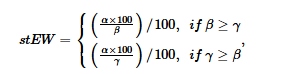ABSTRAK
Ada bukti yang berkembang mengenai peran hibridisasi introgresif dalam mendorong adaptasi spesies (yaitu, introgresi adaptif) karena meningkatnya studi genomik pada keragaman taksa selama beberapa dekade terakhir. Namun, hibridisasi introgresif dianggap, dan masih dianggap, sebagai proses penyeragaman yang menghambat proses evolusi adaptasi terhadap tekanan seleksi. Meskipun ada kemajuan metodologis, masih ada kesenjangan utama dalam memahami bagaimana introgresi adaptif karena hibridisasi berfungsi di seluruh kelompok taksonomi dan tingkat biologis. Studi ini memiliki tiga tujuan: (1) untuk mengeksplorasi tren historis dalam pemahaman introgresi adaptif, khususnya dimensi genomik dan fungsionalnya; (2) untuk menyelidiki karakteristik organisme struktural yang memengaruhi pola introgresi adaptif; dan (3) untuk mengevaluasi bagaimana introgresi adaptif berinteraksi dengan mekanisme evolusi yang berlawanan. Kami melakukan tinjauan sistematis terhadap literatur introgresi adaptif dan meta-analisis multidimensi. Tren pengetahuan saat ini telah dibentuk oleh revolusi genomik. Sejak 2012, studi genomik telah berkontribusi dalam membangun pemahaman yang lebih jelas tentang introgresi adaptif. Jumlah dan variasi studi yang dipublikasikan meningkat dari bakteri hingga mamalia di seluruh gradien kompleksitas, dengan fokus pada tingkat genomik dan secara progresif memiliki konsekuensi pada lebih banyak tingkat organisasi biologis (dari fisiologis dan demografis hingga perilaku/ekologis). Dengan menguji kecenderungan, studi kami juga mengungkap mekanisme evolusi yang terkait dengan introgresi adaptif yang terjadi bersamaan dengan gaya divergensi, yang menunjukkan bahwa proses ini tidak saling eksklusif, bahkan ketika mereka bertindak dalam arah yang berlawanan, yaitu, konvergensi dan divergensi, seperti introgresi autosom (versus pulau diferensiasi dalam kromosom terkait jenis kelamin), seleksi penyeimbang (versus pergeseran genetik), atau seleksi seksual (versus perkawinan asortatif). Keseimbangan ini dimediasi oleh kondisi lingkungan seperti yang sering dilaporkan dalam studi, terlepas dari kompleksitas struktural organisme, yang membentuk jalur proses evolusi spesies yang melakukan introgresi. Mempelajari pola introgresi memiliki implikasi penting untuk memahami adaptasi dalam lingkungan yang berubah dengan cepat.
1 Pendahuluan
Perkawinan silang yang berulang dapat menyebabkan spesies yang berhubungan dekat berbagi materi genetik (termasuk DNA nuklir [nDNA] dan/atau DNA sitoplasma [cDNA]) melalui introgresi (Arnold 2004 ; Hedrick 2013 ; Powell et al. 1999 ). Proses ini secara historis dianggap sebagai proses homogenisasi (Arnold 1992 ; Latch et al. 2006 ), yang menangkal tekanan seleksi lokal dan mengurangi kebugaran populasi yang berbeda dengan memperkenalkan alel di luar rentang adaptif lokal (Bolnick dan Nosil 2007 ; Maguilla et al. 2017 ). Akibatnya, hal ini dianggap sebagai masalah konservasi karena risiko bertindak melawan divergensi dan menyebabkan genangan genetik (lihat Tabel 1 — Glosarium), yang dapat memengaruhi kelangsungan hidup spesies dalam jangka panjang (Maguilla et al. 2017 ). Mengikuti alasan ini, beberapa penulis menganggap introgresi sebagai kerusakan pada integritas spesies, proses maladaptif, atau kemalangan alam yang tidak diharapkan (Llopart et al. 2014 ).
TABEL 1. Glosarium.
Introgresi adaptif
Transfer materi genetik secara alami melalui pembiakan interspesifik dan persilangan balik hibrida dengan spesies induk yang diikuti oleh seleksi berdasarkan alel yang diintrogres (de Lafontaine et al. 2015 ; Song et al. 2011 )
Perkawinan asortatif
Perilaku perkawinan dimana individu menunjukkan preferensi yang lebih tinggi untuk kawin dengan individu yang memiliki fenotipe dan genotipe yang serupa dengan yang diharapkan pada pola perkawinan acak (Herbers 2010 )
Menyeimbangkan pemilihan
Seperangkat tekanan selektif yang secara aktif memungkinkan pemeliharaan beberapa alel dalam kumpulan gen populasi pada frekuensi yang lebih besar daripada yang diharapkan berdasarkan efek pergeseran genetik saja (King et al. 2006 )
Organisasi biologis
Organisasi hierarkis struktur dan sistem biologis yang kompleks (Mazzocchi 2008 )
Pergeseran genetik
Perubahan frekuensi alel yang ada dalam kumpulan genetik suatu populasi sebagai akibat dari pengambilan sampel organisme secara acak (Bolnick et al. 2011 )
Rekombinasi genetik
Pertukaran genetik antara taksa yang berbeda yang menghasilkan kombinasi sifat baru yang berbeda dari sifat yang ditemukan pada spesies induk (Martinsen et al. 2001 )
Penenggelaman genetik
Aliran gen dari spesies yang paling melimpah menuju spesies dengan ukuran populasi yang lebih kecil dapat menyebabkan depresi perkawinan sedarah karena penggantian genotipe lokal (Todesco et al. 2016 )
Introgresi genomik
Distribusi alel introgress di seluruh genom (Gompert dan Buerkle 2009 )
Introgresi geografis
Distribusi geografis alel di seluruh zona hibrida (Payseur et al. 2004 )
Aturan Haldane
Seks heterozigot menunjukkan penurunan kebugaran hibrida yang lebih besar (Haldane 1922 )
Pulau-pulau diferensiasi
Pulau-pulau diferensiasi adalah wilayah genom yang menunjukkan tingkat diferensiasi yang luar biasa tinggi antara populasi atau spesies (Nachman dan Payseur 2012 ) yang dapat terlibat langsung dalam proses isolasi dan divergensi reproduksi atau tidak (Nosil et al. 2012 )
Hibridisasi alami
Kontak perkembangbiakan sekunder antara dua populasi yang berevolusi secara terpisah selama kurun waktu tertentu tetapi dalam kondisi alami (Neri et al. 2017 )
Sapuan selektif
Proses genetika yang melaluinya alel baru yang meningkatkan kebugaran individu meningkat frekuensinya dalam populasi dan menjadi tetap (frekuensi 1) karena seleksi alam (Bay dan Ruegg 2017 ; Kuhlwilm et al. 2019 )
Seleksi seksual
Perilaku perkembangbiakan yang mendukung pasangan kawin dengan keunggulan adaptif (kebugaran lebih tinggi), terlepas dari spesiesnya (Parrett dan Knell 2018 )
Segregasi transgresif
Sifat fenotipik yang ekstrim di luar rentang fenotipik asli spesies induk dihasilkan melalui introgresi dan mengarah pada spesiasi hibrida (Nichols et al. 2015 ; Rieseberg et al. 2003 )
Di sisi lain, sejak publikasi esai teoritis pertama tentang topik tersebut, peran introgresi menjadi subjek kontroversi karena alel yang diintrogresi dapat ditemukan di bagian genom yang sedang diseleksi serta daerah netral, dan dapat berkontribusi pada adaptasi, tidak memiliki dampak, atau dihilangkan karena merusak (penekanan introgresi) (Seixas et al. 2018 ). Introgresi dianggap netral ketika alel yang diintrogresi tidak memiliki konsekuensi fenotipik/fisiologis yang dapat memengaruhi kebugaran garis keturunan/spesies reseptor (Matosiuk et al. 2014 ). Jika tidak, dianggap maladaptif jika memiliki hasil adaptif negatif dengan mengurangi kebugaran atau kelangsungan hidup garis keturunan evolusi di lingkungannya (Minder dan Widmer 2008 ). Faktor-faktor seperti perubahan dalam ukuran populasi, sistem perkawinan, dan tingkat rekombinasi dapat memiliki hasil yang berbeda dan memengaruhi akumulasi variasi genetik yang merusak dalam populasi alami. Variasi yang merusak—baik resesif maupun aditif—dapat mengacaukan deteksi introgres adaptif (Kim et al. 2018 ).
Namun, bertentangan dengan asumsi historis bahwa introgresi terutama berdampak netral (Van Valen 1976 ; Räsänen dan Hendry 2008 ), bukti baru-baru ini mendukung bahwa alel yang menguntungkan lebih mudah mengalami introgresi daripada yang netral (Harrison dan Larson 2014 ). Proses acak, seperti pergeseran genetik (Gompert et al. 2013 ; Wade dan Goodnight 2006 ) dapat menghilangkan alel maladaptif dan merusak dalam beberapa generasi atau mempertahankan yang lain dengan kecenderungan untuk bertahan sebagai netral dan bergantung pada demografi, mengikuti proses seleksi di mana alel maladaptif dan merusak cenderung dihilangkan. Alel netral hanya tunduk pada pergeseran genetik, sementara alel yang menguntungkan cenderung meningkat frekuensinya (Pease et al. 2016 ). Proses selektif ini menyebabkan introgres adaptif (lihat Tabel 1 — Glosarium) semakin didokumentasikan sebagai kekuatan pendorong adaptif utama yang mengarah pada adaptasi yang lebih cepat daripada mutasi de novo (Pfeifer dan Kapan 2019 ).
