
Abstrak
- Pada komunitas tumbuhan dengan mosaik semak-rumput, jenis utama serasah meliputi serasah herba dan bentuk campurannya dengan daun dan batang semak. Akan tetapi, hubungan kuantitatif antara jenis komposisi serasah dan dekomposisi serasah, serta bagaimana proses ini dipengaruhi oleh aktivitas penggembalaan (misalnya pengendapan kotoran), sebagian besar masih belum dieksplorasi.
- Di sini, percobaan inkubasi serasah selama tiga tahun dilakukan di padang rumput Alpen yang ditumbuhi semak belukar. Kami bertujuan untuk menjelaskan efek individual dan interaktif dari jenis serasah (hanya serasah rumput (L), campuran serasah rumput dan daun semak belukar (L1) dan campuran serasah rumput, daun semak belukar dan batang semak belukar (L2)) dan penambahan kotoran yak (tanpa penambahan, hanya penambahan kotoran, hanya penambahan urin, penambahan kotoran dan urin) pada laju dekomposisi serasah dan pelepasan nutrisi. Selain itu, kualitas kimia awal serasah, iklim mikro tanah, nutrisi tanah dan atribut mikroba tanah dianalisis untuk memastikan mekanisme pendorong yang mendasarinya.
- Kami menemukan bahwa jenis serasah dan kotoran yak memiliki efek individual dan interaktif yang signifikan pada dekomposisi serasah. Secara khusus, jenis serasah terutama memengaruhi dekomposisi serasah dengan meningkatkan konsentrasi karbon dan lignin awal dan mengurangi konsentrasi nitrogen dan fosfor pada serasah. Sebaliknya, kotoran yak berdampak positif pada dekomposisi serasah melalui peningkatan pH tanah dan aktivitas mikroba tanah. Yang terpenting, serasah berkualitas rendah (L2) mengurangi efek positif kotoran yak pada dekomposisi serasah. Sementara penambahan kotoran yak tidak mengubah arah keseluruhan dekomposisi serasah, itu memengaruhi besarnya efek yang disebabkan oleh pencampuran serasah. Khususnya, imobilisasi nitrogen sementara diamati selama tahap awal dekomposisi, dan pola pelepasan fosfor bersih terlihat jelas di seluruh proses dekomposisi. Namun, durasi imobilisasi nitrogen berbeda tergantung pada jenis serasah dan perawatan kotoran yak.
- Temuan kami memberikan bukti lokal yang menjelaskan efek interaktif jenis serasah dan penambahan kotoran yak pada proses dekomposisi serasah dan pelepasan nutrisi. Hasil ini berkontribusi pada pemahaman yang lebih mendalam tentang bagaimana aktivitas penggembalaan memodulasi siklus nutrisi dalam ekosistem padang rumput yang diserbu semak belukar.
Baca Ringkasan Bahasa Sederhana gratis untuk artikel ini di blog Jurnal.
1. PENDAHULUAN
Dekomposisi serasah merupakan proses ekologi yang krusial di hutan, padang rumput, dan banyak ekosistem lainnya (Joly et al., 2023 ). Memang, sebagian besar karbon dan energi yang mengalir melalui ekosistem ini berasal dari degradasi serasah tanaman (Zhang et al., 2023 ). Serasah tanaman ini memicu jaring makanan ‘coklat’, yang dicirikan oleh mikroba heterotrofik dan detritivora yang menggunakan dan mentransfer bahan organik mati ke konsumen tingkat tinggi (Pigani et al., 2022 ). Secara global, variasi yang diamati dalam dekomposisi bahan organik tanaman dijelaskan sekitar 70% oleh kombinasi kondisi lingkungan bersama dengan kualitas dan kuantitas serasah (Canessa et al., 2021) . ).
Bahasa Indonesia: Di bawah kondisi lingkungan yang konsisten, dekomposisi serasah terutama diatur oleh sifat fisikokimia awal serasah dan keanekaragaman spesies serasah (Meier & Bowman, 2008 ). Misalnya, banyak penelitian telah menunjukkan bahwa serasah dengan konsentrasi nutrisi yang tinggi, seperti nitrogen (N), fosfor (P), kalsium (Ca) dan magnesium (Mg), cenderung terurai lebih cepat karena sifatnya yang diinginkan oleh pengurai (García-Palacios et al., 2016 ). Baru-baru ini, sifat fisik seperti luas daun spesifik (SLA), massa kering daun dan konsentrasi nutrisi, yang mencerminkan sifat struktural jaringan, telah terbukti berkorelasi dengan laju dekomposisi serasah (Helsen et al., 2020 ). Selain itu, kualitas serasah awal tidak hanya memengaruhi hilangnya massa serasah tetapi juga menentukan laju imobilisasi dan mineralisasi nutrisi (Ni et al., 2018 ; Scott & Binkley, 1997 ). Misalnya, derajat imobilisasi nitrogen bersih sering dikaitkan dengan konsentrasi nitrogen awal dari serasah yang membusuk. Ketika nitrogen terbatas, N dapat ditransfer dari tanah ke serasah melalui hifa jamur, yang menyebabkan peningkatan konsentrasi nitrogen dari serasah yang membusuk (Parton et al., 2007 ). Song et al. ( 2017 ) mengungkapkan bahwa rasio C: N awal rata-rata serasah daun Leymus chinensis melampaui 40. Selanjutnya, serasah daun memasuki fase sementara imobilisasi nitrogen. Saat dekomposisi berlangsung dan rasio C: N turun ke kisaran 25–40, mineralisasi nitrogen dimulai, yang mengakibatkan pelepasan bentuk nitrogen anorganik. Fosfor, yang sangat mobil dan kurang mudah diserap oleh organisme, menunjukkan pelepasannya dari serasah terutama didorong oleh kualitas awal dan aktivitas mikroba tanah (Gijsman et al., 1997 ; Pan et al., 2024 ). Misalnya, Liao et al. ( 2023 ) mengamati bahwa morfologi dan kualitas serasah memainkan peran penting dalam hilangnya fosfor dari serasah daun selama tahap pertengahan hingga akhir dan dari serasah akar selama tahap awal hingga pertengahan.
Bahasa Indonesia: Setelah semak-semak merambah ke padang rumput, komposisi komunitas tumbuhan di lanskap ini berubah, sehingga mengubah kualitas serasah tumbuhan yang memasuki ekosistem (Aguirre et al., 2021 ; Ding & Eldridge, 2023 ). Perbedaan massa substrat antara serasah semak dan serasah rumput dapat menyebabkan perbedaan dalam dekomposisi serasah, yang dipengaruhi oleh efek langsung dari kualitas awal yang bervariasi dan efek tidak langsung dari interaksi pencampuran serasah (Poulette & Arthur, 2012 ). Misalnya, di Pyrenees, serasah dari dua spesies semak ( Juniperus communis dan Cytisus balansae ) mengandung konsentrasi tinggi senyawa yang membandel (misalnya lignin, suberin dan lipid) dan konsentrasi rendah N atau P relatif terhadap serasah rumput. Serasah rumput terurai lebih cepat daripada serasah jenis semak, dan ketika serasah rumput dan semak hidup berdampingan, laju degradasi kedua serasah semak meningkat dibandingkan ketika mereka terurai secara independen selama masa inkubasi (Montané et al., 2010 ). Bukti eksperimental dari padang rumput semi-gurun di Arizona menemukan bahwa serasah mesquite beludru ( Prosopis velutina ) terurai lebih cepat daripada serasah lovegrass, yang dikaitkan dengan rasio C: N awal yang lebih tinggi untuk mesquite (15,7 ± 0,1) dibandingkan dengan serasah lovegrass (45,0 ± 0,1) (Throop & Archer, 2007 ). Memang, perambahan semak telah terjadi di padang rumput Alpen di Dataran Tinggi Tibet karena pemanasan iklim dan penggembalaan berlebihan (Zhang et al., 2023 ). Spesies semak seperti Potentilla fruticosa , Salix cupularis , dan Spiraea alpina telah menjadi spesies dominan, menggantikan alang-alang ( Kobresia spp.) dan tumbuhan perdu (Wang, Michalet, et al., 2023 ). Perubahan komposisi spesies ini kemungkinan akan memengaruhi laju dekomposisi serasah dan, akibatnya, penyimpanan karbon ekosistem. Namun, hubungan antara proses dekomposisi serasah dan perubahan komposisi serasah tanaman (misalnya batang semak, daun semak) masih kurang dipahami.
