al., 2023 ; Lowry et al., 2013 ). Pergeseran komposisi komunitas tersebut dapat memiliki implikasi signifikan bagi stabilitas dan fungsi ekosistem (Hahs et al., 2023 ; McKinney, 2006 ).
Urbanisasi telah terbukti menyederhanakan jaringan ko-keberadaan spesies, yang berpotensi mengurangi redundansi ekologis dan membuat komunitas lebih rentan terhadap gangguan lingkungan (Pechlivanis et al., 2024 ). Gangguan antropogenik dapat merestrukturisasi jaringan interaksi mamalia, mengubah pola ko-keberadaan spasial dan memodifikasi dinamika komunitas (Curveira-Santos et al., 2024 ). Meskipun ada tantangan koeksistensi dengan manusia, ruang hijau di lingkungan perkotaan berpotensi bertindak sebagai oasis bagi satwa liar, mendukung keanekaragaman hayati dan menjaga konektivitas ekologis (Ives et al., 2016 ; Rega-Brodsky et al., 2022 ). Hubungan spesies-area (SAR) menunjukkan bahwa ruang hijau yang lebih besar umumnya mendukung keanekaragaman spesies yang lebih tinggi karena ketersediaan habitat dan sumber daya yang lebih luas (Kallimanis et al., 2008 ). Seiring dengan meningkatnya luas habitat, jumlah spesies yang dapat didukungnya biasanya meningkat secara logaritmik, menjadikan ukuran ruang terbuka hijau perkotaan sebagai faktor penting dalam melestarikan keanekaragaman hayati (Beninde et al., 2015 ). Ketersediaan sumber daya dan gangguan manusia merupakan faktor utama yang membentuk interaksi spesies dan struktur jaringan di lanskap perkotaan; di dalam ruang-ruang ini, jaringan interaksi spesies dapat berubah sebagai respons terhadap fluktuasi musiman dalam kelimpahan sumber daya, aktivitas manusia, dan kondisi habitat, yang selanjutnya memengaruhi dinamika ekologi (Gilbert et al., 2022 ).
Bersamaan dengan pergeseran dalam struktur komunitas, musim juga secara krusial membentuk komunitas hewan karena dinamika dalam iklim dan sumber daya makanan selanjutnya mengubah interaksi, distribusi, dan perilaku spesies (Porter et al., 2022 ; White & Hastings, 2020 ; Gambar 1 ). Namun, urbanisasi dapat bertindak sebagai penyangga terhadap ekstrem musiman, mengurangi efek kelangkaan makanan musim dingin dan fluktuasi suhu melalui efek pulau panas perkotaan dan sumber makanan antropogenik (Filazzola et al., 2024 ; Marcacci et al., 2023 ; Vlaschenko et al., 2023 ). Subsidi antropogenik memengaruhi spesies yang berhibernasi dengan mengubah ketersediaan sumber daya. Misalnya, suhu yang lebih hangat dan peningkatan akses ke makanan manusia telah mengurangi periode hibernasi beruang hitam ( Ursus americanus ) di Colorado, AS, yang berpotensi meningkatkan konflik dan kematian manusia-beruang (Johnson et al., 2018 ). Musiman juga membentuk aktivitas manusia, yang pada gilirannya memodulasi perilaku satwa liar (Doherty et al., 2021 ). Misalnya, rekreasi manusia di siang hari telah menyebabkan beruang hitam dan anjing hutan ( Canis latrans ) mengalihkan aktivitas mereka ke periode nokturnal, sementara spesies yang lebih sensitif seperti kucing hutan ( Lynx rufus ) mengurangi penggunaan habitat mereka di dekat jalan setapak (Lewis et al., 2021 ). Pola-pola ini menyoroti bagaimana lingkungan perkotaan merestrukturisasi interaksi spesies, dengan konsekuensi potensial bagi jaringan ekologi dan stabilitas komunitas.
Pelacakan dinamika spasial dan temporal sangat penting untuk memahami bagaimana fragmentasi habitat, ketersediaan sumber daya, dan aktivitas manusia membentuk keanekaragaman hayati di lingkungan perkotaan (Leveau et al., 2021 ; Malhotra et al., 2022 ). Metode pemantauan seperti perangkap kamera, sensor akustik, dan observasi sains komunitas memberikan wawasan tentang keberadaan dan aktivitas spesies (misalnya Gámez & Harris, 2021 ; Gibb et al., 2019 ; Pecorelli et al., 2019 ) dan teknologi canggih seperti pelacakan GPS dan citra satelit memberikan wawasan lebih jauh tentang pola pergerakan dan penggunaan habitat di berbagai musim (Brown et al., 2023 ; McDuie et al., 2019 ; Severson et al., 2021 ). Namun, metode ini bergantung pada deteksi langsung dan mungkin kurang mewakili spesies yang tersembunyi atau nokturnal, sehingga menimbulkan bias terkait ukuran tubuh, perilaku, dan probabilitas deteksi (Newey et al., 2015 ). DNA Lingkungan (eDNA), yang menangkap materi genetik yang dilepaskan oleh organisme ke lingkungan sekitarnya, menyediakan pendekatan alternatif untuk pemantauan keanekaragaman hayati, khususnya di lingkungan yang sulit untuk melakukan pengamatan langsung (Clare et al., 2022 ; Hallam et al., 2021 ). Tidak seperti perangkap kamera, yang utamanya mendeteksi mamalia darat berukuran sedang hingga besar yang melewati jangkauan sensor, eDNA memungkinkan deteksi spesies yang lebih luas, termasuk taksa yang lebih kecil, sulit ditangkap, dan nokturnal yang mungkin tidak tertangkap oleh metode survei tradisional (Leempoel et al., 2020 ). Dalam ekosistem perkotaan, tempat fragmentasi habitat dan tekanan antropogenik mendorong pola keanekaragaman hayati, eDNA dapat menyediakan alat yang ampuh untuk mendeteksi pergeseran musiman dalam struktur komunitas yang mungkin terabaikan (Johnson et al., 2021 ).
Mamalia memainkan peran ekologis utama di lingkungan perkotaan, memengaruhi ekosistem melalui pengaturan mangsa, penyebaran benih, dan interaksi fungsional lainnya (Lacher et al., 2019 ; Russo et al., 2016 ), sementara juga berinteraksi dengan populasi manusia melalui penularan penyakit, pengendalian hama pertanian, dan signifikansi budaya (Ávila-Nájera et al., 2018 ; Hunold & Mazuchowski, 2020 ; Shultz et al., 2023 ). Memahami pergeseran musiman dalam keanekaragaman mamalia perkotaan sangat penting untuk menilai bagaimana aktivitas manusia dan perubahan lingkungan memengaruhi populasi satwa liar. Sementara penelitian telah meneliti keanekaragaman spesies perkotaan, lebih sedikit perhatian diberikan pada peran musiman dalam menyusun komunitas mamalia perkotaan, khususnya dalam konteks interaksi spesies dan dinamika jaringan. Interaksi spesies membentuk jaringan ekologi, dan menyelidiki bagaimana hubungan ini bergeser secara musiman adalah kunci untuk memahami ketahanan dan fungsi ekosistem perkotaan.