Semakin banyak bukti telah menunjukkan bahwa adaptasi terhadap kondisi baru dapat terjadi melalui introgresi alel yang menguntungkan (Jones et al. 2018 ; Lexer et al. 2004 ; Nadachowska-Brzyska et al. 2012 ). Introgresi adaptif meningkatkan kapasitas adaptif dan mendorong lompatan evolusi, melewati tahap evolusi antara (Acosta dan Premoli 2018 ). Dengan demikian, adaptasi yang lebih cepat dapat terjadi ketika variasi genetik yang ada ini berasal dari spesies lain yang lebih baik beradaptasi dengan kondisi lingkungan. Mekanisme ini dapat meningkatkan kelangsungan hidup spesies (Liu et al. 2015 ; Norris et al. 2015 ), mendorong perluasan jangkauan mereka (Aleksic et al. 2018 ), dan bahkan mendukung penyelamatan evolusi (Aeschbacher et al. 2017 ). Paradoksnya, dalam beberapa keadaan, introgres adaptif bahkan dapat menyebabkan divergensi spesies (Alexander et al. 2017 ; Antelope et al. 2017 ), misalnya, dengan mendorong spesiasi hibrida melalui segregasi transgresif (Arnold et al. 2012 ).
Karena langka, alel introgres yang menguntungkan dapat disukai, misalnya, dengan menyeimbangkan seleksi (Nadachowska-Brzyska et al. 2012 ), dan dengan cepat terbentuk dalam spesies reseptor melalui sapuan selektif (Bay dan Ruegg 2017 ; Kuhlwilm et al. 2019 ). Tidak seperti mutasi baru, yang dimulai dengan prevalensi 1/2 N, alel introgres mungkin memiliki prevalensi “awal” yang lebih tinggi, tergantung pada tingkat hibridisasi (Pfeifer dan Kapan 2019 ). Dengan demikian, prevalensi awal yang tinggi dari alel introgres juga dapat memfasilitasi fiksasinya, terlepas dari efek kebugaran. Setelah menetap secara stabil, alel-alel ini dapat bertindak sebagai sumber penting variasi genetik dan fenotipik baru di beberapa titik dalam sejarah evolusi (Baldassarre et al. 2014 ; Cahill et al. 2015 ). Hanya satu gen yang diintrogresikan dapat memiliki potensi untuk mendorong variasi dan akibatnya memiliki dampak ekologi yang besar (Bradshaw dan Schemske 2003 ).
Munculnya pendekatan sekuensing throughput tinggi mengakibatkan pertumbuhan eksponensial bukti ilmiah yang mendokumentasikan terjadinya introgresi dan, pada tingkat yang lebih rendah, konsekuensi adaptif dan evolusionernya (Taylor dan Larson 2019 ). Dari kumpulan literatur global, introgresi adaptif menonjol sebagai proses yang sangat kompleks yang bekerja melintasi proses divergensi dalam semua kelompok taksonomi yang mencakup berbagai tingkat kompleksitas struktural, termasuk bakteri (Palmer et al. 2010 ), protista (Nader et al. 2019 ), fungi (Dunn et al. 2013 ), tumbuhan, dari bryophyta (James et al. 2008 ) hingga angiospermae (Taylor et al. 2009 ), dan hewan, baik invertebrata (Norris et al. 2015 ) dan vertebrata (Liu et al. 2015 ). Tekanan lingkungan, baik alami (Matosiuk et al. 2014 ) dan antropogenik (Pardo-Diaz et al. 2012 ), mendorong introgres adaptif pada tingkat genomik, yang menyebabkan konsekuensi pada berbagai tingkat organisasi biologis (lihat Tabel 1 — Glosarium), dari fisiologi (Storchová et al. 2004 ) hingga perilaku (McDonald et al. 2001 ) dan demografi (Ray dan Excoffier 2009 ). Memahami bagaimana introgres adaptif dapat memainkan peran penting sebagai kekuatan evolusi akan memerlukan tinjauan luas tentang status pengetahuan tentang proses introgres adaptif di seluruh pohon kehidupan dan dalam semua dimensi biologisnya (Pfennig 2021 ). Tinjauan sistematis dan meta-analisis menggabungkan dan mengevaluasi kumpulan data besar, memberikan wawasan tentang tren dan kesenjangan pengetahuan, menyediakan landasan untuk penelitian masa depan tentang adaptasi di lingkungan yang berubah dengan cepat (O’Dea et al. 2021 ). Protokol standar tinjauan sistematis memungkinkan reproduktifitas dan mencegah pengambilan keputusan subjektif selama proses tinjauan (Page et al. 2021 ; Shamseer et al. 2015 ). Meta-analisis melalui pendekatan jaringan multidimensi menyediakan kerangka kerja integratif dan holistik untuk menggabungkan data biologis dari tinjauan sistematis standar (Bradley et al. 2005 ). Meta-analisis dapat membantu memperluas pemahaman tentang dinamika evolusi yang mendorong adaptasi spesies di berbagai tingkat organisasi biologis.
Karena penelitian tentang introgres adaptif berkembang pesat, tinjauan sistematis diperlukan untuk mensintesis dan mengevaluasi kumpulan bukti yang berkembang ini. Studi ini bertujuan untuk memeriksa dimensi biologis dari introgres adaptif melalui meta-analisis jaringan multidimensi. Karakteristik intrinsik organisme—seperti organisasi seluler (prokariotik atau eukariotik), keberadaan kromosom tertaut-kelamin, dan pola seleksi seksual—kemungkinan memainkan peran penting dalam membentuk sifat adaptif yang timbul dari introgres. Demikian pula, arsitektur genom memainkan peran mendasar dalam membentuk kemungkinan introgres adaptif. Fitur genom struktural, terutama dalam hal ketidakcocokan genetik dan variasi struktural, seperti inversi kromosom, translokasi, dan fusi, dapat bertindak sebagai penghalang aliran gen dengan mengurangi rekombinasi atau mendorong isolasi reproduktif (Hoffmann dan Rieseberg 2008 ; Feder et al. 2012 ).
Introgres adaptif tidak terjadi secara terisolasi; ia diimbangi dan dimodulasi oleh mekanisme evolusi lain yang terkait dengan divergensi spesies, termasuk ketidakcocokan genetik dan perkawinan asortatif. Misalnya, pulau-pulau diferensiasi—didefinisikan sebagai daerah genom yang menunjukkan tingkat diferensiasi yang luar biasa tinggi antara populasi atau spesies (Nachman dan Payseur 2012 )—dapat bertindak sebagai penghalang aliran gen, baik dengan memperkuat isolasi reproduksi atau dengan menyediakan lingkungan selektif yang membatasi atau meningkatkan penyebaran alel yang diintrogres. Untuk mengeksplorasi dinamika ini, kami mendefinisikan tiga tujuan utama: (1) untuk mengeksplorasi tren historis dalam pemahaman introgres adaptif, khususnya lokalisasi seluler dan genomiknya, transmisi terarah, dan korelasi potensial dengan mekanisme evolusi lain, seperti pergeseran genetik atau perkawinan asortatif; (2) untuk menyelidiki bagaimana karakteristik organisme, termasuk unit organisasi struktural dan fungsionalnya (baik prokariotik atau eukariotik), serta ada/tidaknya kromosom tertaut kelamin dan strategi perkawinan, memengaruhi pola pada sifat yang terkait dengan introgres adaptif; (3) mengevaluasi interaksi antara introgres adaptif dan mekanisme evolusi utama yang timbul dari divergensi yang secara bersamaan dapat bertindak sebagai kekuatan penyeimbang.
Untuk mengatasi tujuan-tujuan ini, kami menguji hipotesis berikut: (1) Kemajuan dalam genomik telah meningkatkan pemahaman kita tentang introgres adaptif secara signifikan, yang mengarah pada pola pengetahuan yang lebih stabil dan lebih jelas dalam studi-studi terkini, di berbagai kelompok taksonomi dan tingkat biologis; (2) Organisme yang lebih kompleks—secara genetik, fisiologis, dan perilaku—diharapkan menunjukkan pola introgres adaptif yang lebih rumit dan beragam di berbagai tingkat biologis dibandingkan dengan organisme yang lebih sederhana; (3) Kekuatan introgres adaptif dan divergensi dapat hidup berdampingan dan bertindak secara bersamaan dalam populasi, bahkan ketika proses-proses ini mendorong hasil evolusi yang berlawanan (misalnya, konvergensi versus divergensi). Mengklarifikasi tujuan-tujuan ini dan menguji hipotesis-hipotesis ini dapat memberi kita pandangan-pandangan baru yang disintesis tentang bagaimana introgres dapat bertindak sebagai kekuatan evolusi yang membentuk jalur-jalur evolusi dan menawarkan implikasi-implikasi terapan yang krusial untuk memahami adaptasi dalam lingkungan-lingkungan yang berubah dengan cepat.
