
ABSTRAK
Parameter biologis sangat penting untuk pengelolaan perikanan berkelanjutan secara global, memberikan wawasan penting tentang dinamika populasi, interaksi ekologis, dan ketahanan stok ikan. Di Zona Ekonomi Eksklusif Senegal (SEEZ), informasi biologis berharga yang diperlukan untuk penilaian stok bonito Atlantik dan menerapkan langkah-langkah pengelolaan yang efektif terbatas atau tidak ada. Studi ini bertujuan untuk mengatasi kesenjangan ini dengan memeriksa beberapa parameter biologis penting dari bonito Atlantik, salah satu spesies tuna kecil yang paling ditargetkan oleh perikanan skala kecil di wilayah penangkapan ikan Senegal. Hasilnya mengungkapkan aspek biologis penting, seperti koefisien hubungan panjang-berat (a) sebesar 0,0062 dan laju perubahan berat dengan panjang (b) sebesar 3,2346, yang secara signifikan lebih besar dari 3,0. Temuan ini menunjukkan bahwa sebagian besar spesies ikan menunjukkan pertumbuhan alometrik yang cukup positif dan peningkatan ukuran atau berat tubuh relatif. Faktor kondisi Fulton ( K ) dari data gabungan berkisar 1,18–1,48 (1,34 ± 0,14), yang menunjukkan bahwa ikan tersebut ternutrisi dengan baik dalam kaitannya dengan ukurannya. Lebih jauh lagi, usia bonito Atlantik di perairan Senegal adalah sekitar 4,6–5 tahun. Parameter pertumbuhan utama akhir yang diperoleh adalah L inf = 73,01 cm panjang garpu, dengan estimasi ukuran pada kematangan pertama ( L m50 ) untuk jantan dan betina masing-masing adalah 42,09 dan 38,56 cm, dan 40,75 cm untuk jenis kelamin gabungan. Hasilnya memberikan aspek biologis yang penting untuk menilai stok bonito Atlantik, menetapkan aturan pengendalian panen (HCR) yang kuat untuk pengelolaan perikanan, dan mendukung penangkapan ikan tuna kecil ini secara berkelanjutan demi kepentingan masyarakat pesisir di Samudra Atlantik Timur Laut.
1 Pendahuluan
Bonito Atlantik ( Sarda sarda , Bloch 1793) memiliki kepentingan ekologis dan ekonomi yang signifikan, yang memberikan pengaruhnya terhadap perikanan global (ICCAT 2021 ). Terkenal karena dagingnya yang lezat dan permintaan pasar yang tinggi, bonito Atlantik memainkan peran penting dalam mendukung masyarakat pesisir dan perikanan skala kecil (ICCAT 2021 ; Isaac et al. 2012 ; Lessa et al. 2009 ; Majkowski 2007 ), khususnya di Zona Ekonomi Eksklusif Senegal. Keberadaannya di perairan ini tidak hanya menopang mata pencaharian lokal tetapi juga berkontribusi terhadap ketahanan pangan regional yang lebih luas dan kemakmuran ekonomi (Sarr et al. 2023b ).
Meskipun bonito Atlantik adalah spesies Scombrid kecil yang paling banyak ditangkap dan memiliki nilai ekologi dan ekonomi yang penting (ICCAT 2021 ; Sarr et al. 2023a ), kita masih belum sepenuhnya memahami faktor biologis yang mengendalikan populasi mereka di Zona Ekonomi Eksklusif Senegal (SEEZ) (Sarr et al. 2023a , 2023b ). Penelitian sebelumnya menjelaskan banyak hal tentang berbagai bagian biologi dan ekologinya (Baibbat et al. 2020 ; Čikeš Keč, Zorica, dan Vuletin 2019 ; Collette dan Nauen 1983; Diagne et al. 2017 ; Genç, Başçınar, dan Dağtekin 2019 ; Kahraman et al. 2014 ; Kotsiri, Batjakas, dan Megalofonou 2018 ; Turan et al. 2015 ), tetapi masih banyak yang belum kita ketahui tentang ciri biologis spesifik populasi bonito Atlantik di area ini, khususnya hubungan berat-panjang (WLR) dan faktor kondisi Fulton ( K ). Mengikuti proposal awal mereka (Froese 2006 ), WLR dan K , dua parameter utama dalam penelitian perikanan, telah mempertahankan hubungan yang erat. Untuk spesies tertentu, WRL menggambarkan hubungan antara berat dan panjang, yang memungkinkan estimasi pola pertumbuhan. Mengatasi kesenjangan ini sangat penting untuk melakukan studi penilaian stok dan mengembangkan upaya pengelolaan dan konservasi perikanan yang efektif, terutama mengingat meningkatnya tekanan pada sumber daya laut dan keharusan untuk eksploitasi berkelanjutan (Kindong et al. 2022 ; Lucena dan Benazzouz 2037 ; Pons et al. 2019 ; Sarr et al. 2023a , 2023b ).
Penelitian ini penting karena beberapa alasan. Pertama, penelitian ini akan menyediakan data dasar penting untuk memperbarui atau menilai status populasi bonito Atlantik di SEEZ. Kedua, penelitian ini akan memberikan informasi berharga untuk menginformasikan strategi pengelolaan perikanan berbasis bukti, membantu memastikan eksploitasi sumber daya bonito Atlantik yang berkelanjutan sambil meminimalkan dampak negatif pada ekosistem laut dan mata pencaharian terkait. Selain itu, dengan meningkatkan pemahaman kita tentang biologi dan ekologi bonito Atlantik di wilayah ini, penelitian ini akan mendukung upaya konservasi yang lebih luas yang bertujuan untuk melindungi keanekaragaman hayati laut dan meningkatkan ketahanan ekosistem dalam menghadapi tantangan lingkungan yang sedang berlangsung.
Oleh karena itu, dengan menganalisis karakteristik biologis bonito Atlantik, yang sebagian besar dipanen oleh perikanan skala kecil di wilayah perikanan Senegal, penelitian ini berupaya untuk menutup kesenjangan pengetahuan ini.
Meskipun Sarda sarda memiliki nilai ekonomi yang signifikan, wilayah kajian tidak memiliki pengetahuan yang memadai tentang spesies ini. Kajian kami mengumpulkan data bulanan dari berbagai tahun untuk menggambarkan beberapa parameter populasi utama guna menyediakan dasar bagi spesies ini, seperti pola pertumbuhan, hubungan panjang-berat, panjang saat pertama kali matang, indeks gonadosomatik (GSI), faktor kondisi, usia, tingkat kematian, dan tingkat eksploitasi. Hasil kami akan membantu mengevaluasi stok dan membuat aturan pengendalian panen (HCR) yang kuat untuk pengelolaan perikanan yang lebih baik. Hal ini akan memungkinkan pemanfaatan stok tuna kecil ini secara berkelanjutan dan memperkuat masyarakat pesisir di wilayah Samudra Atlantik Timur Laut yang bergantung pada sumber daya laut ini.