Efek jenis serasah pada tingkat dekomposisi dimodulasi bersama oleh aktivitas penggembalaan (Duan et al., 2013 ). Penggembalaan memengaruhi dekomposisi serasah melalui tiga cara utama, yaitu pengguguran daun oleh hewan (herbivora), penginjakan, dan pengembalian nutrisi melalui ekskresi (Huang & Hou, 2024 ). Ternak secara substansial mengurangi biomassa pucuk tanaman melalui penggembalaan dan penjelajahan, dan mereka mengembalikan sekitar 60%–90% nutrisi yang tertelan ke tanah dalam bentuk urin dan kotoran (Cheng et al., 2016 ). Di Dataran Tinggi Tibet, distribusi ekskresi tidak merata karena perilaku makan dan istirahat yak yang bervariasi (Cai et al., 2013 ). Kotoran ini sering ditemukan pada atau tercampur dengan serasah tanaman, mengalami dekomposisi secara bersamaan dari waktu ke waktu. Penelitian sebelumnya telah menunjukkan bahwa kotoran yang kaya nutrisi mempercepat dekomposisi serasah tanaman (Cai et al., 2013 ). Mekanisme teoritis utamanya adalah: (i) transformasi hara dari kotoran berkualitas tinggi ke serasah berkualitas rendah melalui pemindahan atau pelindian mikroba (Wang, Li, et al., 2023 ); (ii) peningkatan komposisi lapisan serasah karena efek komplementaritas fauna tanah dan pengurai (Georgiou et al., 2017 ); dan (iii) perbaikan kondisi lingkungan mikro melalui perubahan pH tanah, kadar air dan suhu (Cheng et al., 2016 ). Misalnya, penambahan kotoran sapi mempercepat degradasi karbohidrat struktural serasah tanaman (misalnya hemiselulosa, selulosa dan lignin), yang dikaitkan dengan nitrogen ekstra dalam aktivitas bakteri yang ditingkatkan kotoran (Wang, Li, et al., 2023 ). Deposisi kotoran yak tidak memiliki dampak signifikan pada pembusukan lignin karena efek pengimbang dari dampak negatif pada fotodegradasi dan dampak positif pada aktivitas mikroba (Liang et al., 2018 ). Sebaliknya, penelitian tentang interaksi antara urin dan serasah tanaman selama proses dekomposisi masih terbatas. Aplikasi urin menghasilkan kondisi yang sedikit asam (kisaran pH 5,2 hingga 5,8) di area yang ditanami, yang mendukung pertumbuhan bakteri (Cai et al., 2017 ). Urin sintetis yang mengandung natrium (Na) meningkatkan median dekomposisi kayu hampir dua kali lipat dibandingkan dengan urin-Na dan plot kontrol dalam sistem detrital di Ekuador (Clay et al., 2015 ). Sebagian besar penelitian saat ini berfokus pada proses dekomposisi independen dari kotoran ternak dan serasah tanaman (Liang et al., 2018). ), sementara interaksi antara kedua proses ini dalam ekosistem pegunungan yang beralih dari padang rumput ke semak belukar sebagian besar masih belum dieksplorasi.
Tujuan dari penelitian kami ada dua: (i) untuk menjelaskan efek dan mekanisme pendorong yang mendasari jenis serasah dan kotoran yak, dan interaksinya, pada dekomposisi serasah; dan (ii) untuk menentukan pola pelepasan konsentrasi N dan P serasah dan membandingkan perbedaan ini. Kami berhipotesis bahwa (i) jenis serasah tanaman dan kotoran yak memiliki efek individual dan interaktif yang signifikan pada laju dekomposisi serasah. Serasah batang semak berkualitas rendah melemahkan dampak positif kotoran yak pada dekomposisi serasah. Mekanisme yang dengannya jenis serasah dan kotoran yak memengaruhi dekomposisi serasah berbeda. Jenis serasah terutama memengaruhi kualitas serasah awal, sedangkan kotoran yak terutama mengatur iklim mikro tanah, nutrisi tanah, dan aktivitas mikroba tanah (Gambar 1 ). (ii) Pola pelepasan konsentrasi N dan P berbeda secara signifikan di bawah perlakuan yang berbeda. Dalam kasus konsentrasi N, ada imobilisasi sementara pada tahap dekomposisi awal; sedangkan untuk konsentrasi P, ada pola pelepasan bersih selama periode inkubasi.
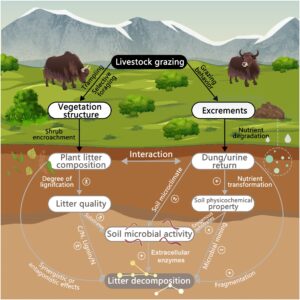
2 BAHAN DAN METODE
2.1 Deskripsi situs
Percobaan ini dilakukan di Stasiun Pengamatan dan Penelitian Nasional Ekosistem Lahan Basah Alpen Sichuan Zoige, yang terletak sekitar 4 km di utara Kabupaten Hongyuan, Provinsi Sichuan, Tiongkok (lintang 32°48′ LU, bujur 102°33′ BT, ketinggian sekitar 3500 m di atas permukaan laut). Suhu rata-rata di area tersebut adalah 1,5°C, berkisar antara -9,7°C pada bulan Januari hingga 11,1°C pada bulan Juli. Curah hujan tahunan rata-rata adalah 747 mm, dengan lebih dari 70% jatuh selama musim hangat yang pendek (Gambar S1 ). Vegetasi lokal dikategorikan sebagai padang rumput Alpen yang diserobot oleh semak belukar, terutama terdiri dari alang-alang seperti Kobresia cristata (L.) Pers., rumput termasuk Elymus nutans Griseb., serta forb dominan seperti spesies Saussurea , di samping semak belukar yang didominasi oleh Potentilla fruticosa L. Tanah di daerah tersebut diklasifikasikan sebagai Mat-Cryic Cambisols menurut taksonomi WRB (IUSS Working Group WRB, 2015 ). Wilayah tersebut terutama dikelola untuk penggembalaan yak, tetapi penggembalaan telah dikecualikan selama 6 tahun sebelum melakukan percobaan pada tahun 2017. Sifat-sifat tanah di lokasi penempatan serasah konsisten. Horizon tanah permukaan (0–10 cm) dicirikan oleh 6,51 ± 0,45% karbon total, 0,46 ± 0,03% nitrogen total, 0,03 ± 0,001% fosfor total dan pH tanah 5,32 ± 0,16. Percobaan dekomposisi serasah dilakukan pada bulan Oktober 2017 sampai dengan Desember 2019, dengan total waktu inkubasi selama 660 hari.