2 Bahan dan Metode 2.1 Penelusuran Literatur
Kami melaksanakan telaah sistematis dengan mencari studi yang menilai pola introgresi yang dihasilkan dari adaptasi. Mengikuti protokol telaah PRISMA standar (Moher et al. 2009 ; O’Dea et al. 2021 ), kami menggunakan ISI Web of Knowledge, Scopus, Google, dan Google Scholar sebagai mesin pencari (Falagas et al. 2008 ). Kami memilih sekumpulan istilah pencarian (lihat Informasi Pendukung untuk detailnya) yang terkait dengan hibridisasi alami (lihat Tabel 1 — Glosarium), genomik, introgresi, dan adaptasi, dengan sengaja menghindari istilah yang terkait dengan intervensi manusia langsung atau tidak langsung (misalnya, pembiakan atau manipulasi genetik), seperti domestikasi dan skenario invasif, membatasi pencarian pada introgresi genomik alami (lihat Tabel 1 — Glosarium) dengan konsekuensi adaptif yang potensial. Kata kunci dipilih dari daftar kata kunci dalam makalah referensi dan melalui konsultasi tim ahli dalam biologi evolusi. Untuk memperoleh jumlah publikasi terbanyak mengenai topik tersebut, kumpulan kata kunci terpilih terakhir mencakup istilah dan ekspresi yang paling umum dan tidak ambigu (lihat Informasi Pendukung untuk detail pencarian). Pencarian mencakup periode dari tahun 1900 hingga November 2019 dan dibatasi pada bahasa Inggris.
Pencarian kami menghasilkan 820 publikasi. Dari kumpulan data ini, kami menghapus duplikat dan mengecualikan esai teoritis, teks opini, makalah yang tidak ditinjau sejawat, dokumen di luar topik, dan jurnal non-ISI. Kemudian kami memeriksa daftar referensi dari publikasi yang tersisa untuk mencari karya tambahan yang menyertakan istilah yang diteliti dan tidak dihitung dalam pencarian awal. Dari kumpulan publikasi global ini, kami mengecualikan semua tinjauan dan bab buku. Baik judul maupun abstrak dari 436 artikel yang tersisa dibaca untuk menilai relevansinya dengan penelitian, berdasarkan: (1) apakah proses introgresi adaptif merupakan hasil dari proses alami, dan (2) apakah introgresi adaptif diidentifikasi oleh penulis sebagai penjelasan paling masuk akal di balik proses adaptif yang diamati. Kumpulan data akhir terdiri dari 358 penelitian (Gambar S1 — Informasi Pendukung ). Semua makalah dianalisis secara lengkap oleh penulis pertama.
2.2 Tinjauan Sistematis
Kami mengekstrak informasi bibliografi dari 358 studi, termasuk periode temporal dan lokasi spasial fenomena introgres adaptif yang dilaporkan dalam setiap studi, taksa yang menjalani proses introgres, dan metode studi utama (jenis eksperimen—termasuk introgres yang berpotensi diidentifikasi tanpa data molekuler dengan metode seperti eksperimen persilangan—jenis penanda introgres molekuler, dan apakah sekuensing genom utuh dilakukan). Akhirnya, kami mencatat karakteristik utama introgres yang dirujuk dalam setiap studi dan hasil adaptif terkait. Informasi terperinci tentang variabel yang diekstraksi tersedia di Tabel S1 ( Informasi Pendukung ). Penting untuk mempertimbangkan bahwa tidak selalu pasti bahwa alel/sifat diintrogres secara adaptif. Oleh karena itu, kami mendasarkan kesimpulan ini pada interpretasi penulis sebagaimana dinyatakan dalam kesimpulan studi, dan pada bukti yang dijelaskan tentang gen dan/atau sifat bersama yang berpotensi memiliki hasil adaptif.
2.3 Meta-Analisis 2.3.1 Perubahan Temporal dalam Publikasi
Perubahan temporal dalam jumlah makalah yang diterbitkan diuji melalui General Linear Models (GLM) oleh Expert Modeler Methods yang tersedia pada perangkat lunak IBM SPSS Statistics (v. 22; SPSS Inc., Chicago, IL). Pendekatan ini dilengkapi dengan regresi linear, yang dinilai melalui koefisien determinasi, antara jumlah makalah yang diterbitkan dan waktu. Kami mendefinisikan titik kritis sebagai momen temporal ketika garis tren eksponensial berhenti menunjukkan kemiringan yang berkurang (Maroco 2010 ).
2.3.2 Pengaruh Karakteristik Struktural Organisme terhadap Pengetahuan Introgresi Adaptif
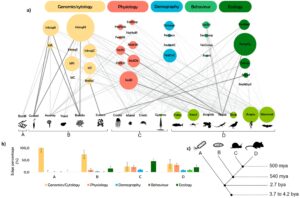
Kami mengukur tingkat kompleksitas di seluruh kelompok taksonomi dengan menjumlahkan klasifikasi biner untuk tiga karakteristik penting dalam sejarah evolusi mereka. Secara khusus, kami menetapkan nilai biner (0 atau 1) untuk setiap kelompok berdasarkan kriteria berikut: unit organisasi struktural dan fungsional (prokariotik = 0; eukariotik = 1), ada atau tidaknya kromosom tertaut kelamin (tidak ada = 0; ada = 1), dan strategi perkawinan yang dominan (acak = 0; seleksi seksual = 1). Metode ini memberikan metrik standar untuk menilai kompleksitas, seperti yang dirinci dalam Tabel S2 ( Informasi Pendukung ). Kami kemudian mengatur kelompok-kelompok ini menurut tingkat kompleksitas genetik dan perilaku yang dikuantifikasi, yang juga kemungkinan berkorelasi dengan kompleksitas fisiologis dan ekologis, menciptakan gradien kompleksitas taksonomi yang tersedia pada Gambar 2. Contoh kelompok taksonomi kompleks termasuk spesies eukariotik dengan kromosom tertaut kelamin dan mekanisme perilaku kompleks seleksi seksual, seperti mamalia dan angiospermae. Kami melakukan analisis jaringan bipartit menggunakan grafik sumber terbuka viz. platform Gephi 0.10 ( https://gephi.org/ ) tentang karakteristik introgres adaptif yang dilaporkan dalam berbagai tingkat organisasi biologis (Genomik/sitologi—Lokasi introgres/pulau diferensiasi dalam genom/sel; Fisiologi—Kendala fisiologis pada pola introgres adaptif; Demografi—Dampak demografi pada arah dan jumlah/pola introgres adaptif; Perilaku—Dampak perilaku pada pola introgres adaptif; dan Ekologi—Pengaruh ekologis pada pola introgres adaptif) dan kelompok taksonomi di seluruh gradien kompleksitas taksonomi.
Signifikansi faktor Kelompok Taksonomi (A—Prokariota, B—Eukariota sel tunggal, C—Eukariota dengan kromosom tertaut kelamin dan pasangan acak, dan D—Eukariota dengan seleksi seksual) pada persentase koneksi di lima tingkat organisasi biologis (“Genomik/sitologi,” “Fisiologi,” “Demografi,” “Perilaku,” dan “Ekologi”) dinilai menggunakan MANOVA, setelah memvalidasi asumsi normalitas dengan uji Kolmogorov–Smirnov ( p > 0,05 untuk semua kelompok). Asumsi homogenitas varians-kovarians diverifikasi dengan uji Levene. Analisis statistik dilakukan dengan menggunakan perangkat lunak SPSS. Ketika MANOVA mendeteksi efek signifikan, ANOVA dilakukan untuk setiap variabel, diikuti oleh uji post hoc HSD Tukey (Tabel S3 — Informasi Pendukung ).
2.3.3 Gaya Introgresi dan Divergensi Adaptif—Pendekatan Jaringan Multilayer
Kami melakukan analisis multilayer melalui platform visualisasi dan analisis jaringan multilayer sumber terbuka MuxViz ( http://muxviz.net/ ). MuxViz adalah pendekatan untuk merepresentasikan berbagai macam sistem yang kompleks, seperti struktur biologis, pada berbagai tingkat organisasi biologis, menggunakan jaringan genetik, neuronal, dan transportasi empiris (De Domenico et al. 2014 ). Grafik jaringan multilayer dapat divisualisasikan dengan membuat hubungan antara dua atau lebih variabel yang diekstraksi, sebagai node, yang menggabungkannya ke dalam berbagai tingkat (lapisan) (De Domenico et al. 2014 ). Hubungan ini terjadi ketika dua karakteristik studi kasus (dua variabel yang diekstraksi) secara simultan dijelaskan dalam makalah yang sama yang, tergantung pada apakah mereka saling bergantung atau tidak, dapat diarahkan atau tidak diarahkan. Di sini, lapisan grafik dirancang untuk merepresentasikan berbagai tingkat organisasi biologis: (1) Genomik/sitologi; (2) Fisiologi; (3) Perilaku; (4) Demografi; dan (5) Ekologi. Kami menggunakan algoritma Large Graph Layout (LGL) pada dua dimensi dengan susunan node multipleks dan intersepsi sebagai metode templat, di mana semua node dapat dihubungkan (koneksi intra dan inter-layer) (De Domenico et al. 2014 ).
Bahasa Indonesia: Untuk menghitung properti jaringan khusus multilayer, kami melakukan algoritma sentralitas vektor eigen untuk menganalisis keserbagunaan vektor eigen melalui ekstensi multilayer, yang mengukur kepentingan setiap node (distandarisasi antara 0 dan 1) tergantung pada koneksinya di dalam dan lintas lapisan (De Domenico et al. 2014 ). Ukuran node diwakili oleh algoritma sentralitas PageRank (distandarisasi antara 0 dan 1) yang memeringkat karakteristik studi kasus (lihat Informasi Pendukung ) untuk mengukur kepentingan setiap node dalam jaringan dengan korelasi dengan metrik vektor eigen (De Domenico et al. 2015 ).