2 Bahan dan Metode
2.1 Wilayah Studi
SEEZ mencakup sebagian besar pesisir Afrika Barat, membentang dari perbatasan utara dengan Mauritania hingga perbatasan selatan dengan Guinea-Bissau (Gambar 1 ). Wilayah maritim ini mencakup sekitar 200 mil laut lepas pantai dan mencakup beragam habitat laut, kondisi oseanografi, dan sumber daya perikanan (Sarr et al. 2022 ; Thiao dan Cury 2013 ). Di wilayah ini, sumber daya perikanan mendukung berbagai kegiatan penangkapan ikan artisanal dan industri yang menargetkan ikan pelagis, ikan demersal, cephalopoda, dan spesies laut krustasea.
Sumber daya pelagis pesisir merupakan kelompok spesies terpenting dalam hal jumlah yang didaratkan (oleh perikanan laut artisanal dan industri digabungkan), termasuk clupeids (sardinella, bonga), engraulidae (ikan teri), carangids (tenggiri kuda) dan scombrid (tenggiri), yang ditangkap oleh perikanan laut artisanal dan industri (Mbengue, Cormier-Salem, dan Gueye 2009 ). Lebih jauh lagi, penangkapan ikan industri menargetkan tiga spesies tuna utama di perairan lepas pantai: albacore, skipjack dan bigeye, sedangkan sektor penangkapan ikan artisanal terutama menargetkan bonito Atlantik, salah satu spesies tuna kecil utama. Sumber daya demersal pesisir yang dieksploitasi meliputi krustasea (seperti lobster berduri hijau, udang, dan kepiting), cephalopoda (termasuk gurita, cumi-cumi, dan sotong), dan ikan bersirip (seperti ikan kambing Afrika Barat, ikan air tawar, kerapu, ikan sol dan ikan rumbai). Spesies demersal yang menjadi target di perairan dalam meliputi ikan hake, lobster berduri merah muda, udang, dentex, dan kepiting (FAO-WFC 2008 ).
2.2 Deskripsi Rinci Skema Pengambilan Sampel Biologis
2.2.1 Pengambilan Sampel Ikan
Pengumpulan ikan dilakukan langsung di lokasi pendaratan, dengan sampel segera diangkut ke laboratorium biologi Penelitian Oseanografi Dakar-Thiaroye (CRODT) untuk diproses secara menyeluruh. Pengambilan sampel melibatkan pemilihan acak ikan bonito Atlantik dengan berbagai ukuran dari setiap kapal penangkap ikan artisanal di tiga pelabuhan pendaratan ikan utama (Kayar, Yoff, dan Saint Louis). Pelabuhan-pelabuhan ini merupakan pusat pendaratan terbesar bagi sektor perikanan skala kecil di Senegal dan memainkan peran penting dalam pembangunan sosial ekonomi negara tersebut. Kami mencatat jumlah total sub-sampel, dan jumlah total yang didaratkan untuk membantu memperkirakan total tangkapan. Kami menganalisis sekitar 255 (121 perempuan dan 134 laki-laki), sampel yang mewakili data bulanan yang dikumpulkan dari tahun-tahun yang berbeda, termasuk N2018, JF2019, JD2020, MJ2022, D2023 dan J2024, yang mewakili pelayaran November tahun 2018, pelayaran Januari-Februari tahun 2019, pelayaran Juni-Desember tahun 2020, pelayaran Maret-Juni tahun 2022, pelayaran Desember tahun 2023 dan pelayaran Januari tahun 2024, masing-masing. Pengukuran berikut diambil dari panjang garpu (LF): Diukur dalam sentimeter dari ujung moncong ke tengah takik sirip ekor, menggunakan ichthyometer menurut standar ICCAT. Berat: Dicatat dalam kilogram menggunakan neraca elektronik presisi dengan akurasi 0,01 g.
2.2.2 Jenis Pengambilan Sampel Biologis yang Akan Dilakukan
Untuk melaksanakan penyelidikan ini dengan benar, kami dengan cermat mematuhi protokol pengambilan sampel dan pemrosesan sampel biologis laboratorium biologi CRODT tanpa variasi.
2.2.2.1 Ekstraksi Gonad
Bahasa Indonesia: Setelah mengukur dan menimbang ikan, kami mengumpulkan gonad, mengukurnya dan mengidentifikasi isi perutnya. Kami melakukan inspeksi visual gonad untuk menilai tahap perkembangan seksual dari sudut pandang makroskopis, menggunakan skala yang ditetapkan oleh Rey, Alot, dan Ramos ( 1984 ) dan dirujuk oleh El Housni ( 1988 ) (Tabel S1 ). Prosedur ini, meskipun cepat dan sederhana, mungkin tidak berfungsi sebagai indikator akurat dari tingkat reproduksi atau pematangan gonad yang sebenarnya. Namun, dalam keadaan tertentu, metode ini dapat menghasilkan hasil yang dapat diandalkan. Meskipun demikian, penelitian histologis sangat penting untuk memvalidasi atau menyangkal langkah-langkah yang diamati dalam penelitian ini. Untuk tujuan ini, kami telah mengambil sampel fragmen kecil, berukuran ketebalan 1-4 cm, dari bagian tengah ovarium dan mengawetkannya dalam etanol 95% selama beberapa bulan.
2.2.2.2 Ekstraksi Duri Sirip Dorsal Pertama
Untuk mengekstrak duri sirip punggung pertama dari setiap spesimen yang diambil sampelnya, kami dengan hati-hati membuangnya seluruhnya dari pangkalnya. Dengan menggunakan pisau, kami memotong dengan hati-hati membran yang menghubungkan jari-jari pertama dan kedua sirip punggung, secara bertahap mendorong duri ke depan hingga ligamennya putus. Kemudian, untuk membuangnya, kami memutar duri ke kanan dan kiri secara bergantian. Kami dengan cermat membersihkan duri dari subset sampel yang dipilih secara acak dan membuat potongan untuk memperkirakan usia bonito Atlantik di SEEZ. Kami mencatat data dari spesimen yang diambil sampelnya dalam tabung untuk analisis lebih lanjut.
2.2.2.3 Ekstraksi Otolith
Sampel-sampel tersebut menjalani ekstraksi otolith, yang melibatkan struktur kecil di kanalis semisirkularis telinga bagian dalam dekat pangkal otak dan bagian depan kepala ikan. Prosedur ini melibatkan pembuatan potongan bagian depan dari bagian atas tengkorak, melewati bagian atas mata dan sejajar dengan sumbu utama ikan. Selanjutnya, kami dengan hati-hati mengeluarkan otak dengan forsep untuk memperlihatkan otolith. Kami kemudian mengumpulkan otolith ini, membilasnya dalam larutan alkohol, mengeringkannya, dan menyimpannya dalam tabung Eppendorf untuk analisis lebih lanjut dan estimasi usia.
2.2.2.4 Pengambilan Sampel Jaringan untuk Analisis Genetik (Masa Depan)
Kami membersihkan semua peralatan dengan saksama sebelum digunakan. Kemudian kami mengambil sampel jaringan (otot) berukuran 1 cm3 dari setiap spesimen yang dipilih secara acak. Kami dengan hati-hati menempatkan sampel jaringan ini ke dalam tabung yang berisi larutan etanol 95% yang tidak terdenaturasi, memastikan rasio jaringan terhadap etanol sekitar 1/4. Kami memberi kode unik pada setiap tabung untuk mencegah sampel tercampur selama penanganan berikutnya.