2.2 Percobaan Dekomposisi Serasah
2.2.1 Penyiapan serasah tanaman
Melalui pemantauan plot tertentu, ditentukan bahwa komposisi serasah di padang rumput Alpen yang ditumbuhi semak terutama terdiri dari serasah rumput, daun semak, dan batang semak. Pada bulan Oktober 2017, serasah rumput yang tumbang, daun semak, dan batang semak dikumpulkan dari lapangan dan diangkut ke laboratorium. Selanjutnya, serasah yang dikumpulkan segera disterilkan pada suhu 121°C selama 20 menit (Liao et al., 2023 ; Strickland et al., 2009 ). Langkah sterilisasi ini dilakukan untuk memastikan bahwa sumber komunitas mikroba yang paling mungkin dalam sistem eksperimen adalah tanah, bukan serasah itu sendiri. Akhirnya, semua sampel serasah yang dikumpulkan dikeringkan dalam oven pada suhu 60°C selama 24 jam di laboratorium. Sampel yang dikeringkan kemudian dipotong-potong sekitar 2 cm menggunakan gunting dan dihomogenkan dengan hati-hati untuk meminimalkan ‘efek ukuran’ (Bärlocher et al., 2020 ). Serasah tanaman kemudian dibagi menjadi dua subsampel, dengan yang pertama digunakan untuk analisis kimia serasah awal dan yang kedua disimpan dalam kantong kertas pada suhu ruangan untuk percobaan dekomposisi serasah. Dalam penelitian ini, tiga perlakuan jenis serasah adalah hanya serasah rumput (L), campuran serasah rumput dan daun semak (L1) dan campuran serasah rumput, daun semak dan batang semak (L2). Secara khusus, L1 terdiri dari serasah rumput dan daun semak dalam rasio 3:2, sedangkan L2 terdiri dari serasah rumput, daun semak dan batang semak dalam rasio 3:1:1, menyerupai rasio yang diamati di lapangan.
2.2.2 Persiapan kotoran yak
Kami melakukan pengamatan dan mencatat perilaku ekskresi yak selama enam hari berturut-turut sebelum melakukan percobaan dekomposisi serasah. Aktivitas yang dicatat meliputi berat kotoran harian (dalam kilogram) dan volume urin (dalam liter), serta ukuran petak untuk kotoran dan urin. Setiap yak menghasilkan rata-rata 5–6 tumpukan kotoran segar per hari, dengan setiap tumpukan berbobot sekitar 1 kg dan menempati area sekitar 0,04 m 2 . Setiap yak juga mengeluarkan rata-rata 8,5 liter urin per hari, dengan setiap buang air kecil sekitar 2 L dan menutupi area sekitar 0,20 m 2 .
Kotoran yak (kotoran segar dan urin) dikumpulkan dari yak betina dewasa di area perkemahan yang berdekatan dengan lokasi pengambilan sampel tanah. Yak dikurung di dalam perkemahan pada malam hari, dan sampel kotoran segar mereka dikumpulkan dalam ember plastik keesokan paginya hingga jumlah yang dibutuhkan untuk percobaan diperoleh. Total kotoran yang dikumpulkan dari hewan kemudian dicampur dengan hati-hati dan disimpan sebelum digunakan. Kotoran yak segar mengandung 370,10 g kg −1 total karbon dan 6,9 g kg −1 total nitrogen. Selain itu, sampel urin segar dikumpulkan secara manual (di bawah ekor yak oleh penjaga) ke dalam ember plastik keesokan paginya. Total urin kemudian dicampur dan dipindahkan ke wadah tertutup, yang ditempatkan dalam pendingin dengan bungkusan es dan diangkut ke laboratorium dalam waktu 2 hari. Sampel disimpan pada suhu −20°C selama sekitar 15 hari untuk mencegah hilangnya nitrogen. Sebelum digunakan, urin beku dicairkan pada suhu 4°C dan disaring dengan hati-hati melalui kertas saring untuk menghilangkan endapan. Urine yak segar mengandung 106,20 g kg −1 total karbon dan 13,1 g kg −1 total nitrogen.
2.2.3 Percobaan dekomposisi serasah sawah
Sebelum memulai percobaan penguraian serasah, petak penelitian (20 m × 10 m) ditutup dengan pagar untuk mencegah yak yang merumput merusak kantong serasah di semak belukar pegunungan Alpen. Semua bagian di atas tanah, termasuk batang herba tegak, serasah tanah (serasah rumput, daun semak dan batang semak, dll.), disingkirkan rata dengan tanah.
Untuk menilai dekomposisi serasah di lapangan, digunakan teknik kantong serasah. Kantong serasah yang digunakan berukuran 15 cm × 15 cm dan terbuat dari kain kasa nilon 2 mm. Ukuran kasa ini dipilih untuk menghindari hilangnya fragmen serasah yang lebih kecil selama periode inkubasi di lapangan, sekaligus tetap memungkinkan akses ke serasah oleh mikrofauna dan mesofauna tanah, tetapi membatasi akses ke makrofauna (Vossbrinck et al., 1979 ).
Berdasarkan survei sebelumnya mengenai biomassa serasah di atas tanah per satuan luas, setiap kantong serasah diisi dengan 10 g serasah campuran yang dikeringkan dengan udara untuk memastikan bahwa lingkungan mikro di dalam kantong serasah tersebut mirip dengan lingkungan alami dan memiliki cukup serasah sisa untuk pengukuran pada waktu pengambilan sampel terakhir. Kantong serasah diberi jarak setidaknya 20 cm satu sama lain dan diletakkan secara acak di permukaan tanah petak. Untuk memastikan homogenitas perlakuan penambahan kotoran, kotoran yak segar dan urin yang telah dikumpulkan dan dibekukan masing-masing dicampur secara menyeluruh. Selanjutnya, campuran tersebut disebarkan secara merata di atas kantong serasah pada hari ketika serasah diletakkan di petak ladang. Perlakuan kotoran yak meliputi penambahan kotoran saja (D), penambahan urin saja (U), penambahan kotoran dan urin (DU), dengan perlakuan kontrol (CK) yang tidak menerima penambahan kotoran. Bercak-bercak kotoran dan urin yang disimulasikan ini mirip dengan volume dan ukuran kotoran yak yang secara alami diendapkan di ladang. Secara khusus, untuk perlakuan D, setiap tumpukan kotoran yang ditambahkan memiliki berat sekitar 1 kg, diameter sekitar 15 cm, dan ketebalan sekitar 5 cm. Untuk perlakuan U, 0,5 L urin yak segar ditaburkan secara merata di atas setiap kantong serasah. Perlakuan DU melibatkan penambahan urin ke dalam perlakuan kotoran. Sebanyak 288 kantong serasah dibuat, yang terdiri dari tiga jenis serasah tanaman, empat jenis perlakuan kotoran yak, empat ulangan, dan enam waktu pengambilan. Selama periode percobaan, perkecambahan dan pertumbuhan tanaman selanjutnya yang berasal dari bank benih di tanah atau kotoran yak dipantau dan segera dibuang.
2.3 Pengumpulan dan analisis sampel serasah dan tanah
2.3.1 Pengumpulan sampel serasah dan tanah
Kantong serasah diambil pada hari ke-60, 180, 300, 420, 540 dan 660 masa inkubasi, dengan 48 kantong diambil pada setiap acara pengambilan sampel. Setelah diambil, kantong serasah dibersihkan dengan cermat untuk menghilangkan kotoran, tanah halus dan bentuk kontaminasi lainnya seperti pasir dan tanah bangkai hewan, dan kemudian dikeringkan pada suhu 65°C selama 48 jam sebelum ditimbang untuk menentukan massa kering yang tersisa. Sisa serasah digiling hingga 0,05 mm menggunakan ball milling dan kemudian disimpan untuk analisis kimia. Suhu tanah dan kadar air tanah di bawah setiap kantong serasah pada kedalaman 10 cm diukur menggunakan Field-Scout TDR-100 (Spectrum Technologies, Plainfield, IL, AS). Sampel tanah dikumpulkan menggunakan auger tanah pada kedalaman 0–10 cm di bawah setiap kantong serasah. Tiga inti tanah berdiameter 5 cm dicampur bersama untuk membuat satu sampel dari setiap plot. Selanjutnya, tanah komposit dibagi menjadi tiga bagian: satu bagian dikeringkan dengan udara untuk pengukuran fisikokimia tanah selanjutnya. Sub-sampel kedua disimpan pada suhu 4°C untuk pengukuran selanjutnya karbon dan nitrogen biomassa mikroba tanah. Bagian terakhir diawetkan pada suhu -80°C untuk ekstraksi DNA selanjutnya, amplifikasi PCR dan sequencing Illumina Miseq untuk membandingkan keragaman komunitas mikroba di antara berbagai perlakuan (Caporasoa et al., 2011 ).