Untuk menstandardisasi intensitas koneksi antar node, kami menggunakan teknik normalisasi. Untuk analisis eksploratori, kami menghitung persentase berdasarkan pembagian jumlah koneksi yang diidentifikasi antara dua node dengan jumlah total tipe koneksi berbeda yang ada dalam basis data. Basis data ini mencakup total 210 edge yang berbeda, yang berfungsi sebagai keseluruhan semesta untuk analisis kami. Lebih jauh, kami merancang metode untuk menilai kekuatan koneksi antar node dengan memperkenalkan persamaan standardisasi Edge Weights. Persamaan ini mempertimbangkan ukuran node, yang diukur dari segi jumlah makalah yang terkait dengannya. Dengan menerjemahkan nilai ini ke dalam skala mulai dari 0 hingga 1, kami dapat mengukur intensitas koneksi dalam kaitannya dengan konteks spesifik pengetahuan terkini tentang kedua node yang terhubung. Edge weight, yang dilambangkan dengan edge thickness, mewakili jumlah total makalah di mana pola terhubung dijelaskan antara dua node. Hubungan ini dihitung sebagai standardized edge weight ( st EW ) menurut persamaan berikut:
di mana α merupakan simbol bobot tepi total yang diukur dari jumlah total makalah yang secara bersamaan mendeskripsikan dua karakteristik introgres adaptif; β merupakan simbol jumlah total makalah yang mendeskripsikan salah satu karakteristik; ϒ merupakan simbol jumlah total makalah yang mendeskripsikan karakteristik kedua.
Metrik stEW dibuat untuk tujuan perbandingan. Selain itu, menyajikan jumlah pasti makalah saat menilai hubungan antara dua simpul bertujuan untuk menawarkan informasi tambahan. Meskipun tidak terlalu berguna untuk membuat perbandingan, jumlah makalah memberikan wawasan berharga tentang pemahaman terkini tentang hubungan dan simpul tertentu.
3 Hasil 3.1 Evolusi Temporal dalam Tren Pengetahuan tentang Introgresi Adaptif
Jumlah publikasi tentang introgresi adaptif telah tumbuh secara eksponensial sejak 2012, didorong oleh kemajuan dalam pengurutan genom dan teknologi komputasi. Tren ini mencakup berbagai bidang, khususnya biologi evolusi, dengan peningkatan yang nyata dalam penelitian yang menggunakan pengurutan genom secara keseluruhan.
Kami hanya mencatat beberapa studi yang diterbitkan selama abad ke-20, dengan prevalensi tertinggi pada tahun 1990-an, yang melihat peningkatan dalam bukti yang diterbitkan untuk introgresi adaptif, khususnya dari studi eksperimental terkontrol botani yang melibatkan penggunaan lokus sifat kuantitatif (QTL) (Gambar S1 — Informasi Pendukung ). Sedikit peningkatan dalam jumlah publikasi antara tahun 2000 dan 2012 diamati bersamaan dengan meningkatnya perkembangan metodologis dari proyek Genom Manusia ( https://www.genome.gov/ ). Namun, lebih dari 75% dari studi yang dianalisis diterbitkan setelah tahun 2012, ketika jumlah rata-rata tahunan makalah yang diterbitkan meningkat dari 1,9 ± 3,1 (1966–2011) menjadi 34,5 ± 10,1 (2012–2019), yang setara dengan peningkatan tahunan rata-rata sebesar 1824,14% dari makalah (
= 97,37, p < 0,01).
Peningkatan tingkat publikasi ini merupakan hasil dari penerapan pendekatan genomik secara luas, terutama melalui munculnya high throughput sequencing (HTS) dan penggunaan whole-genome sequencing dan single nucleotide polymorphism (SNP) (Gambar S1 — Informasi Pendukung ). Teknik-teknik ini terbukti efektif dalam mendeteksi, dengan cara yang hemat biaya, area genomik yang mengalami introgresi dan nilai adaptifnya. Meningkat secara eksponensial seiring waktu (GLM_SNPs [family = “poisson”]: R 2 = 0,80, p < 0,05, GLM_Whole-Genome [family = “poisson”]: R 2 = 0,76, p < 0,05), teknik-teknik ini juga berkontribusi pada pertumbuhan eksponensial pengetahuan introgresi adaptif, yang terbukti dari jumlah kumulatif makalah yang diterbitkan dari waktu ke waktu (GLM_Total Number of Papers [family = “poisson”]: R 2 = 0,94, p < 0,05) (Gambar S1 — Informasi Pendukung ). Pola pertumbuhan ini terbukti dalam berbagai bidang ilmiah (Antropologi, Filogeni/Genealogi, Ilmu Tanaman, dll.), dan khususnya dalam Biologi Evolusi (Gambar S1 — Informasi Pendukung ).
Revolusi genomik memungkinkan pembentukan pemahaman yang lebih jelas dan pola yang stabil dalam tren pengetahuan, khususnya tentang karakteristik genomik, dampaknya pada fisiologi, serta arah introgres adaptif (Gambar 1 ). Pola-pola ini meliputi: (i) frekuensi yang lebih besar dari kejadian introgresi nuklir autosomal di samping pulau-pulau diferensiasi dalam kromosom terkait-jenis kelamin; (ii) kejadian proses seleksi penyeimbangan yang jauh lebih tinggi dibandingkan dengan pergeseran genetik, serta tren introgresi yang berlaku dari asli ke penjajah dibandingkan dengan arah sebaliknya; dan (iii) keseimbangan yang penting, dengan sekitar setengah dari makalah yang diterbitkan setiap tahun menunjukkan beberapa derajat kemandulan dalam garis keturunan yang terlibat dalam introgresi, sementara separuh lainnya melaporkan tidak adanya kemandulan (Gambar 1 ). Sebaliknya, tidak ada pola yang jelas dalam aspek perilaku (Gambar 1 ). Khususnya, setelah tahun 2012, terdapat peningkatan yang signifikan dalam bukti yang mengindikasikan keberadaan kekuatan yang berbeda, seperti pulau-pulau diferensiasi, pergeseran genetik, atau perkawinan asortatif, dalam studi yang dianalisis mengenai introgres adaptif (Gambar 1 ).
Perkembangan temporal tren pengetahuan menurut persentase makalah yang diterbitkan per tahun yang memberikan bukti karakteristik spesifik introgresi adaptif. Garis putus-putus menunjukkan titik kritis yang mewakili awal revolusi genomik dalam studi introgresi adaptif.
3.2 Kompleksitas Struktur Organisme dan Karakteristik Introgresi Adaptif
Kompleksitas organisme memengaruhi pola introgres adaptif. Proses tingkat genomik mendominasi pada prokariota, sementara faktor perilaku dan ekologi memperoleh kepentingan pada eukariota yang lebih kompleks, khususnya yang memiliki mekanisme seleksi seksual (Gambar 2 ). Meta-analisis kami terhadap tren dalam literatur yang diterbitkan menunjukkan tiga karakteristik utama yang tampaknya mengkondisikan introgres adaptif yang dijelaskan: sel eukariotik, keberadaan kromosom seks, dan pengembangan mekanisme perilaku seleksi seksual (lihat Tabel 1 — Glosarium). Genom/sitologi mencakup hampir setengah dari total koneksi antara kelompok taksonomi dan klaim introgres adaptif (46,7% ± 30,17%). Fisiologi mencakup sekitar 20% koneksi (20,1 ± 16,88), diikuti oleh aspek demografis (6,3% ± 6,69%). Perilaku hanya mewakili 3,5% ± 5,81% dari total koneksi (Gambar 2 ). MANOVA mengungkapkan bahwa faktor Kelompok Taksonomi memiliki efek signifikan pada persentase koneksi di lima tingkat organisasi biologis (Jejak Pillai = 1,390; F (15,36) = 2,072; p = 0,037; Power = 0,887). Mengikuti signifikansi multivariat, uji post hoc HSD Tukey menunjukkan bahwa perbedaan signifikan terutama diamati dalam lapisan genomik/sitologi, dengan koneksi yang jauh lebih tinggi pada organisme uniseluler dan koneksi yang jauh lebih rendah pada eukariota multiseluler, terlepas dari strategi reproduksinya. Khususnya, eukariota dengan seleksi seksual menunjukkan koneksi perilaku yang jauh lebih tinggi dibandingkan dengan semua kelompok lainnya (Tabel S3 — Informasi Pendukung ).