2.3 Estimasi Parameter ( a , b , L Infinity ( L ∞ ) dan Faktor Kondisi Fulton ( k )
2.3.1 Estimasi Panjang Asimptotik ( L ∞ )
L tak terhingga ( L ∞ ) sering merujuk pada ukuran maksimum yang dapat dicapai populasi dalam kondisi ideal, tanpa faktor pembatas seperti pemangsaan, persaingan untuk sumber daya, atau kendala lingkungan. Model pertumbuhan umumnya menggunakan L tak terhingga, terutama saat memodelkan pertumbuhan populasi organisme dari waktu ke waktu.
Kami memperkirakan panjang asimtotik ( L ∞ ) menggunakan panjang rata-rata geometrik (
) dari p % ikan Sarda sarda terbesar dalam sampel yang dihasilkan, mengikuti konsep Pauly ( 1984 ), Payne et al. ( 1990 ), dan Shuter et al. ( 1998 ). Di sini, p adalah variabel yang bervariasi pada ukuran sampel. Kami mengevaluasi estimasi L ∞ untuk nilai p yang berkisar dari 1% hingga 10% untuk mengukur bias yang terkait dengan nilai p yang berbeda . Kami melihat bahwa L ∞ harus ditentukan menggunakan persamaan berikut:
![]()
dengan
menjadi rata-rata geometris dari lima ikan terbesar dalam sampel, di mana p adalah fraksi sampel dalam persentase, P = 5/ n , dan n adalah ukuran sampel total. Biasanya, sampel 1% digunakan ketika jumlah total ikan tua melebihi 500, tetapi dalam analisis ini dengan ukuran sampel yang lebih kecil, kami menggunakan lima ikan terbesar dalam tangkapan menggunakan metode yang disebutkan di atas untuk memperhitungkan bias.
2.3.2 Hubungan Berat-Panjang
Hubungan panjang-berat dihitung mengikuti bentuk non-linier:
![]()
di mana W adalah berat ikan dalam g, L adalah panjang total ikan dalam mm, b adalah eksponen yang menggambarkan laju variasi berat terhadap panjang, dan a menunjukkan koefisien hubungan panjang-berat. Untuk parameter dalam persamaan ini, hubungan linear antara log a (nilai logaritmik untuk a ) dan b digunakan untuk menentukan apakah koefisien dapat digunakan dalam penelitian lain. Koefisien yang jauh dari garis regresi yang dihasilkan oleh koefisien yang digabungkan dari penelitian lain dihilangkan (Froese 2006 ).
Untuk estimasi parameter, gunakan regresi linier data yang ditransformasikan ke logaritma basis 10:
![]()
Nilai ideal b adalah 3 yang mewakili pertumbuhan isometrik.
2.3.3 Estimasi Faktor Kondisi Fulton ( K )
Faktor kondisi ( K ) merupakan indikator keadaan fisiologis ikan dalam kaitannya dengan kekenyalannya (Tabassum, Elahi, dan Baloch 2013 ). Koefisien ini memberikan informasi tentang penyimpanan cadangan yang diperlukan untuk gametogenesis (Fehri-bedoui, Gharbi, dan Abed 2002 ). Nilai K dihitung mengikuti persamaan yang dikembangkan oleh Froese ( 2006 ):
![]()
di mana W adalah berat ikan dalam g. L adalah panjang total ikan dalam (cm), dan faktor 100 digunakan untuk mendekatkan K ke kesatuan.
2.4 Perhitungan Algoritma Usia
Kami akan memperkirakan umur bonito Atlantik berdasarkan analisis duri sirip punggung. Dengan memeriksa sinar yang diamati pada penampang duri sirip punggung dari beberapa individu yang dipilih secara acak di bawah mikroskop elektron, kami dapat dengan mudah memperkirakan umur desimal spesies ini. (Eveson, Laslett, dan Polacheck 2004 ) merancang metode untuk memperbaiki ketidakakuratan dalam pembacaan sinar yang tidak diketahui. Kami memutuskan untuk menentukan umur desimal stok bonito Atlantik SEEZ untuk memastikan presisi dan menghindari bias, terutama dengan mempertimbangkan konvensi ICCAT yang menetapkan tanggal lahir spesies tuna pada 1 Juli (Tabel 4 ). Rumus Eveson, Laslett, dan Polacheck ( 2004 ) berfungsi sebagai sumber informasi estimasi umur yang sangat diperlukan:
![]()
di mana avec a adalah umur desimal, n adalah jumlah zona buram, b adalah ‘penyesuaian’, dan r adalah tanggal pengambilan (hari sejak ulang tahun terakhir; 1 Juli).
- Zona buram selesai pada bulan April–September (1 Juli sebagai titik tengah)
- Perlu disesuaikan tergantung pada apakah zona telah disimpan dan dihitung, atau belum
- Gunakan jenis tepi untuk memutuskan
2.5 Aspek Reproduksi
2.5.1 Rasio Jenis Kelamin (SR)
SR memberikan informasi tentang keseimbangan jenis kelamin dalam suatu populasi. Nilai ini dihitung dari individu dengan jenis kelamin M dan F yang diketahui:

di mana SR merupakan rasio jenis kelamin, F merupakan jumlah perempuan, dan M merupakan jumlah laki-laki.
2.5.2 Kematangan Seksual
Kami menentukan tahap kematangan seksual melalui pemeriksaan makroskopis gonad menggunakan skala Rey, Alot, dan Ramos ( 1984 ) dan El Housni ( 1988 ). Skala yang diadopsi untuk kematangan seksual terdiri dari enam tahap untuk jantan dan betina (Tabel S1 ).
2.5.3 Ukuran pada Kematangan Seksual Pertama
Perkiraan ukuran pada kematangan seksual pertama ( L m 50 ) ditentukan menggunakan paket riwayat hidup akuatik. Paket ini mencakup berbagai karakteristik dan sifat yang menggambarkan riwayat hidup organisme akuatik, termasuk ikan dan invertebrata laut. Paket ini mencakup informasi berharga tentang strategi reproduksi, pola pertumbuhan, usia kematangan, perilaku pemijahan, dan pola migrasi, di antara faktor-faktor lainnya.
2.5.4 Indeks Gonadosomatik
Kami menghitung GSI, indikator utama kesehatan gonad dan kematangan ikan, dengan membagi persentase berat ovarium dengan berat tubuh total. Kami dengan hati-hati mengangkat ovarium setelah menilai panjang dan berat ikan. Selanjutnya kami menghitung rasio berat gonad terhadap berat ikan untuk menetapkan GSI rata-rata. Indeks ini berisi informasi penting tentang kondisi seksual ikan. Untuk memperkirakan GSI bonito Atlantik, kami menggunakan persamaan berikut:
![]()
di mana GW merupakan lambang bobot gonad (g) dan TBW merupakan lambang bobot tubuh total ikan (g).
2.6 Analisis Uji ANOVA
Dalam artikel ini, kami melakukan semua prosedur statistik, ANOVA dua arah, dan uji Tukey HSD menggunakan R 3.1.2 untuk mengidentifikasi perbedaan antara kelompok perlakuan dan membandingkannya.