2.3.2 Analisis kimia
Konsentrasi total karbon dan total nitrogen dari serasah dan sampel tanah ditentukan menggunakan alat analisis elemen (Flash EA1112, Thermo Scientific, West Palm Beach, FL, AS). Total kandungan fosfor dari serasah dan sampel tanah dianalisis menggunakan alat analisis injeksi aliran FIAstar 5000 (Foss Tecator, Högnäs, Swedia). pH tanah diukur menggunakan meteran pH DMP-2 mV (Quark Ltd., Nanjing, Cina) dengan rasio tanah kering terhadap air 1:2,5.
Konsentrasi serat deterjen netral (NDF%) dan serat deterjen asam (ADF%) dari serasah tanaman diukur melalui ANKOM 2000 Automated Fiber Analyzer (A2000i, Fiber Analyzer, Amerika), mengikuti metodologi yang dijelaskan oleh Van Soest et al. ( 1991 ). Pencernaan lignin deterjen asam (ADL%) dalam larutan H 2 SO 4 72% , yang diperoleh dengan metode lignin sulfat, digunakan untuk menganalisis kandungan lignin dari fraksi ADF (Trofymow et al., 2002 ). Konsentrasi hemiselulosa dihitung sebagai NDF% − ADF%.
Konsentrasi karbon (MBC) dan nitrogen (MBN) biomassa mikroba di dalam tanah diukur menggunakan metode fumigasi-ekstraksi kloroform yang dimodifikasi (Setia et al., 2012 ). Keragaman komunitas bakteri dan jamur dievaluasi melalui polymerase chain reaction denaturing gradient gel electrophoresis (PCR-DGGE) yang menargetkan gen 16S rRNA dan 18S rRNA, masing-masing. Dinamika keragaman mikroba selama dekomposisi serasah dinilai menggunakan indeks keragaman Shannon-Wiener dan indeks keragaman Chao1.
2.4 Pernyataan replikasi
| Skala inferensi | Skala penerapan faktor minat | Jumlah replikasi pada skala yang sesuai |
|---|---|---|
| Jenis serasah | Merencanakan | 3 |
| Jenis kotoran | Merencanakan | 4 |
| Serasah tanaman | Merencanakan | 24 |
| Tanah | Merencanakan | 24 |
2.5 Perhitungan numerik
Persamaan yang digunakan untuk menentukan massa serasah yang tersisa dan konstanta dekomposisi serasah ( k ) dari waktu ke waktu adalah sebagai berikut (Olson, 1963 ):
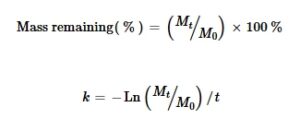
Dalam persamaan ini, k merupakan konstanta dekomposisi. M 0 merupakan massa kering awal, M t merupakan massa kering pada titik waktu tertentu ( t ), dan t merupakan waktu dekomposisi serasah yang diukur dalam hari (Swift et al., 1979) . ).
Sisa serasah hemiselulosa, lignin, N dan P digabungkan menggunakan persamaan berikut (García-Palacios et al., 2016 ):
![]()
Pada persamaan ini, RCt (%) merupakan representasi dari hemiselulosa, lignin, N dan P yang tersisa pada serasah. M 0 merupakan representasi dari massa kering awal serasah, M t merupakan representasi dari massa kering serasah pada setiap waktu pengambilan sampel, C 0 merupakan representasi dari konsentrasi awal hemiselulosa, lignin, N dan P sebagai persentase dari massa serasah sebelum inkubasi, dan C t melambangkan konsentrasi unsur-unsur ini sebagai persentase massa serasah pada setiap kejadian pengambilan sampel.
Laju pelepasan hemiselulosa, lignin, N dan P selama periode dekomposisi sebelumnya pada setiap peristiwa pengambilan sampel diperkirakan menggunakan rasio massa kering elemen yang hilang terhadap massa kering awal (Song et al., 2017) . ).
Massa nutrisi serasah pada setiap waktu pengambilan sampel dihitung dengan persamaan berikut:
![]()
Laju pembusukan hemiselulosa, lignin, N, P dan S dihitung dengan persamaan berikut:
![]()
Dimana Laju pelepasan hara melambangkan laju peluruhan hemiselulosa, lignin, N, P, dan S, NM sebelum dan NM sesudah masing-masing melambangkan massa hara sebelum dan setelah penguraian serasah, dan t adalah waktu penguraian serasah yang diukur dalam hari.
2.6 Analisis statistik
Semua analisis dilakukan menggunakan R versi 4.4.1 dan RStudio 2024.04.2 dengan paket relevan yang dikutip di bawah ini. Semua gambar dibuat menggunakan Origin 2024b (Origin Lab Corporation, Northampton, Massachusetts, AS) dan paket ‘ ggplot2 ‘. Sebelum melakukan analisis, semua data diuji kenormalannya menggunakan uji kesesuaian Shapiro–Wilk dan homogenitas varians menggunakan uji Levene. Generalized linear mixed modelling (GLMM) dengan paket glmmTMB digunakan (Brooks et al., 2017 ; Nooten et al., 2024 ) untuk memeriksa dampak jenis serasah tanaman, kotoran yak, dan interaksinya terhadap massa sisa bahan kering serasah tanaman, serta hemiselulosa, lignin, N, dan P yang tersisa dari serasah. Dalam model ini, jenis serasah tanaman dan kotoran yak dimasukkan sebagai efek tetap, sedangkan waktu pengambilan sampel diperlakukan sebagai efek acak. Pada setiap waktu pengambilan sampel, kami melaksanakan analisis varians dua arah (ANOVA) untuk mengevaluasi pengaruh jenis serasah tanaman, kotoran yak, dan interaksinya terhadap massa residu DM serasah tanaman, beserta hemiselulosa, lignin, N, dan P yang tersisa.
ANOVA satu arah digunakan untuk menguji pengaruh kotoran yak terhadap massa sisa bahan kering serasah, konstanta dekomposisi serasah Δ, laju pelepasan nutrisi serasah Δ, sifat tanah dan indeks keanekaragaman mikroba tanah untuk setiap jenis serasah tanaman pada setiap waktu pengambilan sampel. Uji Tukey’s HSD dalam kemasan ‘ agricolae ‘ dilakukan untuk mengevaluasi perbedaan di antara rata-rata, dengan tingkat signifikansi ditetapkan pada p < 0,05.
Analisis regresi linier disesuaikan untuk menguji hubungan statistik antara massa serasah yang tersisa dan N dan P serasah yang tersisa. Analisis korelasi Pearson dilakukan dengan menggunakan paket ‘ corrplot ‘ untuk mengeksplorasi hubungan antara hilangnya serasah (konstanta dekomposisi, laju pelepasan hemiselulosa serasah, lignin, N dan P) dan faktor lingkungan. Analisis hutan acak, yang dilakukan dengan menggunakan paket ‘ rfPermute ‘, mengidentifikasi prediktor yang paling berpengaruh dari iklim mikro tanah, nutrisi tanah dan atribut mikroba tanah pada laju dekomposisi serasah, laju pelepasan N serasah dan laju pelepasan P serasah. Persentase peningkatan dalam kesalahan kuadrat rata-rata (%MSE) variabel digunakan untuk menentukan signifikansi prediktor potensial berdasarkan analisis hutan acak.