Jaringan bipartit dari karakteristik introgres adaptif yang dilaporkan, dalam berbagai tingkat organisasi biologis, dan kelompok taksonomi, melintasi gradien kompleksitas taksonomi (ukuran simpul menunjukkan jumlah total kertas yang menjelaskan setiap karakteristik dan ketebalan tepi menunjukkan bobot tepi standar (stEW) dari setiap koneksi (a). Bagan batang yang menunjukkan efek faktor Kelompok Taksonomi (A—Prokariota, B—Eukariota sel tunggal, C—Eukariota dengan kromosom tertaut kelamin dan perkawinan acak, dan D—Eukariota dengan seleksi seksual) pada persentase koneksi di seluruh tingkat organisasi biologis. Batang menunjukkan nilai rata-rata dengan deviasi standar. Hasil signifikan dari beberapa perbandingan rata-rata terperinci disediakan dalam Tabel S3 ( Informasi Pendukung ) (b). Kladogram dengan momen pembelahan evolusioner potensial dari fungsionalitas introgres adaptif (c). Lapisan genomik/sitologi (Introgres autosom [IntrogA], Pulau diferensiasi autosom [IslA], Introgresi Nuklir [IntrogN], Introgresi pada Kromosom Seks [IntrogS], Introgresi Sitoplasma [IntrogC], Pulau-pulau diferensiasi dalam nukleus [IslN], Pulau-pulau diferensiasi pada Kromosom Seks [IslS], Pulau-pulau diferensiasi Sitoplasma [IslC], dan keberadaan seleksi penyeimbang [BalSel]); Lapisan Fisiologi (dampak endogen pada introgresi adaptif [EndImp], kepatuhan aturan Haldane [HalRule], pengecualian terhadap aturan Haldane [NoHalR], infertilitas betina [FemStl], infertilitas jantan [MalStl], semua infertilitas [AnStl], introgresi bias jenis kelamin [SexBI], arah introgresi bias paternal [PatDir], dan arah introgresi bias maternal [MatDir]); Lapisan Demografi (demografi kesenjangan [Demogr], kesenjangan demografi [NoDem], arah introgres dari penduduk asli ke penjajah [NatToC], dan pengecualian dari arah penduduk asli ke penjajah [NoNatC]); Lapisan perilaku (adanya seleksi seksual [SexSel], ada perkawinan asortatif [Assort], ada persaingan antarspesies intraseksual [SexComp], dan tidak adanya persaingan antarspesies intraseksual [NoComp]); dan Lapisan ekologi (introgresi yang dimediasi lingkungan [EnvMed], iterasi antara periode alopatrik dan simpatrik sepanjang sejarah evolusi [TempCy], dampak eksogen pada introgresi adaptif [ExImp], penjelajahan alel [AlSurf], dan tidak ada penjelajahan alel [NoAlSf]).Pada organisme yang secara evolusi lebih tua dan kurang kompleks, yaitu pada prokariota, introgres adaptif ditangani pada tingkat genom, yang mewakili satu-satunya lapisan yang ada di sebagian besar jaringan prokariota ( st EW = 0,11). Untuk spesies eukariotik bersel tunggal yang lebih tua, seperti protista, genom/sitologi mewakili 73,3% ± 25,3% dari semua koneksi dengan karakteristik introgres adaptif yang dipelajari. Berbeda dengan prokariota, penelitian pada kelompok ini menunjukkan fokus yang lebih jelas pada aspek sitologi introgres adaptif, seperti posisi pulau introgres di dalam sel (nukleus versus sitoplasma) (Gambar 2 ).
Reproduksi seksual pada spesies eukariotik multiseluler dengan kromosom seks heteromorfik membawa tantangan baru pada penyelidikan introgresi adaptif. Untuk eukariota yang lebih kompleks ini, penelitian menunjukkan penekanan yang jelas pada aspek fisiologis, yang selaras erat dengan signifikansi genom dan sitologi dalam hal persentase koneksi (21,5% ± 18,3%, st EW [fisiologi] = 0,07; 23,0% ± 19,5%, st EW [genom/sitologi] = 0,08, masing-masing), khususnya dalam penelitian pada krustasea (44,4%, st EW = 0,15), remis (24,0%, st EW = 0,10) dan gymnospermae (17,7%, st EW = 0,05). Pada eukariota multiseluler yang lebih tua ini dengan kromosom seks yang berbeda, introgresi adaptif sering dipengaruhi oleh faktor demografi. Setelah reproduksi menjadi stokastik di zona kontak, asimetri demografi ditunjukkan dalam literatur untuk menjadi mekanisme fundamental untuk mempromosikan introgresi terarah dari spesies yang paling melimpah ke yang paling sedikit melimpah. Gymnospermae mungkin adalah contoh yang paling menonjol dari ini (20,6%, st EW = 0,13). Sebaliknya, pada spesies eukariota yang lebih baru dan kompleks seperti burung atau mamalia (secara genomik dan perilaku), studi introgresi adaptif cenderung membahas masalah yang terkait dengan perilaku seksual (Gambar 2 ). Eukariota dengan preferensi kawin adalah satu-satunya yang karakteristik perilakunya ditunjukkan untuk mempengaruhi kemungkinan introgresi adaptif yang berhasil (8,4% ± 6,4%, st EW = 0,15). Pola ini sangat jelas dalam studi burung dan reptil (12,1%, st EW = 0,21 dan 21,5%, st EW = 0,33, berturut-turut).
Akhirnya, penelitian secara konsisten menyoroti pengaruh faktor ekologi (misalnya, paparan pestisida, perubahan suhu) pada introgres adaptif, terlepas dari kompleksitas taksonomi spesies, dari eukariota bersel tunggal (15,0% ± 22,4%, st EW = 0,06), eukariota yang lebih tua dengan kromosom tertaut seks dan pilihan pembiakan stokastik (45,5% ± 39,2%, st EW = 0,22) hingga eukariota yang mampu melakukan seleksi seksual (20,1% ± 8,2%, st EW = 0,18) (Gambar 1 ).
3.3 Mekanisme Evolusi dan Karakteristik Introgresi Adaptif yang Diketahui
Introgresi adaptif pada spesies eukariotik sebagian besar terjadi pada DNA nuklir, sering kali didorong oleh seleksi keseimbangan dan, tampaknya, tidak bergantung pada waktu kejadian introgresi. Pada spesies dengan kromosom terkait jenis kelamin, di mana introgresi lebih jarang terjadi, sering kali sejalan dengan aturan Haldane, yang biasanya menunjukkan pola bias jenis kelamin yang terkait dengan kemandulan atau ketidakmampuan untuk bertahan hidup. Pilihan pasangan acak atau seleksi seksual memengaruhi introgresi terarah, dengan asimetri demografis sering kali mendukung aliran gen penduduk asli ke penjajah dan membentuk dinamika bias jenis kelamin. Faktor lingkungan, baik alami maupun antropogenik, juga memainkan peran penting, sering kali dimediasi oleh siklus divergensi historis.
3.3.1 Introgresi Adaptif pada Spesies Eukariotik
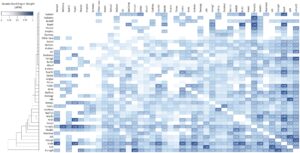
Kami menemukan penelitian yang menunjukkan bukti alel introgres dengan efek adaptif baik dalam DNA sitoplasma (cDNA) maupun DNA nuklir (nDNA), baik secara eksklusif maupun bersamaan ( st EW = 0,43, N = 31 makalah) (Gambar 3 ), meskipun ada beberapa kasus ketidakcocokan genetik, termasuk ketidaksesuaian sitonuklir (stEW = 0,38, N = 12) (Gagnaire et al. 2012 ). Namun, banyak kasus yang dijelaskan menunjukkan tingkat introgresi sitoplasma yang lebih rendah (atau bahkan tidak ada), terutama jika dibandingkan dengan introgresi nuklir (Brown et al. 2019 ; Carling dan Brumfield 2008 ; Hulsey et al. 2013 ; Morales et al. 2017 ). Hasil penelitian kami menunjukkan bahwa introgresi nuklir bukan hanya jenis transfer gen introgresi yang paling sering dilaporkan dalam genom eukariotik, tetapi juga karakteristik introgresi yang paling umum di antara semua penelitian yang dipublikasikan (Eigenvector = 1,00, PageRank = 0,98, N = 144 paper) (Gambar 4 ). Namun, kami tidak menemukan hubungan antara waktu introgresi (peristiwa kuno versus peristiwa terkini) dan lokasi genom introgresi (cDNA versus nDNA) (
= 3,46; p > 0,05). Seleksi keseimbangan lebih lazim terjadi dalam studi introgresi adaptif dibandingkan pergeseran genetik (Vektor eigen = 0,11, PageRank = 0,26, N = 32 makalah) (
= 13,23; p < 0,01), yang terutama berdampak pada introgresi nuklir ( st EW = 0,50, N = 18 makalah) (Gambar 3 ).
Peta panas bobot tepi terstandarisasi berpasangan ( st EW) di antara karakteristik introgres adaptif yang dilaporkan dan kelompok taksonomi.
Metrik eigenvektor dan PageRank dari analisis jaringan multilayer yang mempertimbangkan nilai multilayer (di berbagai level) dan nilai agregat (tanpa mempertimbangkan level). Ukuran lingkaran menunjukkan rata-rata bobot tepi standar (stEW) antara berbagai karakteristik introgresi adaptif di antar-level (sisi kiri) dan intra-level (sisi kanan).
3.3.2 Introgresi Adaptif pada Spesies dengan Kromosom Terkait Jenis Kelamin
Hasil kami menunjukkan bahwa ada lebih sedikit kasus introgres adaptif yang dilaporkan dalam cDNA (Eigenvector = 0,40, PageRank = 0,34, N = 72 paper) daripada ke dalam autosom (Eigenvector = 0,90, PageRank = 0,74, N = 84 paper) (Carling dan Brumfield 2008 ). Namun, keduanya jauh lebih sering dilaporkan daripada yang ada dalam kromosom terkait seks (Eigenvector = 0,21, PageRank = 0,24, N = 17 paper). Introgresi dalam kromosom terkait seks tidak pernah dijelaskan secara bersamaan dengan introgresi cDNA, terjadi tepat bersamaan dengan introgresi autosom ( st EW = 0,53, N = 9 paper) (Gambar 3 ). Spesies dengan kromosom seks heteromorfik, khususnya mamalia, menunjukkan tingkat introgresi yang lebih rendah pada kromosom terkait seks dibandingkan dengan autosom (
= 19,14; p < 0,01).