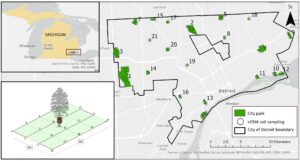
Di sini, kami menyelidiki variasi musiman dalam keanekaragaman mamalia, komposisi komunitas, dan jaringan interaksi di seluruh taman kota. Dengan menggunakan eDNA dari sampel tanah yang dikumpulkan selama musim dingin dan musim panas di 21 taman kota di Detroit, Michigan (Gambar 2 ), kami menjawab pertanyaan penelitian berikut: (1) Bagaimana variasi musiman memengaruhi kekayaan spesies mamalia (keragaman alfa), komposisi komunitas (keragaman beta), dan kumpulan spesies secara keseluruhan (keragaman gamma) di taman kota? (2) Bagaimana karakteristik taman, seperti ukuran dan aktivitas manusia, membentuk pola musiman dalam komunitas mamalia? (3) Bagaimana jaringan interaksi spesies berbeda antara musim dingin dan musim panas, dan apa yang diungkapkan oleh perbedaan ini tentang pergeseran musiman dalam ekologi mamalia perkotaan? Kami berhipotesis bahwa keanekaragaman gamma akan tetap stabil di seluruh musim, sementara keanekaragaman alfa dan beta akan berfluktuasi, yang mencerminkan perilaku musiman khusus spesies. Kami memperkirakan taman yang lebih besar akan menunjukkan pergantian musiman yang lebih besar, karena heterogenitas habitat mendukung spesies dengan pola aktivitas musiman yang berbeda. Selain itu, kami mengantisipasi perubahan musiman dalam eDNA yang terkait dengan manusia, yang mencerminkan pola penggunaan taman dan pergeseran dalam interaksi satwa liar perkotaan. Kami memperkirakan pergeseran musiman dalam struktur jaringan mamalia yang didorong oleh ketersediaan sumber daya. Di musim panas, peningkatan sumber daya dapat menyebabkan penyebaran yang lebih luas, mengurangi sentralitas derajat dan kepadatan jaringan. Di musim dingin, makanan dan tempat berlindung yang terbatas dapat memaksa spesies untuk saling berdekatan, meningkatkan pengelompokan dan kepadatan jaringan. Kami juga memperkirakan manusia dan spesies domestik akan memainkan peran utama dalam jaringan ini, dengan ko-kemunculan yang lebih tinggi dengan satwa liar selama musim panas ketika aktivitas manusia di ruang hijau perkotaan lebih besar. Temuan kami memberikan wawasan tentang bagaimana lingkungan perkotaan dan dinamika musiman membentuk keanekaragaman hayati, menginformasikan strategi konservasi dan perencanaan perkotaan untuk mendukung satwa liar di kota.
2.1 Wilayah Studi
Kami mengumpulkan sampel tanah dari 21 taman kota di Detroit, kota terbesar di Michigan (Gambar 2 ). Karena pengambilan sampelnya non-invasif, tidak diperlukan persetujuan etika hewan dan izin tidak diperlukan mengingat jumlah pengambilan tanah yang sedikit. Taman-taman ini termasuk dalam studi penjebakan kamera jangka panjang dan ukurannya bervariasi dari ~1,6 hingga 480 ha. Selain itu, taman yang diambil sampelnya bervariasi dalam habitat, tingkat kunjungan manusia, dan tingkat gangguan yang berasal dari keberadaan hewan peliharaan (Gámez & Harris, 2021 ; Lima et al., 2021 ). Di Detroit, karnivora asli terbesar yang ada adalah coyote, yang hidup berdampingan dengan rubah abu-abu ( Urocyon cinereoargenteus ), rubah merah ( Vulpes vulpes ), rakun ( Procyon lotor ), oposum Virginia ( Didelphis virginiana ) dan sigung belang ( Mephitis mephitis ). Herbivora seperti rusa berekor putih ( Odocoileus virginianus ) dan kelinci ekor kapas timur ( Sylvilagus floridanus ) hadir bersama dengan sedikitnya 15 spesies dari ordo Rodentia (iNaturalist, 2024 ). Terletak di tenggara Michigan, Detroit memiliki vegetasi khas wilayah hutan gugur selatan dengan spesies pohon seperti maple ( Acer spp.), oak ( Quercus spp.), dan elm Amerika ( Ulmus americana ) serta semak dan pohon hias non-asli (Vankat, 1979 ). Detroit menunjukkan iklim kontinental sedang yang dicirikan oleh perbedaan suhu yang signifikan, yang menampilkan musim panas yang hangat hingga panas dengan suhu berkisar antara 70 hingga 90°F (21–32°C), dan musim dingin yang dingin dengan suhu berkisar antara 20 hingga 35°F (−6 hingga 2°C), dan rata-rata hujan salju sekitar 42 inci (~107 cm) (Loidi et al., 2022 ). Curah hujan relatif merata sepanjang tahun dengan jumlah yang sedikit lebih tinggi di musim semi dan musim panas yang dipengaruhi oleh kedekatannya dengan Great Lakes (Oswald et al., 2012 ).
2.2 Pengumpulan sampel eDNA dan alur kerja molekuler
Kami menyelesaikan pengambilan sampel tanah eDNA di seluruh Detroit pada bulan Februari dan Juli 2023, yang mewakili musim dingin dan musim panas. Selama pengambilan sampel tanah, eDNA dikumpulkan hingga di empat lokasi per taman dengan jumlah lokasi pengambilan sampel mencerminkan ukuran taman; misalnya, taman Palmer dan Rouge, yang lebih besar, masing-masing memiliki empat lokasi pengambilan sampel selama periode pengumpulan musim panas. Secara total, 32 sampel tanah dikumpulkan di musim dingin dan 33 di musim panas. Di setiap lokasi, kami mengumpulkan total 200 mL tanah dalam tabung sentrifus bebas RNAse 50 mL di sepanjang empat transek paralel di sekitar pohon fokus dan perangkap kamera (Gambar 2 ). Tingkat deteksi dari eDNA tanah telah terbukti lebih tinggi saat menggabungkan subsampel dalam kisi besar daripada dari satu titik unik (Andersen et al., 2012 ), dan untuk meminimalkan potensi bias terhadap taksa yang terkait dengan pohon, tanah dikumpulkan dari area yang meluas hingga 4 m dari pohon fokus. Oleh karena itu, kami mengumpulkan 10 mL tanah dari lapisan atas hingga kedalaman tanah 10 cm menggunakan sekop dan spatula dari 20 titik di sepanjang transek. Untuk meminimalkan kontaminasi, penutup sepatu sekali pakai dikenakan dari jarak sekitar 5 M dari kamera jebak untuk menghindari jejak tanah dan DNA ke area pengambilan sampel. Kami membersihkan semua peralatan sebelum setiap lokasi pengambilan sampel dengan pemutih dan kemudian membilasnya dengan air suling. Sampel disimpan dalam es di lapangan dan kemudian disimpan pada suhu -30°C hingga ekstraksi.
Kami mengekstrak DNA dari 65 sampel tanah dalam rangkap tiga, menghasilkan 195 ekstraksi eDNA, menggunakan Qiagen DNeasy Powersoil Pro Kit (Qiagen, AS). Untuk mengamplifikasi eDNA, kami menggunakan set primer MiMammal-U, yang menargetkan fragmen ~170 bp dari daerah variabel gen 12S rRNA (Ushio et al., 2017 ). Masing-masing dari 195 sampel eDNA diamplifikasi dalam rangkap tiga, dan replikasi yang dihasilkan dikumpulkan kembali untuk setiap sampel eDNA. (Kondisi dan optimalisasi PCR dirinci dalam Informasi Pendukung : Metode). Satu kontrol ekstraksi negatif dan satu kontrol PCR negatif diamplifikasi dan diurutkan dengan sampel untuk memantau potensi kontaminasi. Persiapan perpustakaan dan pengurutan ujung berpasangan (2 × 150 bp, menargetkan 2 juta pembacaan per sampel) dilakukan pada Illumina NovaSeq6000 di Yale Center for Genome Analysis.