Pemodelan persamaan struktural (SEM) digunakan untuk mengevaluasi efek langsung dan tidak langsung dari jenis serasah dan kotoran yak pada dekomposisi serasah. Dalam model apriori terstruktur (Gambar S2 ), kualitas serasah awal, iklim mikro tanah dan nutrisi tanah diperkenalkan sebagai pendorong dekomposisi utama, dan mikroba tanah digunakan sebagai pendorong utama. Variabel-variabel ini dipilih karena secara tradisional dianggap sebagai faktor pengendali yang dominan (Hättenschwiler et al., 2005 ). Kualitas serasah awal menunjukkan efek warisan dari perbedaan awal dalam kimia serasah di antara spesies yang berbeda, termasuk total C, lignin, N, P dan rasio C: N, lignin: N. Iklim mikro tanah mengacu pada kondisi iklim dalam lapisan tanah di bawah permukaan, khususnya yang melibatkan suhu tanah dan kelembaban tanah. Nutrisi tanah menunjukkan zat nutrisi yang ada di dalam tanah, dan aktivitas mikroba tanah menunjukkan keragaman mikroorganisme tanah (bakteri dan jamur) yang menjajah serasah selama dekomposisi. Mengingat korelasi yang kuat di antara faktor-faktor dalam setiap kelompok, kami melakukan analisis komponen utama (PCA) untuk membangun indeks fungsional multivariat sebelum menerapkan prosedur SEM (Chen et al., 2019 ; Veen et al., 2010 ). Selanjutnya, komponen pertama (PC1), yang mencakup 56,8% dan 50,9% dari total varians dalam kualitas serasah awal dan mikroba tanah masing-masing, dimasukkan sebagai variabel baru yang mewakili karakteristik kelompok gabungan ke dalam analisis selanjutnya (Tabel S3 ; Gambar S12 ). Koefisien jalur dan nilai p terkaitnya diperkirakan menggunakan metode kemungkinan maksimum. Uji chi-kuadrat dan nilai p terkaitnya digunakan untuk menyesuaikan model (kesesuaian yang baik ketika 0 ≤ χ 2 ≤ 2 dan 0,05 < p ≤ 1,00). Kesalahan rata-rata kuadrat akar dari perkiraan (RMSEA: kesesuaian yang baik ketika 0 ≤ RMSEA ≤ 0,05) dan indeks kesesuaian komparatif (CFI; kesesuaian model yang baik ketika 0,97 ≤ CFI ≤ 1) digunakan untuk mengevaluasi kesesuaian model. Analisis SEM dilakukan menggunakan IBM SPSS Amos 26.0 (Chicago, IL: Amos Development Corporation).
3 HASIL
3.1 Kualitas kimia awal serasah tanaman
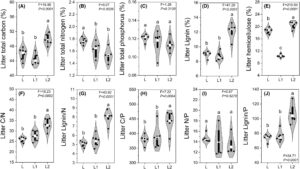
Komposisi kimia awal serasah sangat bervariasi antara jenis serasah tanaman (Gambar 2 ). Konsentrasi komponen struktural (yaitu C dan lignin) di kedua jenis serasah L dan L1 secara signifikan ( p < 0,05) lebih rendah daripada yang ada di jenis serasah L2 (Gambar 2A,E,F ). Sebaliknya, jenis serasah L2 mengandung konsentrasi komponen nutrisi yang lebih rendah (yaitu N dan P) daripada jenis serasah L dan L1 (Gambar 2B,C ). Selain itu, jenis serasah L1 memiliki konsentrasi lignin, hemiselulosa, dan rasio Lignin:P terendah (Gambar 2D,E,I ).
3.2 Perubahan konstanta dekomposisi serasah dan laju pelepasan nutrisi
Baik jenis serasah tanaman maupun kotoran yak secara signifikan memengaruhi massa sisa bahan kering serasah dan hemiselulosa, lignin, N, dan P serasah yang tersisa (Tabel S1 ). Selain itu, selama tahap dekomposisi awal, terdapat efek interaktif yang signifikan antara jenis serasah tanaman dan kotoran yak pada indikator serasah ini, kecuali untuk N serasah yang tersisa (Tabel S1 ).
Bahasa Indonesia: Saat menganalisis efek jenis serasah saja, kami menemukan bahwa dibandingkan dengan jenis serasah L tanpa mempedulikan perlakuan kotoran yak, jenis serasah L1 mendorong konstanta dekomposisi serasah; sebaliknya, jenis serasah L2 mengurangi konstanta dekomposisi serasah (Gambar S3a ). Secara rinci, nilai perubahan (Δ) konstanta dekomposisi serasah meningkat sebesar 17,0%, 33,7% dan 21,26% dengan jenis serasah L1 di bawah perlakuan CK, D dan DU, berturut-turut (Gambar S3a ). Nilai perubahan (Δ) konstanta dekomposisi serasah menurun sebesar 7,7%, 10,9%, 31,9% dan 6,2% dengan jenis serasah L2 di bawah perlakuan CK, D, U dan DU, berturut-turut (Gambar S3a ). Selain itu, laju pelepasan lignin Δ litter dan laju pelepasan N Δ litter dengan tipe litter L1 masing-masing adalah 2,7% dan 2,3%, dan laju pelepasan N dengan tipe litter L2 masing-masing adalah -1,1% dan -0,1% pada perlakuan D dan DU (Gambar S3c ). Laju pelepasan N Δ litter dengan tipe litter L1 masing-masing adalah 5,5% dan 2,1%, dan laju pelepasan N dengan tipe litter L2 masing-masing adalah -7,8% dan -3,8% pada perlakuan D dan DU (Gambar S3e ).
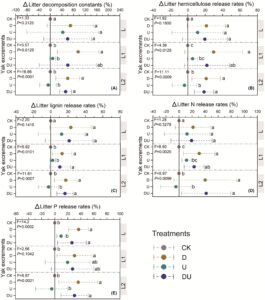
Bahasa Indonesia: Saat menganalisis efek kotoran yak saja, kami menemukan bahwa konstanta dekomposisi serasah dan laju pelepasan nutrisi di bawah perlakuan D dan DU semuanya lebih tinggi daripada yang di bawah perlakuan tanpa penambahan kotoran yak (CK) terlepas dari jenis serasah (Gambar 3 ; Gambar S5 dan S6 ). Dibandingkan dengan perlakuan CK, nilai perubahan (Δ) konstanta dekomposisi serasah (K) meningkat sebesar 69,2%, 95,9% dan 60,3% dengan penambahan D dan sebesar 59,7%, 60,4% dan 53,0% dengan penambahan DU pada perlakuan jenis serasah L, L1 dan L2, masing-masing (Gambar 3A ). Untuk tipe serasah L1 dan L2, bukan tipe serasah L, terdapat perbedaan signifikan ( p < 0,05) dalam konstanta dekomposisi serasah Δ, laju pelepasan hemiselulosa serasah Δ, laju pelepasan lignin serasah Δ dan laju pelepasan N serasah Δ (Gambar 3 ). Secara keseluruhan, penambahan urin yak (perlakuan U) tidak mengubah nilai konstanta dekomposisi serasah Δ dan laju pelepasan nutrisi serasah Δ secara signifikan ( p > 0,05) dibandingkan dengan perlakuan CK (Gambar 3 ).