Beberapa penelitian, yang mencakup banyak taksa, secara bersamaan telah mengidentifikasi pulau-pulau diferensiasi yang terutama terletak pada kromosom terkait kelamin, serta contoh-contoh introgresi autosomal ( st EW = 0,57, N = 36 makalah; Gambar 3 ). Meskipun demikian, terdapat pengecualian penting dalam lokasi genomik pulau-pulau diferensiasi (Autosom: Eigenvector = 0,37, PageRank = 0,34, N = 32). Hanya satu makalah yang memberikan bukti untuk pulau-pulau diferensiasi sitoplasma atau autosomal dan pergeseran genetik ( st EW = 0,13).
Kami menemukan makalah dengan bukti adanya pulau diferensiasi pada kromosom terkait jenis kelamin bersamaan dengan beberapa tingkat kemandulan/ketidakmampuan untuk bertahan hidup pada 28 makalah ( st EW = 0,51), yang sangat tinggi dibandingkan dengan kasus non-mandul ( st EW = 0,28, N = 13 makalah) (
= 4,23; p < 0,05). Kedua pulau diferensiasi pada kromosom terkait-jenis kelamin dan sterilitas/tidak dapat bertahan hidup seharusnya memengaruhi jenis kelamin heterogametik secara lebih baik ( st EW = 0,53, N = 16 makalah dan st EW = 0,60, N = 18 makalah, masing-masing) dibandingkan dengan jenis kelamin homogametik ( st EW = 0,00, tidak ada makalah dan st EW = 0,33, dua makalah, masing-masing) (Gambar 3 ).
Sesuai dengan aturan Haldane (Haldane 1922 ; Kulmuni dan Pamilo 2014 ), jenis kelamin dengan kromosom seks heteromorfik menunjukkan penurunan kebugaran hibrida yang lebih besar (Eigenvector = 0,40, PageRank = 0,34, N = 30 makalah) melalui peningkatan sterilitas (secara keseluruhan atau sebagian) atau bahkan ketidakviabilitasannya (lihat Tabel 1 — Glosarium) (Kulmuni dan Pamilo 2014 ). Jumlah kasus yang mematuhi aturan Haldane jauh lebih tinggi daripada yang tidak mematuhinya (
= 20,48; p < 0,01). Penerapan aturan Haldane tampaknya berasal secara tidak langsung dari keberadaan pulau-pulau diferensiasi dalam kromosom seks ( st EW = 0,53, N = 16). Namun, dalam beberapa kasus yang menguatkan aturan Haldane, sterilitas atau ketidakviabilitasan tidak dilaporkan ( st EW = 0,43, N = 13 makalah), atau makalah memberikan bukti bahwa pulau-pulau diferensiasi tidak selalu mengakibatkan sterilitas atau ketidakviabilitasan ( st EW = 0,28, N = 13).
Kemandulan/tidak dapat bertahan hidup yang bias jenis kelamin ( st EW [laki-laki] = 0,86, N = 36 makalah; st EW [perempuan] = 0,89, N = 16 makalah) dan dampak endogen pada sel reproduksi dan/atau karakteristik seksual sekunder ( st EW = 0,54, N = 15 makalah) keduanya dapat mendorong introgresi yang bias jenis kelamin (Walsh et al. 2016 ). Introgresi yang bias jenis kelamin adalah salah satu karakteristik introgresi yang paling penting di antara semua studi yang diterbitkan (Eigenvector = 0,59, PageRank = 1,00, N = 95; Gambar 4 ). Hasil kami menunjukkan bahwa sejumlah besar makalah yang mematuhi aturan Haldane ( st EW = 1,00, N = 30) merupakan hasil mendasar dari keberadaan pulau-pulau diferensiasi pada kromosom terkait jenis kelamin (stEW = 0,59, N = 37). Di sisi lain, ko-kemunculan yang tinggi dengan kasus-kasus introgresi pada kromosom terkait jenis kelamin mengungkapkan bahwa aturan Haldane juga berkontribusi terhadap pengenalan bias jenis kelamin dalam proses introgresi ( st EW = 0,65, N = 11 makalah), yang kemungkinan tidak terjadi melalui promosi langsung kemandulan yang bias jenis kelamin (atau bahkan ketidakberlangsungan total) ( st EW = 0,24, N = 4 makalah).
Hasil kami juga menunjukkan konektivitas tinggi dalam literatur antara introgresi yang bias jenis kelamin dan pengecualian terhadap aturan Haldane ( st EW = 1,00, N = 3 makalah), serta kasus tidak adanya sterilitas/tidak dapat bertahan hidup ( st EW = 0,96, N = 45 makalah). Introgresi yang dimediasi oleh betina (Eigenvector = 0,46, PageRank = 0,64, N = 65 papers) umum terjadi di antara studi kasus introgresi adaptif karena beberapa derajat kemandulan/ketidakmampuan jantan ( st EW = 0,81, N = 34 papers) dalam kasus di mana mereka adalah jenis kelamin heterogametik (terutama pada mamalia [ st EW = 0,22, N = 14 papers] dan serangga [ st EW = 0,21, N = 13 papers]), mengikuti asumsi aturan Haldane ( st EW = 0,53, N = 16 papers). Meskipun introgresi yang dimediasi oleh jantan secara signifikan lebih sedikit dilaporkan (Eigenvector = 0,18, PageRank = 0,30, N = 26 papers) (
= 15,87; p < 0,01), pola ini mengikuti pola yang sama sebagai akibat dari introgresi yang bias jenis kelamin ( st EW = 0,96, N = 25 makalah). Pola ini terkait erat dengan tingkat kemandulan/ketidakmampuan betina yang tinggi ( st EW = 0,67, N = 12 makalah) dalam kasus yang lebih jarang terjadi di mana mereka adalah jenis kelamin heterogametik (terutama pada burung [ st EW = 0,54, N = 14 makalah]).
3.3.3 Introgresi Adaptif pada Spesies di Bawah Pilihan Pasangan Acak atau Seleksi Seksual
Asimetri demografi dapat berdampak signifikan terhadap bagaimana introgresi adaptif berlangsung dalam kompleks spesies di mana pemilihan pasangan antara dua spesies yang saling kawin melibatkan beberapa tingkat keacakan. (Vektor eigen = 0,27, PageRank = 0,71, N = 50 makalah). Dalam kasus tersebut, introgresi adaptif lebih sering dilaporkan diarahkan ke spesies yang jumlahnya lebih sedikit (Vektor eigen = 0,08, PageRank = 0,19, N = 17 makalah) (
= 15,28; p < 0,01). Sejalan dengan pola ini, yang sering dihubungkan dengan demografi atau pilihan pasangan stokastik ( st EW = 0,36, N = 18 makalah), kami menemukan arah asimetris dari introgres adaptif dari spesies asli ke spesies penjajah adalah salah satu pola introgres yang paling banyak dikutip (Eigenvector = 0,28, PageRank = 0,68, N = 70 makalah). Arah asli ke spesies penjajah jauh lebih sering dilaporkan daripada contoh yang lebih jarang dari introgres arah terbalik dari spesies penjajah ke spesies asli (Eigenvector = 0,07, PageRank = 0,22, N = 11 makalah) (Gambar 4 ).
Proses seleksi yang seimbang tampaknya berkaitan erat dengan pola demografi penduduk asli ke penjajah ini ( st EW = 0,36, N = 13 makalah). Arah introgresi ini adalah pola yang tampaknya memengaruhi terutama taksa yang lebih tua (misalnya, st EW [Kerang] = 0,50, N = 3 makalah; st EW [Gimnospermae] = 0,29, N = 4 makalah). Namun, nilai tinggi dari bukti yang dipublikasikan tentang introgresi adaptif dari penduduk asli ke penjajah pada spesies mamalia ( st EW = 0,51, N = 36 makalah) menunjukkan pengaruh fenomena lain selain asimetri demografi dalam kelimpahan spesies. Kami menemukan kemungkinan hubungan antara contoh beberapa derajat pilihan pasangan acak dan arah yang berlawanan dari introgresi adaptif dari spesies penjajah ke asli ( st EW = 0,36, N = 4 makalah) (Gambar 3 ).
Seleksi seksual, didefinisikan sebagai perilaku pengembangbiakan yang mengutamakan pasangan kawin dengan keunggulan kebugaran yang tidak bergantung pada spesiesnya (Eigenvector = 0,19, PageRank = 0,77, N = 37 papers), menunjukkan lebih sedikit kasus yang dilaporkan terkait ko-kejadian dengan asimetri demografis ( st EW = 0,22, N = 8 papers). Bukti seleksi seksual menunjukkan tingkat konektivitas yang tinggi dengan kasus introgresi yang bias jenis kelamin ( st EW = 0,43, N = 16 papers), terutama dengan memengaruhi pilihan pengembangbiakan betina atau jantan, tergantung pada apakah studi kasus introgresi dimediasi oleh betina atau jantan ( st EW = 0,30, N = 11 papers dan st EW = 0,27, N = 7 papers, berturut-turut). Fenomena ini lebih umum terjadi dalam literatur dibandingkan perkawinan asortatif, yang merupakan kekuatan perkembangbiakan yang berlawanan dalam arah spesies yang sama (Eigenvector = 0,06, PageRank = 0,16, N = 13 makalah) (
= 10,58; p < 0,01). Akan tetapi, ada contoh penting terjadinya perkawinan asortatif pada saat yang sama dengan introgresi adaptif (Eigenvector = 0,06, PageRank = 0,13, N = 13 makalah). Hal ini dapat terjadi bahkan dalam kasus di mana seleksi seksual muncul sebagai kekuatan penting yang menentukan arah introgresi adaptif ( st EW = 0,15, N = 2 makalah) (Gambar 3 ).