2.3 Bioinformatika dan analisis statistik
Kami menggunakan proses multi langkah untuk menganalisis data sekuens yang dihasilkan. Pertama, kami menghapus sekuens primer dari data yang didemultipleks menggunakan cutadapt v2.3 ( Martin, 2011 ). Kami kemudian menggunakan DADA2 untuk melakukan penyaringan kualitas, menetapkan varian sekuens amplikon (ASV), menggabungkan pembacaan maju dan mundur, dan menghapus sekuens chimeric (Callahan et al., 2016 ). ASV yang terdeteksi dalam ekstraksi negatif dan kontrol PCR dihapus di semua sampel. Untuk penugasan taksonomi, kami meledakkan ASV terhadap basis data blast ‘nt’ yang telah diformat sebelumnya yang diunduh dari NCBI, yang menentukan ID taksa vertebrata, menggunakan BLAST+ v2.15 dengan kesamaan sekuens minimum 96%, dengan minimal lima sekuens basis data per ASV yang dikembalikan. Kami mengurutkan hasil BLAST berdasarkan persentase kesamaan sekuens tertinggi dan nilai-e terendah, dengan hit teratas dipilih sebagai identitas taksonomi yang paling mungkin dari setiap sekuens. Jumlah baca, yang menunjukkan jumlah sekuens yang dikaitkan dengan setiap ASV, direkam untuk analisis lebih lanjut. Kami menggabungkan sampel eDNA ke lokasi kamera, dan tingkat taman dengan menjumlahkan jumlah baca ASV yang diidentifikasi sebagai spesies (atau Genus) menggunakan paket ‘tidyverse’ (Wickham, 2019 ). Spesies yang tidak diidentifikasi sebagai mamalia, yang mencakup 16 spesies di Kelas Aves, dikeluarkan dari analisis lebih lanjut, tetapi mamalia domestik dan deteksi manusia dipertahankan. ASV manusia yang terdeteksi dalam kontrol laboratorium dikeluarkan dari semua sampel selama pemrosesan data untuk memperhitungkan potensi kontaminasi yang diperkenalkan selama pekerjaan laboratorium. Langkah ini memastikan bahwa analisis hilir mencerminkan sinyal lingkungan daripada artefak prosedural. Untuk menilai kelengkapan pengambilan sampel, kami menggunakan paket ‘iNEXT’ (Hsieh et al., 2016 ) untuk melakukan rarefaksi berdasarkan frekuensi deteksi di seluruh sampel dan menghitung interval kepercayaan 95% melalui 100 replikasi bootstrap.
Keanekaragaman gamma diselidiki dengan mengkonsolidasikan deteksi eDNA di seluruh taman dan membandingkannya dengan pengamatan mamalia tingkat penelitian di Detroit, MI, yang tercatat di iNaturalist. Kami membandingkan deteksi eDNA dengan catatan iNaturalist dari dua kerangka waktu: periode pengambilan sampel (1 Januari 2023 hingga 13 Juli 2023) dan jendela 5 tahun yang lebih luas (14 Juli 2018 hingga 13 Juli 2023). Data ada-tidaknya dikompilasi, dan uji McNemar digunakan untuk menilai perbedaan signifikan dalam probabilitas deteksi antara catatan eDNA dan iNaturalist. Menggunakan paket ‘vegan’ (Oksanen et al., 2019 ), sampel eDNA dijernihkan hingga jumlah bacaan terendah untuk menstandardisasi kekayaan spesies di berbagai upaya pengambilan sampel. Keanekaragaman alfa (kekayaan spesies) dihitung pada tingkat sampel (dengan tiga kali ekstraksi digabungkan), dan tingkat taman (dengan semua lokasi sampel di dalam taman digabungkan). Analisis varians (ANOVA) digunakan untuk menguji perbedaan kekayaan di antara taman, sementara uji Wilcoxon digunakan untuk menilai perbedaan musiman dalam keanekaragaman alfa di dalam taman.
Keanekaragaman beta dalam bentuk indeks ketidakmiripan Sørensen dihitung untuk membandingkan: deteksi eDNA dalam replikasi ekstraksi, lokasi pengambilan sampel dalam taman, dan taman dalam area studi Detroit. Kami menggunakan paket ‘betapart’ untuk menghitung ketidakmiripan Sørensen, indeks berkisar dari 0 hingga 1 dengan nilai rendah menunjukkan tumpang tindih yang tinggi dalam komposisi spesies dan 1 menunjukkan tidak ada spesies yang sama antara komunitas. Kami juga mempartisi ketidakmiripan Sørensen menjadi dua komponennya: pergantian (substitusi beberapa spesies dalam satu komunitas) dan nestedness (ketika set spesies dari satu komunitas adalah bagian dari yang lain; Baselga & Orme, 2012 ). Dendrogram klaster dibuat berdasarkan nilai keanekaragaman beta untuk memvisualisasikan perbedaan komposisi spesies di seluruh taman. Kami memvisualisasikan perubahan tingkat taman dalam komposisi dan kelimpahan komunitas di kedua musim dengan paket ‘pheatmap’ (Kolde, 2019 ). Untuk menguji pengaruh musim dan taman pada komposisi komunitas, kami menggunakan penskalaan multidimensi non-metrik (nMDS) untuk memvisualisasikan komunitas dan menguji variasi signifikan menggunakan ANOSIM dan PERMANOVA.
Kami juga menilai bagaimana karakteristik taman memengaruhi struktur komunitas mamalia. Pertama, kami menghitung proporsi pembacaan sekuensing manusia ( Homo sapiens ) per taman sebagai proksi gangguan antropogenik untuk menilai pengaruh kehadiran manusia terhadap keanekaragaman spesies. Kami mengevaluasi pengaruh kehadiran DNA manusia dan ukuran taman pada komposisi komunitas menggunakan PERMANOVA dan memvisualisasikan kontribusinya terhadap himpunan spesies dengan ordinasi nMDS. Kedua, kami menghitung koefisien korelasi antara DNA manusia dan deteksi sekuens sapi ( Bos taurus ) dan babi ( Sus scrofa ) dalam sampel eDNA untuk mengeksplorasi potensi hubungan antara kehadiran manusia dan sumber makanan antropogenik. Untuk menilai hubungan antara ukuran taman dan keanekaragaman hayati, kami menggunakan model regresi linier untuk memeriksa korelasi antara ukuran taman dan keanekaragaman alfa, ketidaksamaan Sørensen dan komponen pergantian dan keterkaitannya. Kami menggunakan paket ‘mvabund’ untuk membangun model linier umum multivariat (GLM) dan melakukan pengujian univariat untuk menentukan respons musiman khusus spesies di dalam taman. Kami menduga variasi musiman bergantung pada konteks, dipengaruhi oleh karakteristik taman dan lanskap di sekitarnya (Wang et al., 2012 ). Kami juga menduga taman yang lebih besar menampung komunitas yang lebih beragam (keanekaragaman alfa yang lebih tinggi) dan menunjukkan pergantian musiman yang lebih besar tetapi tingkat bersarang yang lebih rendah, yang mencerminkan pergeseran musiman dalam keberadaan spesies.
Terakhir, untuk menyelidiki pola ko-kemunculan spesies, kami menggunakan paket ‘cooccur’ untuk mengidentifikasi pasangan spesies dengan ko-kemunculan yang secara signifikan lebih tinggi dari yang diharapkan secara kebetulan (Griffith et al., 2016 ). Untuk analisis ini, ko-kemunculan didefinisikan di tingkat taman: pasangan spesies dianggap ko-kemunculan jika keduanya terdeteksi dalam setidaknya satu sampel dari taman yang sama selama musim tertentu. Kami membangun jaringan berbasis ada-tidaknya (untuk menghindari bias yang terkait dengan variasi amplifikasi PCR) dan menghitung metrik jaringan utama menggunakan paket ‘igraph’ untuk menganalisis perubahan musiman dalam jaringan interaksi spesies (Csárdi et al., 2024 ). Kami menghitung derajat sentralitas yang menghitung jumlah koneksi langsung (tepi) untuk setiap node dan sentralitas kedekatan sebagai kebalikan dari panjang jalur terpendek rata-rata dari satu node ke semua node lain dalam jaringan. Kami juga menghitung koefisien pengelompokan yang mencerminkan kecenderungan node untuk membentuk kelompok yang terhubung erat. Kami juga menghitung kepadatan jaringan sebagai proporsi kemungkinan koneksi yang terwujud dalam komunitas. Metrik dihitung secara terpisah untuk jaringan musim dingin dan musim panas (Martín González et al., 2010 ; Niquil et al., 2020 ). Untuk membandingkan metrik jaringan ini antar musim, pertama-tama kami menilai kenormalan menggunakan uji Shapiro–Wilk. Berdasarkan hasil ini, kami menerapkan uji Wilcoxon Signed-Rank berpasangan untuk membandingkan derajat sentralitas dan sentralitas kedekatan spesies antar musim. Semua analisis dilakukan menggunakan R versi 4.2.0 (Tim Inti R, 2021 ).