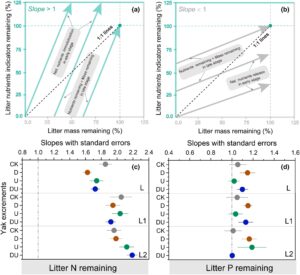
3.3 Perbandingan kemiringan persamaan yang disesuaikan antara massa serasah yang tersisa dan N dan P serasah yang tersisa
Laju pelepasan dan pola N dan P bervariasi secara signifikan di bawah perlakuan yang berbeda (Gambar 4 ; Gambar S6 ). Selama tahap awal dekomposisi (0–300 hari), imobilisasi nitrogen sementara diamati pada serasah L2 dan semua kombinasi ekskresi (Gambar S6 ). Untuk serasah L dan L1, imobilisasi N hanya terjadi pada perlakuan CK dan DU selama periode inkubasi 0–180 hari (Gambar S6 ). Serasah kehilangan N secara proporsional lebih sedikit daripada massa total, dengan kemiringan lebih besar dari 1,0 untuk hubungan ini, yang tidak bervariasi di antara perlakuan serasah dan ekskresi (Gambar 4c ). Sebaliknya, ada pola pelepasan bersih dari total konsentrasi P dari waktu ke waktu untuk semua kombinasi serasah dan ekskresi (Gambar S6 ). Serasah kehilangan P secara proporsional lebih sedikit daripada massa total, menghasilkan kemiringan lebih tinggi dari 1,0 untuk hubungan ini di bawah perlakuan D (Gambar 4d ).
3.4 Faktor-faktor yang mempengaruhi konstanta dekomposisi serasah dan laju pelepasan nutrisi
Konstanta dekomposisi serasah dan laju pelepasan P serasah berkorelasi positif dengan konsentrasi N dan P awal dan rasio N: P awal, namun berkorelasi negatif dengan kandungan karbon awal, kandungan lignin, dan rasio C: N dan lignin: N awal (Gambar S4). ).
Perubahan faktor tanah yang diakibatkan oleh penambahan kotoran yak juga memengaruhi hilangnya massa serasah dan pelepasan nutrisi, tetapi polanya berbeda antara jenis serasah (Gambar 5 ; Gambar S11 ). Secara khusus, untuk jenis serasah L, analisis hutan acak menunjukkan bahwa TC tanah, TN, TP, pH, MBC, indeks Shannon–Weiner jamur tanah, dan kelembapan tanah adalah variabel terpenting untuk hilangnya massa serasah dan laju pelepasan nutrisi (N dan P), dan ada korelasi positif yang signifikan di antara mereka (Gambar S11a,d–f ). TN tanah, MBC, pH tanah dan keanekaragaman bakteri tanah diamati untuk menjelaskan sebagian besar variasi dalam dekomposisi serasah untuk jenis serasah L1, dengan korelasi positif yang kuat (Gambar S11b,g–i ). Namun, untuk jenis serasah L2, tidak ada korelasi signifikan ( p > 0,05) yang diamati antara konstanta massa serasah (K), laju pelepasan nutrisi, dan sifat tanah (kecuali untuk SM; p < 0,05; Gambar S11c,j–l ).
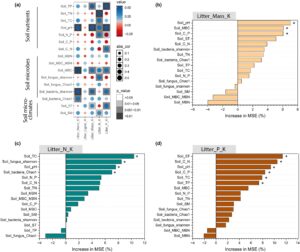
Hasil dari SEM menunjukkan bahwa 55% varians dalam laju dekomposisi serasah dapat dijelaskan (Gambar 6a ). Jenis serasah memiliki dampak signifikan pada dekomposisi serasah dengan mengubah kualitas awal serasah. Ini dicapai dengan meningkatkan konsentrasi C dan lignin dan mengurangi konsentrasi N dan P dari serasah awal (Gambar 6a ). Jenis serasah memiliki efek negatif tidak langsung dengan menurunkan suhu tanah karena kualitas serasah awal (Gambar 6a ). Efek total terstandarisasi dari jenis serasah pada dekomposisi serasah adalah -0,44 (Gambar 6b ). Kotoran yak memiliki efek positif tidak langsung pada dekomposisi serasah dengan meningkatkan aktivitas mikroba tanah (Gambar 6a ), dengan efek langsung terstandarisasi sebesar 0,34 (Gambar 6b ). Selain itu, kotoran yak mempunyai efek positif tidak langsung dengan meningkatkan konsentrasi TP tanah dan pH tanah (Gambar 6a ), dengan efek langsung terstandarisasi masing-masing sebesar 0,27 dan 0,33 (Gambar 6b ).

4 DISKUSI
4.1 Efek individual jenis serasah dan kotoran yak terhadap dekomposisi serasah
Dalam komunitas tanaman dengan mosaik semak-rumput, jenis serasah utama meliputi serasah herba dan bentuk campuran yang terdiri dari daun dan batang semak. Namun, saat ini belum jelas bagaimana komposisi serasah ini memengaruhi dekomposisi serasah dan mekanisme yang mendasarinya. Hasil kami menunjukkan bahwa, dibandingkan dengan jenis serasah L (terlepas dari perlakuan kotoran yak), jenis serasah L1 secara signifikan mendorong konstanta dekomposisi serasah (Gambar S3a ). Peningkatan ini dapat dikaitkan dengan konsentrasi nitrogen dan fosfor yang tinggi, serta rasio C:N yang rendah dan konsentrasi komponen struktural (seperti karbon, dan lignin) pada daun semak, yang secara kolektif mempercepat dekomposisi serasah (Gambar 2 ; Gambar S4 ). Penelitian sebelumnya telah menetapkan bahwa peningkatan nitrogen dalam campuran serasah dapat meningkatkan ketersediaan nutrisi bagi komunitas mikroba, yang mengarah pada efek sinergis pada mineralisasi N tanah dan dekomposisi serasah (García-Palacios et al., 2016 ; Porre et al., 2020 ). Dalam penelitian kami, ketika daun semak dan serasah rumput dicampur, serasah kaya nutrisi (daun semak) dapat memicu efek priming pada serasah miskin nutrisi, memfasilitasi dekomposisi lebih cepat dari serasah ini tanpa harus memperlambat dekomposisi serasah berkualitas tinggi (Gambar 2 ; Gambar S4 ). Sarabi dkk. ( 2021 ) menguatkan temuan ini, menunjukkan bahwa peningkatan spesies tanaman dengan kualitas serasah yang lebih tinggi berdampak positif pada proses dekomposisi serasah dan pelepasan nutrisi berikutnya dari campuran serasah di hutan ek Mediterania semiarid. Sebaliknya, temuan kami mengungkapkan bahwa jenis serasah L2 mengurangi konstanta laju dekomposisi serasah sebesar 14,18% (efek antagonis) (Gambar S3 ). Efek antagonis tersebut biasanya diamati ketika serasah miskin nutrisi memperkuat imobilisasi nutrisi mikroba atau ketika senyawa sekunder penghambat dilepaskan oleh spesies serasah berkualitas rendah (Liu dkk., 2020 ). Dalam penelitian kami, kami menemukan bahwa serasah batang semak mengandung konsentrasi tinggi senyawa yang sulit dihilangkan, termasuk lignin, lignin/N dan lignin/P, ditambah dengan konsentrasi N atau P yang relatif rendah dibandingkan dengan serasah rumput (Gambar S2 ). Kehadiran lebih banyak batang berkayu yang terkait dengan semak dapat menandakan peningkatan jumlah C yang tersimpan dalam puing dan batang berkayu kasar, yang membusuk pada tingkat yang lebih lambat. Lebih jauh, tampaknya kadar senyawa yang larut dalam air lebih rendah pada batang semak daripada pada serasah daun yang sesuai (Hobbie, 1996 ). Temuan ini didukung oleh penelitian sebelumnya. Misalnya, di tundra asam yang lembap, semak peluruh Betula nanamengalokasikan 79% dari total biomassa tahunan di atas tanah ke batang baru dan lama (Shaver et al., 2006 ).), yang terurai tiga kali lebih lambat daripada daun dan satu hingga delapan kali lebih lambat daripada daun dan batang dari rumput dan semak cemara (Hobbie, 1996 ). Di sabana Australia, kandungan lignin yang tinggi pada batang semak A. gayanus mengakibatkan laju dekomposisi serasah campuran 50% lebih lambat dibandingkan dengan serasah rumput saja (Rossiter-Rachor et al., 2017) .