Kompetisi antarspesies intraseksual (Eigenvector = 0,12, PageRank = 0,77, N = 19 papers) adalah perilaku kawin lain yang dilaporkan berkontribusi pada introgresi terarah, dengan bukti yang dipublikasikan menunjukkan tingkat ko-kemunculan yang tinggi dengan seleksi seksual ( st EW = 0,47, N = 9 papers). Hubungan ini menggambarkan bahwa kedua jenis kelamin dapat memiliki peran mendasar dalam seleksi seksual: satu jenis kelamin dapat memilih pasangan yang paling cocok sementara yang lain bersaing secara aktif melawan individu dari spesies lain untuk mendapatkan peluang reproduksi. Lebih sering, penelitian menggambarkan adanya kompetisi intraseksual dibandingkan dengan ketidakhadirannya (
= 10,23; p < 0,01), yang sangat terkait dengan studi kasus yang menunjukkan introgresi yang bias jenis kelamin ( st EW = 0,53, N = 10 makalah). Literatur yang dianalisis menunjukkan adanya ko-kemunculan antara jenis kompetisi ini dan faktor endogen yang memengaruhi arah introgresi adaptif ( st EW = 0,42, N = 8 makalah) (Gambar 3 ).
3.3.4 Bukti Dampak Lingkungan terhadap Introgresi Adaptif
Beberapa makalah menunjukkan contoh faktor eksogen yang memberikan pengaruh terhadap proses introgresi adaptif (Eigenvector = 0,26, PageRank = 0,68, N = 55 makalah), terutama melalui faktor lingkungan alami ( st EW = 0,74, N = 40 makalah) meskipun ada contoh introgresi adaptif yang sangat relevan dalam menanggapi dampak antropogenik (
= 17,36; p < 0,01). Di sisi lain, mengenai tingkat geografis introgres adaptif, masih ada beberapa contoh empiris alel yang diintrogres di luar zona kontak melalui penjelajahan alel (Eigenvector = 0,11, PageRank = 0,27, N = 26 makalah) (Gambar 4 ). Akhirnya, iterasi historis antara periode divergensi dan introgres adaptif yang berbeda adalah salah satu pola yang paling relevan yang dijelaskan dalam studi kasus (Eigenvector = 0,40, PageRank = 0,57, N = 117 makalah) yang sering dimediasi oleh fluktuasi kondisi lingkungan ( st EW = 0,35, N = 19 makalah) (Gambar 3 ).
4 Diskusi 4.1 Temuan Utama
Studi kami menggarisbawahi peran utama introgres adaptif sebagai kekuatan evolusi utama dalam mensintesis pengetahuan lintas taksa dan dimensi biologis yang beragam (Pfennig 2021 ). Temuan utama menyoroti dominasi introgres autosom nuklir, yang sering kali disertai oleh pulau-pulau diferensiasi pada kromosom terkait jenis kelamin, seperti yang ditunjukkan pada Gambar 3 dan 4. Hasil kami secara khusus mengungkapkan pengaruh yang lebih besar dari seleksi penyeimbangan terhadap pergeseran genetik, khususnya yang memengaruhi introgres nuklir, dengan banyak penelitian mengidentifikasi seleksi penyeimbangan sebagai pendorong utama introgres adaptif. Pola introgres sering kali menunjukkan bias arah dari spesies asli ke spesies yang menjajah, sejalan dengan aturan Haldane, dan biasanya menunjukkan pola bias jenis kelamin yang terkait dengan kemandulan atau ketidakberlangsungan hidup. Meta-analisis mengidentifikasi tiga karakteristik organisme utama yang membentuk introgres adaptif: sel eukariotik, kromosom terkait jenis kelamin, dan seleksi seksual. Kompleksitas sifat introgres adaptif meningkat di seluruh pohon kehidupan, dengan bakteri menunjukkan interaksi yang kurang kompleks dibandingkan dengan mamalia. Sementara introgresi beroperasi pada tingkat genomik, kompleksitasnya mencakup berbagai tingkat biologis, yang memengaruhi fitur sitologi, serta dimensi fisiologis, demografis, dan perilaku. Temuan kami menunjukkan bahwa introgresi adaptif beroperasi dalam konteks evolusi yang lebih luas, di mana divergensi dapat hidup berdampingan dengan aliran gen, yang sering kali dimediasi oleh faktor lingkungan alami dan antropogenik.
4.2 Cakupan dan Evolusi Pengetahuan Tentang Introgresi Adaptif
Studi kami mendukung bukti yang berkembang tentang pergeseran dalam pemahaman introgres adaptif, khususnya sejak revolusi genomik awal tahun 2010-an. Munculnya teknologi sekuensing throughput tinggi, seperti sekuensing genom utuh dan RADseq, memungkinkan peneliti untuk mengidentifikasi daerah genom yang sedang diseleksi secara lebih efektif. Hal ini, dikombinasikan dengan penurunan signifikan dalam biaya sekuensing, memperluas bukti untuk introgresi adaptif di rentang taksonomi dan geografis yang lebih luas, membantu memperjelas karakteristik genomik, dampak fisiologis, dan peran evolusionernya (Hayden 2014 ; Kuhlwilm et al. 2019 ). Sejalan dengan kemajuan ini, temuan kami mengonfirmasi tren utama yang muncul pasca-2012, termasuk predominasi introgresi autosom nuklir, yang sering kali digabungkan dengan pulau-pulau diferensiasi dalam kromosom terkait kelamin. Selain itu, analisis kami menunjukkan prevalensi yang lebih tinggi dari seleksi penyeimbangan atas pergeseran genetik, yang sejalan dengan pengamatan dari beberapa studi pasca-revolusi genomik (Nadachowska-Brzyska et al. 2012 ; While et al. 2015 ). Kami juga menemukan bias arah yang jelas dalam pola introgresi, yang mendukung introgresi dari spesies asli ke spesies penjajah, sebuah tren yang konsisten dengan pekerjaan sebelumnya pada aturan Haldane dan pola introgresi yang bias jenis kelamin, seperti yang dibahas dalam studi seperti Mallet ( 2005 ) dan Feder et al. ( 2012 ). Bias ini khususnya terbukti pada spesies yang menunjukkan kemandulan atau ketidakberlangsungan hidup dalam hibrida, yang sejalan dengan temuan kami tentang introgresi yang bias jenis kelamin dalam beberapa taksa.
Meningkatnya pengakuan atas keuntungan evolusi yang disediakan oleh percampuran genetik telah membentuk kembali persepsi tentang introgresi, bergerak melampaui bias historis yang sering meremehkan signifikansinya (Arnold et al. 2012 ). Hasil kami mencerminkan pergeseran ini, karena kami melihat pengakuan yang berkembang atas peran introgresi dalam memfasilitasi evolusi adaptif. Ini memiliki implikasi yang signifikan untuk diskusi masyarakat dan kebijakan (Baker 2016 ; Fredrickson 2002 ) dan untuk bidang terapan seperti biologi konservasi, khususnya dalam membentuk strategi untuk melestarikan keanekaragaman adaptif (Biermann dan Mansfield 2014 ; Chan et al. 2019 ). Demikian pula, industri pertanian dan peternakan telah mengeksplorasi topik ini untuk memanfaatkan wawasan tentang potensi introgresi untuk meningkatkan sifat adaptif (Henkel et al. 2019 ).
Meta-analisis kami juga menyoroti bahwa studi pasca-2012 telah memberikan bukti yang lebih kuat bahwa spesiasi dapat terjadi bahkan dengan adanya aliran gen. Temuan ini konsisten dengan semakin banyaknya literatur yang menunjukkan bahwa hibridisasi dan introgresi dapat mendorong adaptasi tanpa harus menghalangi spesiasi. Konsep ini merupakan dasar dalam studi awal (Mayr 1942 ) dan telah dikembangkan lebih lanjut dalam beberapa tahun terakhir (Harrison dan Larson 2014 ; Mallet 2005 ), yang menggambarkan bahwa kedua proses tersebut tidak saling eksklusif, bahkan ketika kekuatan evolusionernya bekerja dalam arah yang berlawanan, yaitu konvergensi dan divergensi.
Kemajuan metodologis dan teknologis dalam dua dekade terakhir telah menjadi pendorong utama perluasan signifikan dalam penelitian introgresi adaptif. Studi kami menyoroti pengaruhnya saat ini dalam pengakuan yang semakin besar terhadap introgresi adaptif sebagai kekuatan evolusi yang penting, memperluas cakupannya dengan mengintegrasikan dimensi genomik, fisiologis, ekologis, dan perilaku ke dalam pemahaman yang lebih komprehensif tentang adaptasi. Dengan menganalisis berbagai spesies, dari prokariota hingga mamalia, kami menunjukkan bahwa introgresi adaptif tidak terbatas pada beberapa organisme model tetapi merupakan fenomena yang tersebar luas dengan implikasi yang mendalam bagi biologi evolusi.