3 HASIL
3.1 Variasi antar replikasi dan sampel
Kami mengurutkan 195 sampel tanah eDNA yang dikumpulkan dari 21 taman di wilayah Detroit Metro, Michigan. Secara keseluruhan, 35,5 juta pembacaan dihasilkan, dengan rata-rata 200 ribu per sampel. Kami menemukan tingkat variasi yang tinggi di seluruh replikasi sampel dan taman, dengan indeks perbedaan Sørensen keseluruhan sebesar 0,98 di seluruh 195 sampel. Balduck Park (Gambar 2 , nomor 9), taman berukuran sedang (22,7 ha) di Detroit timur, memiliki perbedaan rata-rata terendah di antara sampel eDNA (0,55), yang menunjukkan bahwa sampel eDNA yang dikumpulkan di dalam taman ini relatif mirip dalam komposisi spesies. Sebaliknya, Marruso Park (Gambar 2 , nomor 18), taman kecil (2,2 ha) di Detroit timur laut, menunjukkan tingkat perbedaan yang tinggi (0,97), yang menunjukkan heterogenitas dalam komposisi spesies di dalam taman. Karena tingginya variasi, kami memutuskan untuk menggabungkan sampel eDNA di antara replikasi untuk memperoleh gambaran yang lebih representatif tentang keanekaragaman hayati yang ada di setiap taman.
3.2 Keanekaragaman gamma
Dari 195 sampel eDNA, 176 sampel berhasil menghasilkan sekuens DNA milik 23 spesies mamalia termasuk manusia (Tabel 1 ). Kurva akumulasi spesies menunjukkan bahwa pengambilan sampel eDNA kami secara keseluruhan mencapai cakupan pengambilan sampel 96%, yang menunjukkan efektivitas metodologi kami dalam menangkap sebagian besar keanekaragaman mamalia yang ada di taman (Gambar S1 ). Data dari sampel eDNA per lokasi pengambilan sampel digabungkan dan menyediakan data deteksi spesies dari total 65 lokasi pengambilan sampel. Khususnya, DNA manusia terdeteksi di 58 lokasi pengambilan sampel (semua taman), dan jika ada, rata-rata sepertiga dari sekuens sampel. Selain itu, sekuens dari sapi dan babi terdeteksi dalam 6 dan 2 sampel (masing-masing 5 dan 2 taman). Karena sifat spesies ini sebagai hewan pangan dan ketidakhadiran mereka sebagai kemunculan langsung, kemungkinan besar DNA tersebut berasal dari limbah makanan atau bahan feses yang dicerna, atau kedekatan dengan peternakan perkotaan tempat ternak berada yang berpotensi terjadi penularan DNA melalui angin (Clare et al., 2022 ). Kami juga mendeteksi beberapa spesies domestik lainnya; DNA anjing domestik ( Canis lupus familiaris ) terdapat pada 12 sampel (10 taman), dan kucing domestik ( Felis catus ) pada 13 sampel (11 taman). Untuk hasil terperinci perbandingan iNaturalist, silakan lihat Informasi Pendukung : dokumen.
| Memesan | Keluarga | Nama ilmiah | Nama umum | DNA elektronik | Jendela survei iNaturalist | Jendela 5 tahun iNaturalist |
|---|---|---|---|---|---|---|
| Burung pemakan daging | Bovidae | Bos Taurus | Ternak | X | ||
| Ular berbisa | Odocoileus virginianus | Rusa berekor putih | X | X | X | |
| Suidae | Tulang rusukmu | Babi | X | |||
| Karnivora | Anjing | Anjing lupus familiaris | Anjing Domestik | X | X | |
| Anjing latrans | Anjing hutan | X | X | X | ||
| Vulpes vulpes | rubah merah | X | X | |||
| Kucing | Bunga Felis Catus | Kucing Domestik | X | X | X | |
| Mephitidae | Mefitis mefitis | Sigung Bergaris | X | X | X | |
| Mustelidae | Mustela nivalis | Paling Tidak Musang | X | |||
| Mustelidae | Visi Neogale | cerpelai amerika | X | X | ||
| Procyonidae | Procyon lotor | Rakun Biasa | X | X | X | |
| Didelfimorfia | Burung pipit Didelphidae | Didelphis virginiana | Oposum Virginia | X | X | X |
| Eulipotyphla | Suku Soricidae | Blarina brevicauda (unggas) | Tikus Ekor Pendek Utara | X | X | |
| Sorex cinereus | Tikus bertopeng | X | ||||
| Burung Talpidae | Kondilura krista | Tikus Hidung Bintang | X | |||
| Scalopus akuatikus | Tahi Lalat Timur | X | X | |||
| Lagomorfa | Leporidae | Burung pipit Oryctolagus cuniculus | Kelinci Eropa | X | X | |
| Bunga Sylvilagus floridanus | Burung Kapas Timur | X | X | X | ||
| Hewan pengerat | Burung Castoridae | Tanaman jarak Kanada | Berang-berang Amerika | X | X | |
| Burung pipit | Mikrotus montanus | Tikus Pegunungan* | X | |||
| Mikrotus pennsylvanicus | Tikus Padang Rumput | X | X | |||
| Ondatra zibethicus | Tikus kesturi | X | X | X | ||
| Jamur Tiram | Tikus Kaki Putih | X | X | X | ||
| Jamur Tiram | Rusa Tikus | X | X | X | ||
| Tikus | Otot otot | Tikus Rumah | X | X | X | |
| Tikus norvegicus | Tikus Coklat | X | X | |||
| Sciuridae | Glaucomys volans | Tupai Terbang Selatan | X | X | X | |
| Ictidomys tridecemlineatus | Tupai Tanah Tiga Belas Garis | X | ||||
| Marmut monax | Babi tanah | X | X | X | ||
| Sciurus carolinensis | Tupai Abu-abu Timur | X | X | X | ||
| Ikan pari hitam | Tupai Rubah | X | X | X | ||
| Ikan pari tamias | Chipmunk Timur | X | X | X | ||
| Tamiasciurus hudsonicus | Tupai Merah Amerika | X | X | |||
| Burung Zapodidae | Zapus Hudsonius | Tikus Lompat Padang Rumput | X |
Catatan : Tikus gunung ditandai dengan tanda bintang (*) yang menunjukkan bahwa tikus tersebut tidak diketahui keberadaannya di lokasi ini.
Di seluruh area studi, 20 spesies terdeteksi di musim dingin dan 20 di musim panas; namun, ada perbedaan dalam deteksi spesies antar musim. Tikus cokelat ( Rattus norvegicus ), babi dan sekuens yang diidentifikasi pada ordo Rodentia hanya terdeteksi di musim dingin, sementara groundhog ( Marmota monax ), sigung belang dan muskrat ( Ondatra zibethicus ) hanya terdeteksi di musim panas (Tabel S2 ). Rata-rata perbedaan Sørensen untuk keanekaragaman gamma antar musim adalah 0,15, yang menunjukkan tingkat perbedaan yang rendah yang sepenuhnya disebabkan oleh pergantian. Ini menunjukkan bahwa perbedaan keanekaragaman gamma antar musim sebagian besar disebabkan oleh keberadaan spesies yang berbeda, daripada komunitas satu musim menjadi bagian dari musim lainnya. Ini sejalan dengan hipotesis kami bahwa perbedaan didorong oleh keberadaan (atau ketiadaan) spesies musiman.