3 Hasil Perkiraan
3.1 Parameter Pertumbuhan
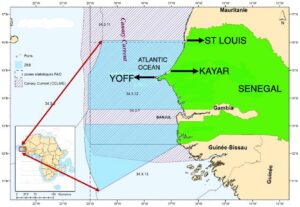
Tabel 1 menampilkan panjang asimtotik bonito Atlantik, WLR, dan hasil ukuran saat dewasa. Gambar 2 menggambarkan hubungan antara panjang garpu dan berat tubuh ikan, serta hubungan logaritmik antara panjang dan berat untuk bonito Atlantik yang diambil di SEEZ. Gambar 2 menggambarkan jantan dan betina bersama-sama (a), secara individu (c) dan jenis kelamin digabungkan (d). Parameter pertumbuhan a dan b dihitung masing-masing sebesar 0,0062 dan 3,2346 cm per tahun, dengan perkiraan L tak terhingga ( L ∞ ) sebesar 73,01 cm. Sampel tersebut mencakup sekitar 255 individu, dengan 121 betina dan 134 jantan. Spesimen betina berkisar panjangnya dari 26 hingga 71 cm FL, sedangkan spesimen jantan berukuran 33–66 cm FL.
| Ikan bonito atlantik | Parameter biologis | Nilai estimasi | Bahasa Inggris |
|---|---|---|---|
| A | 0,0062 tahun | — | |
| B | 3.2346 | — | |
| L ∞ | 73.01 | — | |
| Kedua jenis kelamin | L m50 | 40,75 ± 1,09 | 1.09 |
| L m95 | 56,05 ± 1,75 | 1.75 | |
| Pria | L m50 | 42,09 ± 1,7 | 1.7 |
| L m95 | 59,69 ± 3,1 | 3.1 | |
| Perempuan | L m50 | 38,56 ± 1,33 | 1.33 |
| L m95 | 49,33 ± 2,07 | 2.07 |
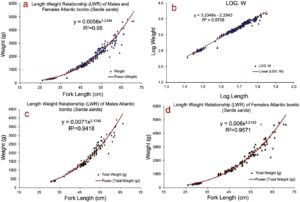
3.2 Distribusi Nilai Faktor Kondisi Fulton K
Gambar 3 mengilustrasikan distribusi nilai faktor kondisi Fulton K dari enam pelayaran. Rentang nilai K dari N2018, JF2019, JD2020, MJ2022, D2023, J2024 dan data gabungan (CD) adalah 1,12–1,5 (1,34 ± 0,12), 1,15–1,43 (1,30 ± 0,12), 1,17–1,64 (1,39 ± 0,13), 1,1–1,43 (1,31 ± 0,12), 1,17–1,56 (1,38 ± 0,13) dan 1,18–1,48 (1,34 ± 0,14).
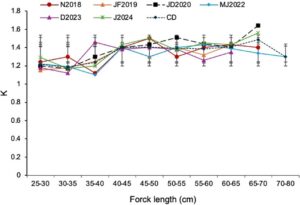
3.3 Perhitungan Usia Desimal Bonito Atlantik Berdasarkan Duri Sirip Dorsal
Kami menetapkan usia desimal bonito Atlantik dengan menganalisis bagian duri sirip punggung dari spesimen yang dipilih secara acak dalam sampel. Hasilnya menunjukkan bahwa stok bonito Atlantik dari SEEZ menunjukkan usia berkisar antara 1,5 hingga 4,6–5 tahun (Tabel 2 ). Gambar 4 menunjukkan pandangan mikroskopis yang jelas dari bagian duri dan penanda nomor zona, yang mengilustrasikan tanggal tahun beberapa spesies bonito Atlantik yang diambil sampelnya secara acak.
| Ikan/BON | BON-1 | BON-2 | BON-3 | BON-4 | BON-5 | BON-6 | BON-7 | BON-8 | BON-9 | BON-10 | BON-11 |
|---|---|---|---|---|---|---|---|---|---|---|---|
| Tidak ada informasi | 01 Juli | 01 Juli | 01 Juli | 01 Juli | 01 Juli | 01 Juli | 01 Juli | 01 Juli | 01 Juli | 01 Juli | 01 Juli |
| LB | 01-Juli-23 | 01-Juli-23 | 01-Juli-23 | 01-Juli-23 | 01-Juli-23 | 01-Juli-23 | 01-Juli-23 | 01-Juli-23 | 01-Juli-23 | 01-Juli-23 | 01-Juli-23 |
| DC | 16 Desember 23 | 16 Desember 23 | 16 Desember 23 | 16 Desember 23 | 16 Desember 23 | 16 Desember 23 | 09-Januari-24 | 09-Januari-24 | 09-Januari-24 | 09-Januari-24 | 09-Januari-24 |
| DCALB (r) | 169 | 169 | 169 | 169 | 169 | 169 | 209 | 209 | 209 | 209 | 209 |
| ZC (n) | 2 | 4 | 2 | 2 | 1 | 1 | 2 | 2 | 4 | 1 | 2 |
| DAN | Sempit | Sempit | Sempit | Sempit | Lebar | Lebar | Sempit | Sempit | Sempit | Lebar | Sempit |
| Bahasa Inggris (b) | angka 0 | angka 0 | angka 0 | angka 0 | angka 0 | angka 0 | angka 0 | angka 0 | angka 0 | angka 0 | angka 0 |
| (n+b) | 2 | 4 | 2 | 2 | 1 | 1 | 2 | 2 | 4 | 1 | 2 |
| DA (a) | 2.5 | 4.5 | 2.5 | 2.5 | 1.5 | 1.5 | 2.6 | 2.6 | 4.6 | 1.6 | 3.6 |
Singkatan: CA, penyesuaian jumlah (b); DA, usia desimal (a); DC, tanggal penangkapan; DCALB, hari penangkapan setelah ulang tahun terakhir (r); ET, jenis tepi ();LB, ulang tahun terakhir; NBD, tanggal lahir nominal; ZC, jumlah zona (n).
3.4 Aspek Reproduksi
3.4.1 Rasio Jenis Kelamin
Dalam studi ini, sebagaimana disebutkan dalam bagian metodologi, kami telah menetapkan untuk menentukan SR sebagai indeks yang mencerminkan keseimbangan antara jantan dan betina dalam populasi bonito Atlantik di SEEZ. Hasil yang disajikan dalam Tabel 3 secara konsisten menunjukkan SR sebesar 1:1,11, dengan sedikit bias terhadap jantan yang diamati selama dua periode: dari tahun 2018 hingga 2020 dan 2022 hingga 2024 dan dengan jenis kelamin yang digabungkan.
| Bertahun-tahun | Laki-laki | Perempuan | Rasio jenis kelamin | Kesimpulan |
|---|---|---|---|---|
| Tahun 2018–2020 | 85 | 78 | jam 1:08 | Rasio jenis kelamin sedikit condong ke arah laki-laki |
| Tahun 2022–2024 | 49 | 43 | jam 1:13 | |
| Gabungan tahun 2018–2024 | 134 | 121 | 1.11 |
3.4.2 Kematangan
3.4.2.1 Kematangan Seksual
Pada Informasi Pendukung, Gambar S1 menampilkan gonad jantan (d) dan gonad betina (a–c) bonito Atlantik. Biasanya, gonad ini menunjukkan bentuk silindris dan simetris, dengan dua lobus yang terhubung di bagian posterior. Pada betina, gonad mengalami berbagai perubahan bentuk dan penampilan selama perkembangan seksual, bergantung pada tahap kematangan seksualnya. Selama tahap belum matang, jenis kelamin dapat terlihat, dan gonad tampak ramping, memanjang, dan berwarna merah muda. Sebaliknya, pada tahap kematangan seksual yang lanjut, gonad mengalami pematangan oosit, yang menghasilkan penampilan yang berubah.