Sementara banyak penelitian telah difokuskan pada karakteristik dekomposisi kotoran ternak dan efek individualnya pada dekomposisi serasah (Wang, Li, dkk., 2023 ), terdapat kekurangan penelitian tentang dampak urin dan campuran kotoran dan urin pada proses yang sama. Namun, dalam kasus di mana tekanan penggembalaan tinggi, kotoran dan urin sering kali diendapkan pada petak padang rumput yang sama secara bersamaan, dan kotoran dan urin hewan betina sering kali tercampur. Dalam penelitian ini, hasil kami menunjukkan bahwa, ketika mempertimbangkan tiga jenis serasah secara komprehensif, perlakuan D dan DU menyebabkan peningkatan konstanta dekomposisi serasah masing-masing sebesar 75,1% dan 57,7% (Gambar 3 ). Sebaliknya, hanya menambahkan urin yak tidak secara signifikan mengubah nilai konstanta dekomposisi serasah Δ dan laju pelepasan nutrisi serasah Δ, terutama pada tahap dekomposisi akhir (Gambar 3 ). Oleh karena itu, penelitian menunjukkan bahwa kontribusi kotoran yak terhadap dekomposisi serasah secara signifikan lebih besar daripada urin yak. Efek positif kotoran pada dekomposisi serasah kemungkinan besar disebabkan oleh peningkatan kelembapan tanah dan aktivitas mikroba (Wang, Li, et al., 2023 ). Sebuah studi baru-baru ini melaporkan bahwa bakteri, bukan jamur, ditemukan terkait dengan degradasi hemiselulosa dan selulosa; kotoran mendukung pertumbuhan bakteri (aktivitas metabolisme dan respirasi spesifik yang lebih tinggi) daripada jamur (Weverka et al., 2023 ). Selain itu, pengendapan kotoran dapat mengurangi efek fotodegradasi dengan memberikan perisai terhadap sinar matahari, tetapi pada akhirnya meningkatkan biodegradasi, sehingga menghasilkan efek minimal pada hilangnya lignin (Cheng et al., 2022 ). Dalam studi ini, analisis SEM menunjukkan bahwa kotoran yak memiliki efek positif tidak langsung pada dekomposisi serasah dengan meningkatkan pH tanah dan aktivitas mikroba tanah (Gambar 6 ; Gambar S9–S11 ). Sementara itu, kadar air tanah juga ditemukan meningkat sampai batas tertentu dengan penambahan kotoran yak (Gambar S8 ). Urine, yang kaya akan natrium, dapat mempercepat degradasi selulosa dan lignin dalam serasah. Misalnya, menambahkan urine sintetis yang mengandung natrium ke dalam sistem detrital di Ekuador menghasilkan peningkatan hampir dua kali lipat dalam dekomposisi kayu rata-rata dibandingkan dengan plot kontrol dan urine-natrium (Clay et al., 2015 ). Hasil kami menunjukkan bahwa penambahan urine yak secara signifikan memengaruhi dekomposisi serasah hanya selama tahap awal (Gambar S5 ). Hal ini dapat dikaitkan dengan hidrolisis nitrogen yang cepat dalam urine menjadi nitrogen terlarut, yang mudah menguap dan mungkin tidak menyediakan nutrisi yang cukup bagi mikroorganisme pendegradasi lignin selama tahap dekomposisi selanjutnya (Shen et al., 2023) . ).15 Kotoran urin rusa kutub yang diberi label N, Barthelemy et al. ( 2018 ) menemukan bahwa nitrogen dari urin dapat dengan cepat diserap oleh bagian tanaman di atas tanah dalam waktu 2 minggu, tetapi rasio penyerapan oleh mikroorganisme tanah dan akar tanaman relatif rendah.
4.2 Efek interaktif jenis serasah dan kotoran yak terhadap dekomposisi serasah
Berdasarkan hasil kami, ada efek interaktif yang signifikan antara jenis serasah tanaman dan kotoran yak pada dekomposisi serasah selama tahap pembusukan awal (Tabel S1 , Gambar S5 dan S6 ). Efek positif penambahan kotoran yak pada dekomposisi serasah menunjukkan variabilitas di antara berbagai jenis serasah (Gambar 3 ; Gambar S3 ). Serasah batang semak berkualitas rendah melemahkan efek positif kotoran yak pada dekomposisi serasah (Gambar 3 ; Gambar S5 dan S6 ). Atenuasi ini dapat dikaitkan dengan ketidakmampuan serasah berkualitas rendah untuk memenuhi kebutuhan energi dan nutrisi mikroorganisme secara memadai selama dekomposisi, sehingga memengaruhi aktivitas mikroba (Bai et al., 2024 ). Misalnya, Song et al. ( 2020 ) melaporkan bahwa aktivitas mikroba tanah dan kadar air tanah lebih rendah di komunitas lokal yang miskin spesies daripada di komunitas lokal yang kaya spesies. Demikian pula, komunitas tanaman berkualitas rendah menyumbang lebih sedikit masukan bahan organik, yang memperburuk persaingan di antara komunitas mikroba tanah dan berpotensi mengakibatkan penurunan aktivitas mikroba tanah. Hasil SEM kami selanjutnya menunjukkan bahwa pengaturan dekomposisi serasah dengan penambahan kotoran yak terutama dimediasi melalui perubahan pH tanah, TP tanah, dan aktivitas mikroba tanah (Gambar 6) ).
Dalam penelitian ini, penambahan kotoran yak memberikan efek signifikan pada dekomposisi serasah dan pelepasan nutrisi, sekaligus mengubah efek campuran dekomposisi serasah. Efeknya bergantung pada tahap dekomposisi (Gambar S3 ; Tabel S1 ). Selama tahap awal dekomposisi, penambahan kotoran yak mempertahankan lingkungan lembap yang bermanfaat bagi aktivitas dan pertumbuhan hifa jamur (Palomino et al., 2023 ). Peningkatan aktivitas jamur lebih sensitif terhadap gradien nutrisi, memfasilitasi transfer nutrisi positif antara berbagai jenis serasah dan secara efektif mendorong penurunan berat badan pada serasah berkualitas rendah. Saat dekomposisi berlangsung, kadar air dalam kotoran yak menurun karena dekomposisinya sendiri, yang mengakibatkan penurunan kapasitasnya untuk transfer nutrisi. Selain itu, ketersediaan nutrisi yang dipercepat yang difasilitasi oleh kotoran yak membuat sifat-sifat serasah dengan kualitas berbeda menjadi lebih mirip, yang menyebabkan hilangnya efek positif transfer nutrisi. Secara bersamaan, karbon yang larut dalam air dalam kotoran yak meringankan keterbatasan metabolik dari serasah berkualitas rendah, secara efektif merangsang aktivitas pengurai mikroba dan menghasilkan enzim ekstraseluler yang kaya, khususnya dalam kondisi yang dicirikan oleh rasio C: N dan lignin: N yang tinggi (Liang et al., 2018 ). Hasil kami menunjukkan bahwa pengendapan kotoran yak meningkatkan kadar air dan keanekaragaman jamur tanah dalam lingkungan dekomposisi (Gambar S7 dan S10 ). Hal ini mendorong jamur tanah untuk secara efisien memanfaatkan serasah berkualitas rendah untuk mendapatkan karbon guna mengimbangi keterbatasan energinya. Selain itu, pengendapan kotoran yak dapat menarik fauna tanah, seperti kumbang kotoran, dan mendorong perkembangan larva Hymenoptera , yang selanjutnya mempercepat dekomposisi (Joly et al., 2023 ; Peng et al., 2023 ). Secara keseluruhan, peran kotoran yak dalam memediasi efek campuran dekomposisi serasah menunjukkan bahwa penentu utama efek pencampuran serasah adalah kualitas serasah, dan penambahan kotoran tidak dapat mengubah arahnya tetapi hanya memengaruhi besarnya efek pencampuran. Yang penting, penambahan kotoran yak mengubah aktivitas mikroba (Gambar S9 dan S10 ), membuatnya lebih sensitif terhadap perbedaan kimia antarspesies, yang memperkuat interaksi spesies dan memodifikasi efek campuran dekomposisi serasah. Ini selanjutnya menyoroti peran penting pengendapan kotoran yak dalam siklus material ekosistem padang rumput.