3.3 Keanekaragaman alfa dan beta
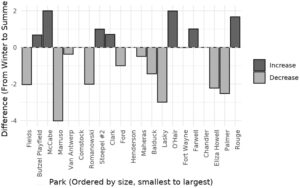
Keanekaragaman alfa yang terdeteksi di taman bervariasi secara signifikan, mulai dari 2 spesies di Butzel Playfield (taman kecil dan terisolasi seluas 1,7 ha) hingga 14 spesies yang terdeteksi di Eliza Howell Park (taman besar seluas 101,2 ha di perbatasan barat Detroit, dan bagian dari koridor hijau dengan Rouge Park) ( F (20,15) = 2,02, p < 0,001; Tabel S2 ). Analisis berpasangan keanekaragaman alfa menurut musim tidak menunjukkan perbedaan signifikan pada tingkat taman individu (uji jumlah peringkat Wilcoxon, V = 28,5, p = 0,07). Dampak musiman minimal pada keanekaragaman alfa diamati, dengan visualisasi nMDS menunjukkan perbedaan rendah antara komunitas musim dingin dan musim panas (ANOSIM −0,003, p = 0,5). Musim hanya mencakup 2% dari variasi komposisi komunitas di tingkat taman ( p = 0,5). Secara keseluruhan, taman-taman tersebut menunjukkan respons yang bervariasi dalam kekayaan spesies terhadap perubahan musiman tanpa tren konsisten yang diamati (Gambar 3 ). Demikian pula, kami mengamati respons yang bervariasi dalam keragaman beta di antara masing-masing taman antara musim dengan indeks perbedaan Sørensen berkisar antara 0,2 hingga 1 (Tabel 2 ). Henderson Park, taman seluas 14,4 ha di Detroit selatan, menunjukkan perbedaan terbesar antara musim yang sepenuhnya dikaitkan dengan pergantian spesies. Sebaliknya, Stoepel Park, taman seluas 11,9 ha di Detroit barat, memiliki perbedaan terendah antara musim yang sepenuhnya dikaitkan dengan bersarang. Dendrogram berkelompok berdasarkan total perbedaan Sørensen menyoroti variabilitas perubahan musiman untuk masing-masing taman dan mengungkapkan taman dengan komposisi spesies yang serupa (Gambar S3 ). Taman-taman dengan tutupan tanaman dan pepohonan yang secara visual lebih rapat (berdasarkan pengamatan lapangan) seperti Eliza Howell, O’Hair, Palmer dan Rouge membentuk gugusan yang jelas, sementara taman-taman yang lebih kecil dengan lebih banyak kesempatan rekreasi dan permukaan kedap air seperti Clark, Comstock dan Lasky membentuk subhimpunan yang lain.
| Titik peta | Taman | Luas taman (hektar) | Divisi musim dingin | Divisi musim panas | Tahun pembagian | SIM | SNE | MENYESAL |
|---|---|---|---|---|---|---|---|---|
| 1 | merah | 479.14 | 6.32 | 7.99 | 10 | 0,25 | angka 0 | 0,25 |
| 2 | Telapak tangan | 119.78 | 9.62 | 7.1 | 11.64 | 0,25 | 0,08 | 0.33 |
| 3 | Eliza Howell | 101.17 | Tanggal 11.21 | 8.99 | 13.99 | 0.22 | 0.11 | 0.33 |
| 4 | Pedagang lilin | 80.93 | 2 | 2 | 3 | 0.5 | angka 0 | 0.5 |
| 5 | Selamat tinggal | 36.42 | 4.01 | 5.02 | 6.17 | 0.2 | 0,07 | 0.27 |
| 6 | Benteng Wayne | 33.58 | 8 | 7.99 | 10 | 0,25 | angka 0 | 0,25 |
| 7 | Rambut O | 31.56 | 5 | 6.99 | 8 | 0.2 | 0.13 | 0.33 |
| 8 | Lasky | 26.3 | 4.99 | 2 | 5 | angka 0 | 0.42 | 0.42 |
| 9 | Botak | 22.66 | 3 | 1.56 | 3.96 | 0.5 | 0,09 | 0.6 |
| 10 | Mahera | tanggal 21.04 | 5.49 | 5 | Tanggal 8.09 | 0.4 | 0,09 | 0.5 |
| 11 | Bahasa Indonesia: Henderson | 14.44 | 1.01 | 1.01 | 2.25 | 1 | angka 0 | 1 |
| 12 | Mengarungi | 13.75 | 3 | 2 | 3 | angka 0 | 0.2 | 0.2 |
| 13 | Clark | Tanggal 12.14 | 3.29 | 4 | 5.42 | 0.5 | angka 0 | 0.5 |
| 14 | Tangga #2 | 11.85 | 2 | 3 | 3 | angka 0 | 0.2 | 0.2 |
| 15 | Bahasa Indonesia: Comstock | 10.52 | 3 | 3 | 4 | 0.33 | angka 0 | 0.33 |
| 16 | Romanowski | 10.52 | 4 | 1,99 | 5 | 0.5 | 0.16 | 0.66 |
| 17 | dari Antwerpen | 7.24 | 3.39 | 3.02 | 6.62 | 0.6 | 0,03 | 0.63 |
| 18 | Marruso | 2.18 | 4.01 | angka 0 | 4.24 | Bahasa Indonesia | Bahasa Indonesia | 1 |
| 19 | McCabe | 2.06 | 1 | 3 | 3 | angka 0 | 0.5 | 0.5 |
| 20 | Lapangan Bermain Butzel | 1.69 | 1 | 1.68 | 2 | angka 0 | 0.33 | 0.33 |
| 21 | Lapangan | 1.61 | 4.03 | 2 | 4.27 | angka 0 | 0.42 | 0.42 |
Catatan : Keragaman beta disajikan dalam bentuk indeks ketidaksamaan Sørensen (SOR) dan komponen-komponennya: perputaran (SIM) dan keberlapisan (SNE).
Untuk mengukur hubungan antara ukuran taman dan keanekaragaman, kami melakukan analisis regresi linier dengan keanekaragaman alfa total, ketidakmiripan Sørensen, dan komponen-komponennya, yaitu pergantian dan keterkaitan. Analisis regresi linier mengungkapkan korelasi positif yang signifikan antara ukuran taman dan keanekaragaman alfa total ( β = 0,0154, R 2 = 0,23, p = 0,03). Hal ini menunjukkan bahwa taman yang lebih besar cenderung menampung komunitas yang lebih beragam, konsisten dengan hipotesis kami bahwa habitat yang lebih besar menyediakan lebih banyak sumber daya dan relung untuk mendukung lebih banyak keanekaragaman spesies. Namun, kami tidak menemukan pengaruh signifikan dari ukuran taman terhadap perbedaan Sørensen ( β = −0,0006, R 2 = 0,08, p = 0,22), pergantian ( β = −0,00003, R 2 = 0,0002, p = 0,95) atau kebersarangan ( β = −0,0004, R 2 = 0,09, p = 0,20) dalam hal keragaman beta.