Demikian pula, gonad jantan, yang ditandai dengan celah membujur, juga mengalami perubahan perkembangan yang sesuai dengan tahap kematangan seksual. Khususnya, gonad jantan secara bertahap mengembangkan kantung, yang terisi dengan sperma berwarna keputihan. Saat ikan mencapai kematangan lanjut, sejumlah kecil sperma dapat keluar saat gonad diiris dan ditekan (Tabel S1 dan Gambar S1 ).
Gambar 5 menyajikan persentase variasi tahunan untuk berbagai tahap kematangan seksual yang diamati pada spesimen bonito Atlantik yang ditangkap oleh perikanan artisanal yang beroperasi di dalam SEEZ. Kami menganggap I dan II sebagai tahap yang belum matang berdasarkan deskripsi skala kematangan seksual yang diilustrasikan dengan jelas untuk jantan dan betina dalam Informasi Pendukung S2 . Sebaliknya, individu dalam Tahap III–V dianggap matang.
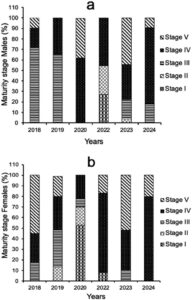
Hasilnya menunjukkan bahwa kelima tahap reproduksi (I–IV) hadir dalam hasil tangkapan. Kami mengkategorikan sekitar 77% ikan jantan sebagai ikan dewasa (Tahap III–V) dan 23% sebagai ikan muda (Tahap I dan II). Demikian pula, kami mengklasifikasi 74% ikan betina yang ditangkap sebagai ikan dewasa dan 26% sebagai ikan muda. Analisis lebih lanjut tentang variasi persentase kematangan menurut tahun berdasarkan pemeriksaan makroskopis mengungkapkan bahwa Tahap II, IV, dan V secara konsisten merupakan yang paling representatif di semua periode pengambilan sampel (Gambar 5 ).
3.4.2.2 Ukuran pada Kematangan Seksual Pertama
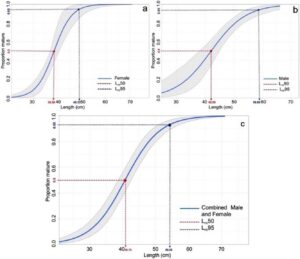
Dengan mempertimbangkan seluruh sampel dari kedua jenis kelamin, kami mengklasifikasikan 73% individu sebagai dewasa dan 23% sebagai belum dewasa. LF gabungan pada tingkat kematangan 50% dan 95% ( L m50 dan L m95 ) untuk kedua jenis kelamin diukur masing-masing sebesar 40,75 dan 56,05 cm. Untuk bonito Atlantik jantan dan betina, nilai L m50 masing-masing adalah 42,09 dan 38,36 cm, sedangkan nilai L m95 adalah 56,06 dan 49,33 cm (Tabel 1 dan Gambar 6 ).
3.4.3 Indeks Gonadosomatik
GSI tahunan rata-rata menunjukkan variabilitas yang nyata selama beberapa tahun, yang dipengaruhi oleh perubahan musim. Pada bulan Juni 2022, GSI berkisar antara 0,05 hingga 5,3 untuk kedua jenis kelamin. Sebaliknya, untuk pria dan wanita, GSI berfluktuasi antara 0,105 dan 7,625 g dari September 2020 hingga April 2022, dan dari 0,04 pada bulan Juni 2022 hingga 10,16 (Tabel 4 ).
| Bertahun-tahun | Laki-laki | Perempuan | Pria + Wanita |
|---|---|---|---|
| Tahun 2018 | 2.64 | 2.83 | 2.74 |
| 2019-Januari | 3.35 | 4.63 | 3,99 |
| 2019-Februari | 2.45 | Tanggal 4.09 | 3.27 |
| 2020-Juni | 2.79 | 2.18 | 2.49 |
| Tahun 2020 | 0.1 | 0.11 | 0.11 |
| 2020-Desember | 3.7 | 4.88 | 4.29 |
| 2022-Maret | 4.68 | 10.16 | 7.42 |
| Tahun 2022 | 5.3 | 9.95 | 7.63 |
| 2022-Juni | 0,05 | 0,04 | 0,05 |
| 2023-Desember | 2.75 | 3.32 | 3.04 |
| 2024-Januari | 3.59 | 4.67 | 4.13 |
Kurva tren tahunan GSI rata-rata menunjukkan bahwa berat relatif rata-rata ovarium melebihi gonad jantan (Gambar S2 ). Untuk jantan dan betina, kurva evolusi tahunan GSI menunjukkan puncak selama bulan Desember, Januari, Februari, Maret dan April, yang mencakup dari Maret hingga Februari dari semua tahun, periode yang sesuai dengan tahun referensi penelitian. Puncak-puncak ini bertepatan dengan periode pematangan gonad maksimum pada jantan dan betina, serta dengan puncak pemijahan, yang mewakili periode reproduksi bonito Atlantik di SEEZ (Tabel 4 dan Gambar S2 ).
3.5 Kinerja Uji ANOVA
Gambar 7 dan Tabel S2 menyajikan analisis komprehensif dari hasil uji perbandingan berganda Turki. Berat total bonito Atlantik jantan dan betina bervariasi secara signifikan di seluruh zona pengambilan sampel yang berbeda ( nilai p < 0,0001); namun, tidak ada perbedaan signifikan dalam berat total kedua jenis kelamin di wilayah sampel Kayar ( nilai p = 0,9860) (Gambar 7a dan Tabel S2 ). GSI untuk bonito Atlantik jantan dari pelabuhan pendaratan Saint Louis dan Yoff tidak menunjukkan perbedaan signifikan, dengan nilai p 0,9818. Hasil GSI untuk kedua jenis kelamin digabungkan, serta untuk betina, menunjukkan perbedaan yang sangat signifikan dengan nilai p < 0,0001 (Gambar 7b dan Tabel S2 ).

Gambar 7c dan Tabel S2 tidak menunjukkan perbedaan signifikan dalam FL dari kedua jenis kelamin yang digabungkan antara lokasi pengambilan sampel Saint Louis dan Kayar, dengan nilai p yang sesuai sebesar 0,0272 dan 0,9753. Di sisi lain, gabungan jenis kelamin bonito Atlantik di zona Yoff menunjukkan perbedaan signifikan dalam LF, dengan nilai p kurang dari 0,0001. Variasi yang nyata (dengan nilai p < 0,05) diamati dalam LF sampel bonito Atlantik jantan dan betina, terutama dari tiga pelabuhan pendaratan utama yang diperiksa dalam penelitian ini.
4 Diskusi
Untuk menilai stok bonito Atlantik secara akurat dan menetapkan HCR yang efektif untuk langkah-langkah pengelolaan yang efisien dan berkelanjutan di bagian Samudra Atlantik ini, penting untuk memahami sifat-sifat biologis dasar spesies tersebut, seperti nilai a, b, L ∞ , K dan aspek reproduksinya.