4.3 Pola pelepasan N dan P pada serasah dengan perlakuan yang berbeda
Hasil kami menunjukkan bahwa imobilisasi N sementara terjadi selama dekomposisi untuk semua perlakuan. Namun, durasi imobilisasi N bervariasi tergantung pada jenis serasah (Gambar 4 ; Gambar S6 ). Untuk jenis serasah L dan L1, imobilisasi berlangsung dari 0 hingga 180 hari masa inkubasi, sedangkan untuk jenis serasah L2, berlanjut hingga 300 hari (Gambar S6 ). Di padang rumput rumput tinggi Kansas, akumulasi nitrogen diamati pada daun tanaman dan akar mati selama tahap awal dekomposisi (Seastedt, 1988 ). Selain itu, dekomposisi serasah di beberapa hutan beriklim sedang dan boreal melibatkan imobilisasi nitrogen awal, diikuti oleh pelepasan berikutnya (Vitousek & Sanford Jr, 1986 ). Pelepasan nitrogen bersih selama dekomposisi serasah terutama dikendalikan oleh rasio C: N, karena pengurai mikroba hanya akan melepaskan nitrogen setelah kebutuhan nitrogen mereka terpenuhi. Pada rasio C:N rendah (yaitu konsentrasi nitrogen tinggi), pengurai dapat memenuhi kebutuhan nitrogen mereka langsung dari serasah. Namun, pada rasio C:N awal yang lebih tinggi, imobilisasi bersih biasanya terjadi saat mikroba mengakses nitrogen dari luar serasah dan mengubahnya menjadi biomassa mikroba atau eksoenzim (Parton et al., 2007 ). Sebuah studi sebelumnya menunjukkan bahwa pelepasan bersih nitrogen serasah terjadi ketika rasio C:N serasah di bawah 5 hingga 15, sedangkan fiksasi nitrogen bersih terjadi pada rasio C:N awal yang lebih tinggi (Moore et al., 2011 ). Dalam studi ini, rasio C:N awal serasah L, L1 dan L2 masing-masing adalah 25,9, 27,6 dan 32,9 (Gambar 2 ). Ini menunjukkan bahwa defisiensi nitrogen merupakan faktor pembatas dalam dekomposisi serasah, dan periode imobilisasi nitrogen yang singkat diperlukan untuk memenuhi kebutuhan mikroorganisme tanah. Rasio C:P kritis yang menyebabkan mineralisasi fosfor dan sulfur bersih terjadi masing-masing sekitar 480 dan 300 (Blair & Crossley Jr, 1988 ; Gosz et al., 1973 ). Rasio C:P awal dari serasah L, L1 dan L2 semuanya lebih rendah dari rasio kritis (Gambar S2). ), yang menunjukkan bahwa ketersediaan fosfor dan sulfur melebihi kebutuhan mikroorganisme tanah.
Penambahan kotoran yak secara signifikan meningkatkan translokasi nutrisi, yang mengarah pada peningkatan pelepasan N dari jenis serasah berkualitas tinggi, seperti L1 (Gambar 3 ). Yang terpenting, durasi imobilisasi N dalam serasah berkualitas rendah (L2) dipersingkat karena penambahan kotoran, menghasilkan fiksasi N yang lebih sedikit dibandingkan dengan perlakuan tanpa penambahan kotoran (Gambar S6 ). Lebih jauh lagi, penambahan kotoran yak juga memfasilitasi translokasi P, terutama pada jenis serasah dengan konsentrasi P tinggi, yang merespons lebih signifikan terhadap efek promosi ini (Gambar S6 ). Selama periode dekomposisi 300 hari, keberadaan kotoran yak mempromosikan degradasi selulosa dan hemiselulosa dalam serasah, yang mengarah ke struktur serasah yang lebih longgar (Singh et al., 2016 ). Perubahan struktural ini memfasilitasi pemanfaatan fosfor intraseluler yang lebih besar oleh mikroba.
4.4 Keterbatasan
Beberapa keterbatasan masih ada di beberapa area studi ini. Pertama, jenis serasah kami terutama terdiri dari serasah rumput, daun semak dan batang semak; namun, indikator lain dari komposisi serasah, seperti gugus fungsi tanaman dalam serasah rumput, memerlukan penyelidikan lebih lanjut mengingat penelitian ekstensif tentang efek campurannya pada dekomposisi serasah (Liu et al., 2020 ). Kedua, ekskreta memperkenalkan makronutrien tambahan (misalnya P, K dan S) dan mikronutrien (misalnya Mg, Ca), yang memainkan peran penting dalam mengatur aktivitas mikroba dan siklus nutrisi. Misalnya, ketersediaan P memengaruhi sintesis adenosin trifosfat (ATP) dan degradasi lignin (Luo et al., 2022 ). Selain itu, studi ini hanya berfokus pada penambahan ekskreta dalam eksperimen inkubasi, tanpa mempertimbangkan aktivitas penggembalaan lainnya, seperti ternak menginjak-injak dan perilaku mencari makan. Oleh karena itu, upaya di masa depan harus menggabungkan eksperimen lapangan untuk mengklarifikasi efek dari faktor-faktor ini.
5 KESIMPULAN
Singkatnya, hasil kami menunjukkan bahwa jenis serasah tanaman dan kotoran yak memiliki efek individual dan interaktif yang signifikan pada dekomposisi serasah, terutama pada tahap awal. Jenis serasah terutama memengaruhi dekomposisi serasah dengan mengubah kualitas awal, sementara kotoran yak memiliki dampak positif pada dekomposisi serasah melalui peningkatan kadar nutrisi tanah dan aktivitas mikroba tanah. Yang penting, penelitian kami mengungkapkan bahwa kualitas serasah awal adalah penentu utama efek pencampuran serasah, dan penambahan kotoran yak hanya dapat memengaruhi besarnya efek ini. Khususnya, kami menyarankan bahwa ada imobilisasi N sementara selama tahap dekomposisi awal dan pola pelepasan P bersih dari waktu ke waktu untuk semua kombinasi serasah dan kotoran. Di masa mendatang, lebih banyak upaya harus menggabungkan eksperimen lapangan untuk mengklarifikasi efek kelompok fungsional serasah tanaman dan kemajuan penggembalaan lainnya pada dekomposisi serasah.