3.4 Komposisi komunitas dan dampak manusia
Komunitas mamalia berbeda secara signifikan antara taman ( R 2 = 0,58, p = 0,005), mengikuti harapan bahwa faktor yang berkontribusi terhadap keanekaragaman bersifat khusus untuk karakteristik taman individu dan kemungkinan atribut lingkungan sekitar. Karnivora (coyote, rakun, sigung belang dan kucing serta anjing domestik) terdeteksi di 17 dari 21 taman (Gambar S4 ). Sigung belang hanya terdeteksi selama musim panas, sementara semua karnivora lainnya terdeteksi di kedua musim. Kucing, anjing, dan rakun adalah karnivora yang paling banyak terdeteksi, sementara coyote terdeteksi hanya di tiga taman: Fort Wayne, Balduck, dan Maheras. Ukuran taman dalam penelitian ini berkisar antara 1,6 hingga 479 ha, tetapi coyote hanya terdeteksi di taman yang lebih besar dari ukuran median 14,4 ha. Taman yang lebih besar seperti Rouge, Eliza Howell, Farwell, dan Fort Wayne menunjukkan deteksi karnivora yang relatif stabil antar musim, yang mungkin mencerminkan ketersediaan habitat dan sumber daya yang lebih beragam yang memungkinkan terbentuknya wilayah jelajah. Sebaliknya, taman yang lebih kecil menunjukkan lebih banyak variabilitas dalam deteksinya.
Kami menemukan korelasi signifikan antara komposisi spesies dan proporsi DNA manusia yang ditemukan dari taman-taman ( R 2 = 0,5022, p = 0,001). Kami menggunakan peta panas untuk memvisualisasikan komposisi spesies dari masing-masing taman selama musim dingin dan musim panas dan untuk mengelompokkan taman-taman dengan komposisi spesies yang serupa (Gambar S2 ). Di musim dingin, 10 taman semuanya menunjukkan kelimpahan relatif yang tinggi dari deteksi manusia dibandingkan dengan spesies lain, dan 4 taman lainnya (O’Hair, Maheras, Fields dan Lasky) dikelompokkan karena kelimpahan relatif yang tinggi dari deteksi kucing domestik. Namun, komposisi komunitas bergeser dari prevalensi deteksi manusia yang tinggi di musim panas; meskipun Rouge, Stoepel, Van Antwerp dan Butzel tetap dikelompokkan karena deteksi ini. Deteksi anjing bertanggung jawab untuk mengelompokkan Farwell, Clark, Fields dan Romanowski, dan deteksi oposum Virginia adalah tema pemersatu di Taman Fort Wayne, Henderson dan Maheras. Kami melakukan GLM multivariat untuk setiap taman untuk membandingkan komposisi spesies di berbagai musim dan mengidentifikasi Balduck, Clark, dan Fort Wayne sebagai taman yang memiliki komposisi spesies yang berbeda secara signifikan di antara musim-musim (lihat Tabel S3 ). Pengujian univariat mengungkapkan bahwa musim memiliki efek yang signifikan pada spesies tertentu. Misalnya, deteksi manusia menurun dari musim dingin ke musim panas ( p = 0,043) di Balduck Park. Kelinci ekor kapas timur (deteksi menurun dari musim dingin ke musim panas ( p = 0,05) di Fort Wayne dan deteksi tupai rubah ( Sciurus niger ) menurun dari musim dingin ke musim panas ( p = 0,018) di Clark.
Pola ko-kemunculan spesies juga menunjukkan variasi musiman yang bernuansa (Gambar 4 ). Baik dalam jaringan musim dingin maupun musim panas, tupai abu-abu timur ( Sciurus carolinensis ) dan manusia merupakan pusat struktur jaringan, dengan nilai derajat sentralitas tertinggi (masing-masing 15 dan 13 untuk musim dingin dan musim panas). Hewan peliharaan, seperti anjing dan kucing, juga menunjukkan konektivitas yang signifikan. Di musim dingin, kucing lebih sering muncul bersamaan dengan rakun ( p = 0,001) dan rusa berekor putih ( p = 0,008) dari yang diperkirakan. Di musim panas, ko-kemunculan yang signifikan mencakup anjing dengan tikus, rusa dengan tupai rubah, dan rakun dengan tupai abu-abu ( p = 0,04 untuk masing-masing). Secara keseluruhan, derajat sentralitas rata-rata menurun dari 6,1 menjadi 4,7 (uji Wilcoxon signed-rant: p = 0,06, V = 72), yang menunjukkan bahwa spesies memiliki lebih sedikit hubungan langsung dengan spesies lain selama musim panas. Namun, tikus rumah ( Mus musculus ) dan tupai terbang ( Glaucomys volans ) menunjukkan peningkatan derajat sentralitas dari musim dingin ke musim panas (Tabel S4 ). Koefisien pengelompokan (0,9 hingga 0,86) dan kepadatan jaringan (0,32 hingga 0,25) keduanya menurun, yang mencerminkan jaringan yang lebih tersebar dan kurang saling terhubung di musim panas.
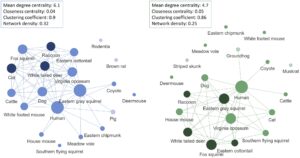
4 DISKUSI
Pemantauan satwa liar di lingkungan perkotaan semakin penting karena perubahan lingkungan membentuk kembali kesesuaian habitat dan perilaku hewan. Variasi musiman memengaruhi ketersediaan sumber daya, interaksi spesies, dan dinamika manusia-satwa liar dengan perubahan suhu, vegetasi, dan aktivitas manusia yang memengaruhi keanekaragaman hayati, dinamika penyakit, dan ketahanan ekosistem perkotaan. Studi kami menyoroti perubahan musiman dalam komunitas mamalia di kota besar di wilayah tengah barat AS, yang menawarkan wawasan untuk perencanaan dan pengelolaan perkotaan. Memahami dinamika temporal ini sangat penting karena memengaruhi fungsi ekosistem, ketahanan keanekaragaman hayati, dan koeksistensi manusia-satwa liar. Temuan kami menggarisbawahi perlunya strategi pengelolaan adaptif yang memperhitungkan perubahan musiman dalam keanekaragaman mamalia.
4.1 Variasi musiman dalam keanekaragaman mamalia
Pergeseran musiman dalam komposisi spesies terlihat jelas, dengan indeks perbedaan Sørensen yang tinggi (0,98) yang diamati di seluruh replikasi, yang menunjukkan variabilitas spasial yang signifikan dalam deteksi spesies, yang mungkin disebabkan oleh pergerakan lokal, preferensi habitat, atau perbedaan tingkat aktivitas antar musim. Sementara desain pengambilan sampel kami, yang mengumpulkan tanah dari beberapa titik subsampel, bertujuan untuk mengurangi variabilitas ini, desain tersebut tidak dapat sepenuhnya menjelaskan pengaruh perilaku hewan dan heterogenitas habitat pada pola deteksi. Menyadari variabilitas ini, kami menafsirkan temuan kami dengan hati-hati dan menekankan pentingnya replikasi dan metodologi standar dalam mengurangi bias pengambilan sampel (Buxton et al., 2021 ; Shirazi et al., 2021 ). Selain itu, pergantian spesies, bukan bersarang, adalah pendorong utama perbedaan musiman, yang menunjukkan bahwa banyak spesies terdeteksi hanya dalam satu musim. Meskipun hal ini mungkin mencerminkan variasi musiman yang sebenarnya dalam hunian dan aktivitas spesies, persistensi DNA dalam tanah juga dapat memengaruhi deteksi (Guthrie et al., 2024 ). DNA yang disimpan di musim dingin mungkin tetap dapat dideteksi hingga musim panas, yang berpotensi meningkatkan estimasi ko-kemunculan spesies antar musim dan kami mengakui bahwa DNA lama, terutama di iklim yang lebih dingin seperti Detroit, mungkin masih berkontribusi pada perbedaan yang diamati dalam komposisi komunitas (Guthrie et al., 2024 ; Valentin et al., 2021 ). Penelitian lebih lanjut yang menggabungkan interval pengambilan sampel yang lebih pendek atau eksperimen degradasi dapat membantu mengukur persistensi DNA dan meningkatkan resolusi perbandingan musiman dalam sistem ini.