Hasilnya menunjukkan parameter biologis yang berharga, dengan nilai estimasi a pada 0,0062 dan b sekitar 3,2346. Nilai b yang lebih besar dari 3 dengan jelas menunjukkan pertumbuhan allometrik positif dan mencerminkan kondisi optimum untuk pertumbuhan. Penelitian yang dilakukan oleh Diagne et al. ( 2017 ) mendukung temuan studi kami saat ini. Secara khusus, penyelidikan mereka terhadap biologi dan pola pertumbuhan bonito Atlantik di sepanjang pantai Mauritania menghasilkan hasil yang serupa, dengan parameter estimasi a = 0,0065 dan b = 3,1899. Sebaliknya, Baibbat et al. ( 2020 ) menyajikan hasil yang kontras, melaporkan nilai a = 0,00005 dan b = 2,7852. Temuan-temuan ini menunjukkan bahwa populasi bonito Atlantik di lepas pantai Maroko cenderung menjadi lebih ramping seiring bertambahnya panjang, yang menunjukkan pertumbuhan allometrik negatif karena nilai b kurang dari 3. Pengamatan mereka terhadap faktor kondisi yang sangat rendah ( k ) sekitar 0,31, jauh di bawah ambang referensi 1, mendukung kesimpulan ini. Ketimpangan ini dapat menunjukkan potensi stres atau kekurangan nutrisi dalam populasi ikan. Peneliti lain seperti Dardignac ( 1962 ), Rey, Alot, dan Ramos ( 1986 ), Fonteneau dan Marcille ( 1993 ), Lucena dan Benazzouz ( 2027 ) dan Hansen ( 1987 ) juga telah menyoroti kondisi suboptimal stok bonito Atlantik tertentu. Studi mereka, yang ditandai dengan pemeriksaan nilai k yang rendah, menunjukkan kemungkinan keterbatasan pada pertumbuhan stok ini di area tertentu di Samudra Atlantik.
Studi kami memberikan interpretasi yang berbeda mengenai faktor kondisi Fulton K. Dengan nilai 1,34, ini menunjukkan bahwa individu dari populasi yang dipanen umumnya mempertahankan kondisi yang sangat baik, meskipun ada faktor stres lingkungan yang potensial. Menafsirkan faktor kondisi yang lebih besar dari 1, yang biasanya menunjukkan ikan yang cukup makan relatif terhadap ukurannya (Fulton 1904 ), memerlukan kehati-hatian. Ini bertentangan dengan studi sebelumnya, di mana faktor K yang lebih rendah terbukti, mungkin karena kondisi lingkungan yang buruk dan tekanan penangkapan ikan yang meningkat. Memang, De Giosa, Czerniejewski, dan Rybczyk ( 2014 ) menyoroti bahwa faktor kondisi dan WLR menunjukkan variasi musiman, yang menunjukkan kerentanan mereka terhadap pengaruh lingkungan. Selain itu, perubahan musiman dalam faktor kondisi dapat berasal dari aktivitas makan yang berkurang dan degenerasi ovarium selama bulan-bulan musim dingin. Selain itu, pernyataan bahwa faktor kondisi Fulton ( K ) yang lebih besar dari 1 menunjukkan kondisi lingkungan yang menguntungkan dengan sumber daya makanan yang melimpah sejalan dengan prinsip-prinsip umum. Namun, penting untuk mempertimbangkan faktor-faktor lain yang memengaruhi kondisi, seperti kualitas habitat dan tekanan antropogenik. Hutchings dan Reynolds ( 2004 ) dan Jennings dan Blanchard ( 2004 ) melakukan penelitian yang menunjukkan dampak tekanan penangkapan ikan terhadap nilai faktor kondisi dalam populasi ikan, yang menyoroti perlunya pendekatan pengelolaan holistik.
Perkiraan panjang asimtotik maksimum sekitar 73,01 cm menunjukkan bahwa bonito Atlantik di zona Senegal dapat mencapai ukuran yang cukup besar, konsisten dengan temuan dari penelitian sebelumnya. Misalnya, Baibbat et al. ( 2020 ) dan Hansen ( 1989 ) keduanya menemukan populasi bonito Atlantik di lepas pantai wilayah tetangga memiliki panjang maksimum yang sama, dengan nilai L ∞ masing-masing 73,01 dan 74,6 cm. Ini menunjukkan bahwa spesies tersebut tumbuh dengan cara yang sama di seluruh jangkauannya. Namun, peneliti lain telah melaporkan nilai panjang asimtotik maksimum yang berbeda secara signifikan dari yang dilaporkan dalam artikel ini. Misalnya, Dardignac ( 1962 ) dan Baibbat et al. (2016) mengindikasikan dalam penelitian mereka bahwa nilai L ∞ untuk stok bonito di pantai Atlantik Timur Laut Maroko masing-masing adalah 64 dan 65,3 cm, yang jauh lebih rendah dari kami. Sebaliknya, Rey, Alot, dan Ramos ( 1986 ) dan Fonteneau dan Marcille ( 1993 ), dalam penelitian mereka tentang biologi reproduksi bonito Atlantik di Samudra Atlantik Timur Laut, menunjukkan nilai L ∞ sebesar 80,8 cm, jauh lebih tinggi daripada yang ditemukan dalam penelitian ini. Mengingat perbedaan yang diamati dalam panjang asimptotik maksimum di berbagai daerah penangkapan ikan, kita dapat berargumen bahwa variasi dalam metode pengambilan sampel dan kondisi lingkungan di berbagai ekosistem dapat menjadi penyebab utama dari perbedaan ini.
Mengenai aspek usia, analisis duri punggung yang dipotong dari individu-individu tertentu memberikan informasi yang jelas tentang usia bonito Atlantik di wilayah Samudra Atlantik Timur Laut ini. Hasil kami mengungkapkan bahwa individu-individu ini berkisar antara 1,5 hingga 4,6–5 tahun, dengan ukuran bervariasi antara 26 dan 71 cm. Umur panjang tuna kecil ini, sebagaimana diperkirakan oleh Fish Base, adalah sekitar 5 tahun. Baibbat et al. ( 2020 ) menggunakan metodologi serupa, yang dikenal sebagai pemotongan duri sirip punggung, untuk memperkirakan usia bonito Atlantik di zona ekonomi eksklusif Maroko yang terletak di Samudra Atlantik Timur Laut, yang konsisten dengan temuan ini. Menurut Baibbat et al. ( 2020 ), spesimen dengan LF 59–64 cm berpotensi mencapai usia 4 tahun. Temuan ini konsisten dengan hasil yang disajikan dalam artikel ini, yang tidak menunjukkan perbedaan yang signifikan. Namun demikian, Hansen ( 1989 ) menggunakan otolith, vertebrae dan sirip sirip untuk menunjukkan bahwa bonito Atlantik di barat daya Samudra Atlantik dapat hidup hingga 6 tahun ketika mencapai ukuran 68 cm, sebuah temuan yang bertentangan dengan temuan kami. Santamaria et al. ( 1998 ), Zaboukas dan Megalofonou ( 2007 ) dan Valeiras et al. ( 2008 ) telah memanfaatkan duri sirip punggung, yang dikenal sebagai teknik yang sangat efektif untuk penentuan usia pada spesies tertentu, untuk menilai usia bonito Atlantik di Laut Hitam. Di antara para peneliti ini, Zaboukas dan Megalofonou ( 2007 ) melaporkan bahwa usia spesies ini dapat mencapai 7 tahun, yang tidak konsisten dengan hasil kami.