Meskipun ada keterbatasan ini, studi kami mencapai cakupan pengambilan sampel 96%, yang menunjukkan bahwa metodologi kami secara efektif menangkap sebagian besar keanekaragaman mamalia yang ada. Pergeseran musiman yang diamati dalam komunitas mamalia selaras dengan pola yang didokumentasikan di lingkungan perkotaan lainnya, di mana faktor-faktor seperti suhu, distribusi sumber daya, dan aktivitas manusia memengaruhi keberadaan dan pergerakan satwa liar. Misalnya, subsidi antropogenik, seperti limbah makanan dan tempat makan burung dapat mengurangi fluktuasi musiman dalam ketersediaan sumber daya, mengubah pola alami migrasi, hibernasi, dan perilaku mencari makan (Crandall et al., 2024 ). Demikian pula, rekreasi manusia sering kali lebih sering terjadi pada bulan-bulan yang lebih hangat, yang dapat menyebabkan pergeseran dalam distribusi spesies, terutama untuk spesies yang sensitif terhadap gangguan (Marion et al., 2020 ). Dinamika musiman ini menggarisbawahi perlunya pemantauan jangka panjang untuk mengantisipasi perubahan keanekaragaman hayati di bawah skenario iklim dan penggunaan lahan di masa depan. Memahami pola-pola ini sangat penting untuk merancang ruang hijau perkotaan yang mendukung satwa liar dan mengurangi hilangnya keanekaragaman hayati. Selain itu, temuan kami memiliki implikasi untuk penularan penyakit zoonosis, karena pergeseran dalam distribusi spesies dapat memengaruhi interaksi patogen-inang, sebuah perhatian utama dalam kerangka kerja Satu Kesehatan (Gilbertson et al., 2023 ).
4.2 Luas taman dan keanekaragaman mamalia
Komposisi komunitas di taman kota dapat menunjukkan perubahan yang halus dan spesifik taman, seperti yang diamati dalam penelitian kami. Pergeseran musiman dalam suhu, ketersediaan makanan dan siklus perkembangbiakan, dikombinasikan dengan faktor-faktor lokal seperti kepadatan vegetasi, ketersediaan air dan keanekaragaman habitat, menciptakan pola siklus yang memengaruhi komposisi spesies dan menyebabkan variasi dalam kehadiran dan kelimpahan antara musim dingin dan musim panas (Johnson & Swan, 2014 ; Threlfall et al., 2017 ). Kami mengamati penurunan koefisien pengelompokan dan kepadatan jaringan yang menunjukkan komunitas mamalia yang lebih tersebar dan kurang saling berhubungan selama musim panas. Pergeseran ini kemungkinan mencerminkan peningkatan ketersediaan sumber daya dan keanekaragaman habitat, yang memungkinkan penyebaran spesies yang lebih luas dan mengurangi pembentukan kelompok yang saling berhubungan erat. Selain itu, taman di daerah kota yang lebih urban sering kali mengalami tingkat polusi, kebisingan, dan gangguan manusia yang lebih tinggi yang dapat memengaruhi komunitas satwa liar (Zhou & Chu, 2012 ). Namun, taman dengan habitat alami yang lebih luas dapat mengurangi dampak ini dengan menyediakan lebih banyak tempat berlindung bagi spesies yang sensitif (Beasley et al., 2023 ). Taman yang lebih besar khususnya menyediakan habitat yang lebih beragam, yang sangat penting untuk mempertahankan kekayaan spesies yang lebih tinggi dibandingkan dengan ruang hijau yang lebih kecil dan lebih terfragmentasi (Beninde et al., 2015 ). Misalnya, anjing hutan terdeteksi hanya di taman yang lebih besar dari ukuran rata-rata 14,4 ha, yang mungkin mencerminkan preferensi mereka terhadap wilayah yang luas dengan sumber daya yang melimpah. Sementara pengambilan sampel tanah kami difokuskan pada area di sekitar pohon fokus, potensi bias terhadap taksa yang terkait dengan pohon diperkirakan minimal. Mamalia besar melepaskan DNA secara luas di seluruh jangkauannya melalui feses, urin, sel kulit, air liur, dan rambut, yang menyebabkan akumulasi DNA di berbagai substrat dan mengurangi pengaruh mikrohabitat tertentu seperti pohon (Leempoel et al., 2020 ). Selain itu, habitat yang diambil sampelnya menunjukkan fitur lingkungan yang bervariasi, termasuk vegetasi semak belukar, yang berkontribusi pada heterogenitas habitat. Taman Eliza Howell, yang dicirikan oleh ukurannya yang besar, vegetasi yang lebat, dan konektivitas ke ruang hijau lainnya, menunjukkan kekayaan spesies yang lebih besar dan struktur komunitas yang berbeda dibandingkan dengan taman yang lebih terisolasi seperti Butzel Playfield. Temuan ini menggarisbawahi perlunya mempertimbangkan variabel ekologi, konektivitas, dan tekanan perkotaan saat mengelola ruang hijau perkotaan untuk mendukung keanekaragaman satwa liar. Taman yang lebih besar memainkan peran penting dalam menjaga keanekaragaman hayati perkotaan dengan menawarkan tempat perlindungan penting bagi satwa liar di lanskap yang terfragmentasi (Zellmer & Goto, 2022)). Hal ini menyoroti pentingnya kebijakan perencanaan kota yang memprioritaskan koridor habitat dan infrastruktur hijau untuk meningkatkan ketahanan spesies. Seiring berlanjutnya urbanisasi, mengintegrasikan desain yang peka terhadap keanekaragaman hayati ke dalam perencanaan kota dapat membantu mengurangi hilangnya habitat dan mendorong koeksistensi spesies.
4.3 Pengaruh manusia terhadap komposisi komunitas
Taman kota berfungsi sebagai ruang rekreasi bagi manusia dan habitat penting bagi satwa liar. Gangguan manusia seperti rekreasi, jalan-jalan anjing, dan membuang sampah sembarangan dapat berdampak negatif pada satwa liar, membentuk kembali komposisi komunitas dengan mengutamakan spesies generalis sambil menggantikan taksa yang lebih sensitif (Beasley et al., 2023 ). Pergeseran ini memiliki konsekuensi ekologis, berpotensi mengganggu interaksi trofik, dinamika penyakit, dan stabilitas keanekaragaman hayati dalam ekosistem perkotaan (Gámez & Harris, 2021 ; Lima et al., 2021 ). DNA manusia adalah urutan yang paling sering terdeteksi di semua taman, yang mencerminkan aktivitas manusia yang signifikan dan interaksi dengan ekosistem satwa liar perkotaan. Taman mengalami lalu lintas pejalan kaki yang tinggi dengan pengunjung yang membawa DNA melalui sampah makanan, produk hewani, dan kotoran (Darling et al., 2021 ). Aktivitas seperti piknik, barbekyu, dan pertemuan rekreasi dapat berkontribusi pada pengenalan produk daging olahan ke lingkungan (Crandall et al., 2024 ). Bahasa Indonesia: Sementara pemasakan dapat mendegradasi DNA, penelitian telah menunjukkan bahwa fragmen DNA yang terdeteksi dapat bertahan bahkan dalam produk daging yang diproses secara berlebihan (Shokralla et al., 2015 ). Deteksi sekuens sapi dan babi kemungkinan berasal dari kotoran manusia atau hewan peliharaan atau satwa liar yang mencari makan dari sumber makanan antropogenik—terutama di musim dingin ketika sumber daya alam langka (Henger et al., 2022 ). Meskipun eDNA manusia tetap dapat dideteksi untuk jangka waktu yang lama, itu belum banyak digunakan sebagai proksi untuk urbanisasi (Antony Dass et al., 2022 ). Perlu dicatat bahwa DNA manusia dalam studi eDNA menimbulkan pertimbangan etika dan hukum yang penting, termasuk masalah privasi dan persetujuan, seperti yang disorot dalam diskusi baru-baru ini (Doi & Kelly, 2023 ). Dalam studi kami, kami memastikan bahwa DNA manusia diperlakukan secara generik, membatasi analisis pada identifikasi tingkat spesies untuk menghindari potensi masalah etika atau pelanggaran privasi. Lebih jauh, kami menyadari perlunya pedoman internasional yang jelas untuk mengarahkan penggunaan eDNA manusia secara bertanggung jawab, menyeimbangkan manfaat penelitian eDNA dengan penghormatan terhadap prinsip-prinsip etika. Sementara kontrol kontaminasi yang ketat telah diterapkan—seperti menghilangkan ASV manusia yang terdeteksi dalam kontrol lab—tetap sulit untuk sepenuhnya memisahkan sinyal yang berasal dari manusia dari kontaminasi. Interpretasi kami berfokus pada eDNA manusia sebagai indikator ekologis dampak antropogenik, menghindari kesimpulan tingkat individu yang sejalan dengan pedoman etika. Di samping manusia, spesies domestik seperti anjing dan kucing memainkan peran sentral dalam jaringan komunitas, memengaruhi struktur dan konektivitas interaksi spesies (Herrera et al., 2022 ; Hughes & Macdonald, 2013 ; Ünal et al., 2019)). Taman dengan aktivitas manusia yang lebih tinggi dapat menarik spesies generalis yang mudah beradaptasi, seperti rakun dan tupai, sekaligus menghalangi spesies yang lebih sensitif terhadap gangguan (Parker & Nilon, 2012 ; Suraci et al., 2021 ). Temuan kami tentang korelasi positif yang signifikan antara komposisi spesies dan proporsi DNA manusia di taman menunjukkan bahwa gangguan manusia memperkuat pergeseran dalam struktur komunitas, yang konsisten dengan penelitian sebelumnya tentang dinamika satwa liar perkotaan (Gámez & Harris, 2021 ). Memahami peran spesies ini memberikan wawasan berharga tentang ketahanan dan stabilitas komunitas satwa liar perkotaan. Sementara penelitian kami berfokus pada mamalia, menggabungkan data dari taksa lain—seperti burung dan invertebrata—akan menawarkan gambaran yang lebih komprehensif tentang jaring makanan perkotaan, keanekaragaman hayati, dan kesehatan ekosistem.
4.4 Keterbatasan penelitian
Meskipun penelitian kami memberikan wawasan berharga tentang keanekaragaman mamalia perkotaan dan dinamika musiman, faktor metodologi dan ekologi tertentu menimbulkan keterbatasan yang harus dipertimbangkan. Kami mengumpulkan data selama satu musim lapangan, yang membatasi kekokohan perbandingan musim dingin vs. musim panas. Variabilitas antartahunan dalam kondisi cuaca, ketersediaan pangan, dan dinamika populasi dapat memengaruhi tren musiman, yang menyoroti perlunya kumpulan data multi-tahun untuk menilai konsistensi pola-pola ini dan lebih memahami dinamika musiman jangka panjang. Meskipun penelitian ini memberikan wawasan berharga tentang keanekaragaman mamalia perkotaan, perbedaan metodologi antara eDNA dan iNaturalist menyoroti bias yang melekat. Deteksi observasional bergantung pada upaya pengamat, yang dapat menjelaskan tidak adanya spesies umum seperti anjing domestik dalam catatan iNaturalist meskipun keberadaannya diketahui di taman. Demikian pula, tidak adanya pengamatan tikus cokelat dalam data iNaturalist dapat disebabkan oleh sifatnya yang samar atau statusnya sebagai spesies yang terabaikan dan ada di mana-mana. Tidak adanya 10 spesies dalam deteksi eDNA menyoroti peluang untuk penyempurnaan metodologis. Kesalahan identifikasi tikus gunung sebagai tikus padang rumput, spesies yang diharapkan di area studi, kemungkinan besar disebabkan oleh kesamaan genetik dan resolusi lokus yang diperkuat yang tidak memadai. Penanda genetik tambahan dapat meningkatkan spesifisitas spesies dan meminimalkan kesalahan identifikasi lintas spesies (Weitemier et al., 2021 ). Tantangan lain dalam studi ini adalah diferensiasi antara urutan anjing hutan dan anjing domestik (Reese et al., 2020 ). Perilaku khusus spesies dan penggunaan habitat kemungkinan memengaruhi perbedaan antara pola deteksi eDNA dan catatan iNaturalist. Gaya hidup bawah tanah tikus mol timur kemungkinan membatasi pengendapan DNA di tanah permukaan, sementara berang-berang Amerika, yang sebagian besar akuatik, tidak mungkin meninggalkan DNA yang dapat dideteksi di tanah terestrial. Deteksi kelinci Eropa pada iNaturalist didasarkan pada penampakan tunggal, sehingga keberadaan DNA-nya tidak mungkin. Sementara eDNA dapat menangkap sinyal sementara, seperti dari sapi dan babi, iNaturalist bergantung pada upaya pengamat, yang berpotensi kurang mewakili spesies yang tersembunyi. Penelitian di masa mendatang dapat memadukan metode pelengkap, seperti pemasangan kamera penjebakan, guna memvalidasi deteksi dan menilai konektivitas lanskap pada komunitas satwa liar perkotaan.
4.5 Arah masa depan dan kesimpulan
Memahami komposisi dan dinamika komunitas satwa liar perkotaan sangat penting untuk perencanaan konservasi yang efektif (Des Roches et al., 2021 ). Kehadiran mamalia di daerah perkotaan sering kali menyebabkan konflik manusia-satwa liar, termasuk kematian di jalan raya, kerusakan properti, interaksi langsung dengan manusia dan hewan peliharaan, serta potensi risiko zoonosis (Santini et al., 2019 ). Kehadiran spesies inti yang stabil sepanjang tahun, ditambah dengan aktivitas manusia, menyoroti perlunya strategi pengelolaan yang ditargetkan yang mengurangi konflik dan mendorong koeksistensi (Klees van Bommel et al., 2022 ). Strategi seperti menciptakan koridor satwa liar, meningkatkan kompleksitas habitat, dan menerapkan program pendidikan publik dapat meningkatkan kesadaran dan mendorong koeksistensi antara penduduk perkotaan dan satwa liar (Basak et al., 2023 ). Ruang hijau perkotaan menyediakan habitat penting bagi satwa liar, memfasilitasi interaksi manusia-alam, dan mendorong minat konservasi (Basak et al., 2022 ). Namun, peran wilayah perkotaan dalam menghubungkan atau memecah ekosistem dan fungsinya sebagai reservoir keanekaragaman hayati atau perangkap ekologi perlu diselidiki lebih lanjut (Schnetler et al., 2021 ; Zuñiga-Palacios et al., 2021 ). Pemantauan jangka panjang populasi mamalia perkotaan dapat membantu mengidentifikasi periode kritis ketika spesies paling rentan terhadap konflik manusia-satwa liar sekaligus menjelaskan bagaimana spesies individu beradaptasi dengan lingkungan perkotaan. Seiring dengan terus berkembangnya kota, mengintegrasikan eDNA ke dalam program pemantauan keanekaragaman hayati dapat menjadi sangat penting untuk melestarikan satwa liar perkotaan dan menjaga keseimbangan ekologi (Donald et al., 2021 ; Webster et al., 2020 ). Pemahaman yang lebih mendalam tentang dinamika ini akan menginformasikan strategi konservasi dan perencanaan perkotaan, memastikan bahwa kebutuhan satwa liar dan manusia terpenuhi.