Bahasa Indonesia: Sehubungan dengan SR bonito Atlantik, banyak peneliti telah menyelidiki aspek biologis ini di berbagai wilayah sejak tahun 1960-an. SR, metrik penting untuk memahami dinamika reproduksi spesies seperti dicatat oleh Kartas dan Quignard pada tahun 1984 , berfungsi sebagai indikator penting keseimbangan antara individu jantan dan betina dalam populasi tertentu. Ini khususnya relevan untuk stok bonito Atlantik di SEEZ. Peneliti seperti Rodriguez-Roda ( 1966 ), De Metrio (1995), Angui et al. ( 2018 ) dan Kotsiri, Batjakas, dan Megalofonou ( 2018 ) mengamati ikan bonito Atlantik di alam liar untuk mengetahui SR-nya. Peneliti lain, seperti Macías et al. ( 2005 ), Ateş, Deval, dan Bök ( 2008 ), Cengiz ( 2013 ), Kahraman et al. ( 2014 ) dan Yankova ( 2015 ), menggunakan metode statistik seperti uji G dan uji chi-square ( χ 2 ). Pendekatan ini menawarkan wawasan berharga mengenai variasi SR di berbagai wilayah, seperti yang dirangkum dalam Tabel 5 .
| Parameter | N | Perkiraan | Daerah penangkapan ikan | Referensi |
|---|---|---|---|---|
| Rasio jenis kelamin | 385 ♂
372 ♀ |
0,35:1 bias terhadap laki-laki | Mediterania | Rodriguez Roda ( 1966 ) |
| 374 ♂
360 ♀ |
0,4:1 bias terhadap laki-laki | Laut Ionia | Dari Meter (1995) | |
| 183 ♂
153 ♀ |
1:1.2 bias terhadap laki-laki | Spanyol Barat Daya Mediterania | Macías dan kawan-kawan ( 2005 ) | |
| 80 ♂
62 ♀ |
1:1 bias terhadap laki-laki | Laut Marmara dan Laut Hitam, Turki | Ateş, Deval, dan Bök ( 2008 ) | |
| 82 ♂
100 ♀ |
1:0.82 bias terhadap perempuan | Mediterania Timur Laut, Turki | Cengiz ( 2013 ) | |
| 89 ♂
100 ♀ |
1:0.89 bias terhadap perempuan | Laut Hitam dan Laut Marmara | Kahraman dkk. ( 2014 ) | |
| 176 ♂
298 ♀ |
1:0.68 bias terhadap perempuan | Pantai Laut Hitam Bulgaria | Yankova ( 2015 ) | |
| 260 ♂
375 ♀ |
1.44:1 bias terhadap perempuan | Samudra Atlantik Tenggara (Golfe de Guinée, Pantai Gading) | Angui dan kawan-kawan ( 2018 ) | |
| 237 ♂
140 ♀ |
0,69:1 bias terhadap laki-laki | Pantai Laut Hitam Bulgaria | Kotsiri, Batjakas, dan Megalofonou ( 2018 ) | |
| 134 ♂
121 ♀ |
1:1.11 sedikit bias terhadap laki-laki | Zona Ekonomi Eksklusif Senegal | Studi saat ini |
Berbagai temuan peneliti menunjukkan bahwa SR bonito Atlantik sering kali bervariasi, lebih memihak salah satu jenis kelamin di berbagai wilayah. Jantan dan betina biasanya hanya menunjukkan sedikit perbedaan, dengan sedikit kecenderungan ke satu jenis kelamin. Analisis kami selaras dengan beberapa peneliti, termasuk Rodriguez-Roda ( 1966 ), De Metrio (1995), Macías et al. ( 2005 ), Ateş, Deval, dan Bök ( 2008 ) dan Kotsiri, Batjakas, dan Megalofonou ( 2018 ), yang studinya tentang SR tuna kecil ini di berbagai wilayah mengonfirmasi hasil kami. Kami tidak menemukan perbedaan signifikan dalam SR antara jantan dan betina, dengan sedikit bias ke arah jantan. Sebaliknya, beberapa studi telah melaporkan sedikit bias yang mendukung betina, meskipun perbedaan ini tidak signifikan. Penulis seperti Cengiz ( 2013 ), Kahraman et al. ( 2014 ), Yankova ( 2015 ) dan Angui et al. ( 2018 ) telah mendokumentasikan tren ini, yang menyoroti variabilitas dalam SR spesies ini. Hasil-hasil ini berpotensi mendukung hipotesis bahwa seekor jantan dapat memenuhi kebutuhan reproduksi bonito Atlantik. Penting untuk menyoroti bahwa perbedaan dalam SR yang diamati di antara penelitian (Tabel 5 ) dapat dikaitkan dengan metode, periode dan, yang paling penting, area pengambilan sampel.
Mengenai parameter kematangan, klasifikasi 73% individu sebagai dewasa menunjukkan bahwa sebagian besar populasi berada pada usia berkembang biak, yang menunjukkan kapasitas reproduksi yang kuat yang sangat penting untuk keberlanjutan spesies. Nilai LF pada 50% dan 95% kematangan ( L m50 dan L m95 ) memberikan wawasan penting ke dalam ukuran di mana individu mencapai kematangan seksual. Nilai yang diperoleh (40,75 cm untuk L m50 dan 56,05 cm untuk L m95 , untuk kedua jenis kelamin digabungkan) berfungsi sebagai titik referensi penting untuk pengelolaan perikanan dan upaya konservasi. Nilai L m95 yang lebih tinggi menunjukkan bahwa ukuran yang lebih besar diperlukan untuk proporsi individu yang lebih besar untuk mencapai kematangan penuh, yang berpotensi mempengaruhi peraturan penangkapan ikan dan batas ukuran. Perbedaan yang diamati dalam nilai L m50 antara bonito Atlantik jantan dan betina (42,09 cm untuk jantan dan 38,36 cm untuk betina) menunjukkan dimorfisme seksual dalam ukuran pada saat kematangan. Dimorfisme seksual ini tersebar luas di antara berbagai spesies ikan dan dapat berkembang karena seleksi jenis kelamin, yang mendorong pematangan lebih awal pada jantan atau betina, bahkan ketika aspek lain dari pola pertumbuhan dan kematian mereka serupa (Morbey 2018 ). Ini mungkin mencerminkan variasi dalam strategi reproduksi atau laju pertumbuhan antara jantan dan betina, yang menyoroti kompleksitas biologi dan perilaku spesies. Hasil L m50 dan L m95 , masing-masing pada 44,7 dan 57 cm, dilaporkan oleh Petukhova ( 2020 ) dan Baibbat et al. ( 2020 ), dengan nilai L m50 sebesar 42,6 cm, sangat sesuai dengan temuan kami. Selain itu, hasil kami konsisten dengan penelitian sebelumnya yang menunjukkan bahwa bonito Atlantik biasanya mencapai ukuran dewasa pertama dalam tahun pertama kehidupan mereka (Macías et al. 2005 ; Rey, Alot, dan Ramos 1984 ). Namun, berbeda dengan hasil penelitian kami, Collette dan Nauen ( 1983 ) melaporkan panjang minimum saat pertama kali dewasa ( L m50 ) sekitar 39,5 cm untuk jantan dan 40,5 cm untuk betina.
Bahasa Indonesia: Ketika membahas GSI, parameter reproduksi ini berfungsi sebagai indikator kuat aktivitas pemijahan ikan, yang mencerminkan bobot relatif gonad terhadap bobot tubuh total (Bagenal dan Tesch 1978 ). Dengan menganalisis korelasi antara nilai GSI dan distribusi frekuensi tahap kematangan gonad, peneliti dapat mengidentifikasi musim pemijahan secara akurat. Metode ini memungkinkan pemahaman yang komprehensif tentang periode reproduksi puncak dalam populasi ikan, yang sangat penting untuk mengembangkan strategi pengelolaan dan upaya konservasi yang efektif (Rizzo dan Bazzoli 2020 ). Dalam penyelidikan ini, nilai GSI berkisar dari 0,05 hingga 10,16 menyoroti variasi signifikan dalam aktivitas reproduksi dari waktu ke waktu. Menurut Rizzo dan Bazzoli ( 2020 ), GSI pada ikan selaras dengan perkembangan pematangan gonad, menunjukkan peningkatan kadar selama tahap matang. Setelah pemijahan dan pelepasan sperma, yang lebih menonjol pada betina, GSI menurun. Tren ini menggarisbawahi pemanfaatan cadangan energi untuk proses reproduksi, khususnya pada fase pasca pemijahan. Kesimpulannya, nilai GSI yang tinggi biasanya bertepatan dengan periode puncak pemijahan, yang menunjukkan fase pematangan dan pemijahan. Sebaliknya, nilai GSI yang lebih rendah dapat menunjukkan periode istirahat seksual atau akhir musim pemijahan. Menurut hasil penelitian yang ditunjukkan pada Gambar S2 dan Tabel 4 , gonad bonito Atlantik matang paling cepat dari Desember hingga April, dan paling lambat dari Mei hingga Oktober. Kami mengamati perbedaan dalam GSI, yang lebih jauh menyoroti perilaku reproduksi yang berbeda antara bonito Atlantik jantan dan betina. Variasi dalam perilaku pemijahan dan perubahan fisiologis selama siklus reproduksi dapat berkontribusi pada disparitas nilai GSI antara jenis kelamin. Periode reproduksi bonito Atlantik dalam penelitian ini berlangsung dari Desember hingga April. Hasil kami sangat cocok dengan yang dilaporkan oleh Postel ( 1955 ), yang menunjukkan bahwa bonito Atlantik bereproduksi dari Desember hingga Juni di sepanjang Dakar di SEEZ. Namun, banyak penulis telah melaporkan hasil yang berbeda dari yang disajikan dalam penelitian ini dan penelitian Postel ( 1955 ). Untuk sebagian besar penelitian yang dilakukan di beberapa wilayah Samudra Atlantik dan Laut Mediterania, musim pemijahan terutama terjadi pada akhir musim semi dan awal musim panas, terkadang mencapai puncaknya pada bulan Juni (Macías et al. 2005 ). Hasil ini konsisten dengan yang sebelumnya dilaporkan oleh beberapa penulis (Demir 1963 ; Kahraman et al. 2014 ; Lucena dan Benazzouz 2037 ; Rey, Alot, dan Ramos 1984 ; Rodriguez-Roda 1966 ; Saber et al. 2019). Berdasarkan temuan berbagai ahli biologi lainnya, kami mengamati adanya perbedaan dalam musim kawin bonito Atlantik. Sebagai kesimpulan, kami dapat mendukung hipotesis bahwa periode reproduksi berbeda berdasarkan lokasi, spesies, dan kondisi lingkungan. Namun, meskipun hasilnya meyakinkan dan memberikan gambaran yang jelas tentang periode reproduksi bonito Atlantik di wilayah Samudra Atlantik ini, kami harus melakukan pengambilan sampel tambahan sepanjang tahun dan memasukkannya ke dalam model untuk sepenuhnya mengeksplorasi biologi reproduksi spesies ini di SEEZ, sehingga memperkuat kesimpulan kami.
4.1 Keterbatasan Penelitian
Kami menyadari bahwa kondisi lingkungan sangat memengaruhi siklus tahun-tahun terbaik bagi spesies pelagis. Oleh karena itu, sayangnya mustahil untuk mengetahui apakah tahun tertentu merupakan tahun terbaik untuk perekrutan atau tidak, dan karena kami tidak memiliki semua informasi, kami perlu menyelesaikan pertanyaan ini secara definitif. Ini adalah salah satu alasan utama kami tidak dapat menggunakan model ELEFAN dan vGBF untuk memperkirakan parameter populasi, termasuk perkiraan mortalitas dan eksploitasi dalam analisis kami. Memang, kami telah mengamati beberapa keterbatasan mendasar saat mengambil sampel atau mengumpulkan data. Tim survei perlu memberikan perhatian khusus pada kekurangan ini dan terlibat dalam pengumpulan informasi yang berkelanjutan dan ketat untuk mengatasi semua aspek yang berpotensi menimbulkan keraguan tentang keakuratan proses pengambilan sampel dan kelayakan data yang dikumpulkan. Meskipun demikian, meskipun upaya luar biasa sedang dilakukan meskipun ada kesulitan dan realitas sosial-ekonomi yang disebabkan oleh medan, umumnya karena dinamika sektor penangkapan ikan artisanal, yang sulit dikendalikan, ada elemen tertentu yang dapat ditingkatkan, seperti kurangnya informasi meteorologi, koordinat zona penangkapan ikan yang tepat, dan yang terpenting, metode pengumpulan data. Karena data merupakan dasar paling penting untuk memperkirakan hasil yang dapat diandalkan, data tersebut harus menjadi subjek studi dan pemantauan yang ketat dan teliti untuk mempertimbangkan semua informasi yang diperlukan dan, yang terpenting, informasi rutin untuk setidaknya satu siklus penuh dalam program survei pengumpulan data di masa mendatang.
5. Kesimpulan
Memahami dinamika populasi bonito Atlantik di Zona Ekonomi Eksklusif Senegal sangat penting untuk menilai status stok spesies ini secara akurat dan menerapkan praktik pengelolaan berkelanjutan di wilayah tersebut. Studi ini bertujuan untuk menyelidiki berbagai karakteristik biologis bonito Atlantik, termasuk parameter pertumbuhan (seperti a, b, L ∞ dan K ) dan aspek reproduksi. Hasil investigasi akan sangat penting untuk model penilaian stok yang menggunakan data yang ada. Ini akan meningkatkan ketepatan status stok bonito Atlantik di wilayah ini dan menyederhanakan proses pembentukan HCR yang efektif untuk studi selanjutnya. Strategi pengelolaan ini akan memainkan peran penting dalam mempromosikan keberlanjutan operasi penangkapan ikan skala kecil, terutama yang menargetkan tuna kecil. Selain itu, analisis lebih lanjut akan fokus pada penilaian demografi, evaluasi tekanan penangkapan ikan dan pengembangan langkah-langkah pengelolaan untuk memastikan konservasi jangka panjang dan penggunaan sumber daya bonito Atlantik yang berkelanjutan di Samudra Atlantik Timur Laut.