4.3 Kompleksitas Struktur Organisme yang Mempengaruhi Karakteristik Introgresi Adaptif
Hasil kami menunjukkan bahwa pada spesies prokariota, introgres adaptif beroperasi terutama melalui rekombinasi genetik melalui mobilisasi konjugatif kromosom, sejalan dengan penelitian sebelumnya (McAshan et al. 1999 ) dan menggarisbawahi pentingnya dalam memberikan keuntungan adaptif, seperti resistensi antibiotik (lihat McAshan et al. 1999 ; Palmer et al. 2010 ). Namun, plasmid menyusun sebagian besar genom tambahan, yang bertanggung jawab atas banyak transfer gen alami mereka (Palmer et al. 2010 ). Prevalensi seleksi penyeimbangan yang diamati dalam meta-analisis kami menunjukkan bahwa proses seleksi, bukan pergeseran genetik, memainkan peran kunci dalam fiksasi alel yang diintrogres, terutama pada spesies prokariotik, yang menangkal proses acak yang dapat menyebabkan alel langka menghilang dalam beberapa generasi. Namun, kejadian yang lebih tinggi dari kasus seleksi keseimbangan yang diamati dalam literatur (Nadachowska-Brzyska et al. 2012 ; While et al. 2015 ) mungkin disebabkan oleh penekanan pada gen yang berpotensi memberikan keuntungan adaptif, sehingga mengurangi dampak pergeseran genetik. Sebaliknya, peristiwa introgresi yang hilang karena pergeseran genetik sering kali sulit untuk diukur hanya karena peristiwa tersebut telah menghilang.
Pada spesies eukariotik, temuan kami menunjukkan bahwa introgresi adaptif sangat dipengaruhi oleh ketidaksesuaian sitonuklir, perbedaan dalam laju rekombinasi, dan seleksi dalam sel. Hal ini mendukung kesimpulan dari meta-analisis kami bahwa introgresi terutama terjadi di dalam nukleus, terutama pada autosom, kemungkinan di daerah yang tidak sepenuhnya terkait dengan cDNA atau tidak resistan terhadap rekombinasi, dan sering didorong oleh tekanan seleksi pada tingkat sel (Burton dan Barreto 2012 ; Derr et al. 2012 ; Matosiuk et al. 2014 ).
Pada spesies eukariotik multiseluler dengan kromosom seks heteromorfik, sementara penelitian kami menunjukkan bahwa introgresi adaptif terjadi terutama pada autosom, pulau-pulau diferensiasi pada dasarnya hadir dalam kromosom terkait seks. Kromosom terkait seks terlibat dalam fungsi-fungsi spesifik dalam reproduksi yang memberikan tekanan, misalnya, pada fisiologi perkembangbiakan. Dengan demikian, introgresi dalam kromosom seks terkadang dapat memiliki konsekuensi negatif seperti kemandulan yang bias jenis kelamin atau bahkan tidak dapat bertahan hidup (Storchová et al. 2004 ). Namun, prevalensi tinggi penelitian yang mengidentifikasi introgresi yang bias jenis kelamin menunjukkan organisme dapat melewati kendala fisiologis ini (yaitu, kemandulan yang bias jenis kelamin atau bahkan tidak dapat bertahan hidup).
Literatur menunjukkan bahwa perbedaan demografi, seperti perbedaan dalam kelimpahan spesies, mendorong perkawinan ke arah spesies yang paling melimpah, khususnya dalam kasus pilihan pasangan acak. Telah diperdebatkan bahwa dalam kasus ekstrem bahkan dapat menyebabkan hilangnya spesies paling langka melalui genangan genetik (Todesco et al. 2016 ). Namun, ketika introgresi adaptif bersifat asimetris dan bias arah terhadap spesies yang kurang melimpah (pola arah tertentu yang muncul dari meta-analisis kami), hal itu dapat memungkinkan spesies ini untuk menggabungkan sifat-sifat menguntungkan dari spesies yang lebih melimpah. Ini adalah proses evolusi potensial yang memungkinkan spesies yang lebih langka untuk beradaptasi dengan lingkungan, mengatasi kendala dari depresi perkawinan sedarah dan genangan genetik.
Pada spesies eukariota yang berevolusi baru-baru ini dan kompleks, perilaku seksual, seperti perkawinan non-acak, dapat mengarah pada perkawinan asortatif, di mana individu cenderung mencari pasangan kawin sejenis, sehingga mengurangi terjadinya introgresi (Hume et al. 2018 ). Namun, hasil kami menunjukkan bahwa ketika perkawinan tidak acak, ada kecenderungan untuk mengutamakan individu yang paling bugar melalui seleksi seksual, terlepas dari spesiesnya (Parrett dan Knell 2018 ). Perilaku ini dapat memungkinkan spesies yang kurang bugar untuk beradaptasi dan berkembang dengan menggabungkan sifat-sifat yang memberikan kebugaran lebih tinggi pada spesies saudaranya. Ini dapat membantu menjelaskan bagaimana spesies ini dapat melewati batasan yang dihasilkan dari perkawinan asortatif.
4.4 Mekanisme Evolusi yang Diketahui dan Karakteristik Introgresi Adaptif
Seiring dengan evolusi organisme dan peningkatan kompleksitas struktural, tantangan baru terhadap introgresi adaptif muncul dari hambatan aliran gen, termasuk perubahan genetik (misalnya, inversi, fusi), inovasi dalam sistem seluler dan fisiologis, dan pergeseran dalam strategi perkawinan. Meskipun demikian, introgresi adaptif masih dapat terjadi. Kekuatan evolusi yang dihasilkan dari divergensi, yang mendorong adaptasi berbeda dalam populasi yang berbeda, tidak serta merta menghalangi introgresi. Sebaliknya, proses seperti diversifikasi populasi dan pertukaran gen sering kali hidup berdampingan dan berinteraksi, mendorong adaptasi dengan cara yang dinamis dan saling berhubungan. Interaksi dinamis ini dieksplorasi lebih lanjut di bagian berikut, yang meneliti mekanisme yang memungkinkan introgresi adaptif tetap ada meskipun ada kekuatan evolusi yang berlawanan.
4.4.1 Menyeimbangkan Seleksi Versus Penyimpangan Genetik
Kemungkinan gen yang diintrogres akan terbentuk dalam genom taksa reseptor masih diperdebatkan. Karena gen ini awalnya langka, gen tersebut cenderung sangat sensitif terhadap proses stokastik seperti pergeseran genetik. Efek ini khususnya terlihat jelas pada populasi yang seleksi seksualnya lemah atau tidak ada, dan ketika ukuran populasi efektifnya rendah (Gompert et al. 2013 ).
Namun, sementara pergeseran genetik dapat menyebabkan hilangnya alel yang terintrogres, seleksi penyeimbang mendorong introgres adaptif dengan mempertahankan beberapa alel pada frekuensi yang lebih tinggi, yang menangkal proses seperti seleksi untuk pasangan pengembangbiakan sejenis (perkawinan asortatif) atau pembuahan sendiri (King et al. 2006 ). Temuan kami menekankan bahwa seleksi penyeimbang sering kali mendukung introgres adaptif, mengurangi hilangnya alel melalui pergeseran dan mendukung pembentukan alel adaptif. Kejadian seleksi penyeimbang yang lebih tinggi menunjukkan bahwa alel yang terintrogres lebih mungkin dipertahankan dalam populasi, sementara pergeseran genetik dapat membatasi persistensi mereka dalam populasi yang lebih kecil. Dengan demikian, seleksi penyeimbang tidak hanya mengurangi hilangnya alel karena pergeseran, tetapi secara aktif berkontribusi pada pembentukan introgres adaptif (While et al. 2015 ). Namun, tidak semua mekanisme seleksi penyeimbang cenderung mendorong introgres adaptif dengan cara yang sama (lihat While et al. ( 2015 ) untuk pembahasan lebih lanjut). Misalnya, gen yang mengalami seleksi bergantung frekuensi negatif, yang memiliki tingkat migrasi lebih tinggi, dapat memungkinkan introgresi dan pembentukan yang lebih efisien dalam kumpulan gen spesies reseptor (Brennan et al. 2013 ). Mekanisme ini mendukung temuan kami bahwa introgresi adaptif lebih mungkin terjadi ketika seleksi penyeimbangan bekerja bersamaan dengan aliran gen.
4.4.2 Introgresi Autosomal Versus Diskordansi Sitonuklear
Variasi struktural seperti inversi kromosom, translokasi, dan fusi dapat berkontribusi pada ketidakcocokan genetik, termasuk ketidaksesuaian sitonuklir. Perubahan struktural ini dapat mengganggu rekombinasi dan menciptakan hambatan pada aliran gen, yang mendorong isolasi dan divergensi reproduksi. Secara khusus, inversi kromosom dapat menekan rekombinasi di zona hibrida, mempertahankan kombinasi alel yang tidak kompatibel atau ketidakcocokan sitonuklir, yang dapat menyebabkan kebugaran yang berkurang pada hibrida. Tinjauan kami mendukung asumsi ini dengan mengidentifikasi serangkaian studi yang konsisten yang menyoroti fenomena seperti inversi kromosom yang sering kali mengandung kompleks gen yang beradaptasi bersama, yang mencegah rekombinasi dengan alel yang tidak kompatibel dari populasi lain (lihat Hoffmann dan Rieseberg 2008 ; Feder et al. 2012 ). Pelestarian ini dapat menstabilkan ketidakcocokan genetik yang dapat melibatkan interaksi antara gen nuklir dan mitokondria, terutama dengan adanya hubungan epistatik atau underdominance (Feder et al. 2012 ). Selain itu, penataan ulang kromosom dapat meningkatkan kemungkinan ketidakcocokan tersebut dengan mengubah ekspresi gen atau mengganggu jaringan regulasi yang penting untuk interaksi sitonuklir (Navarro dan Barton 2003 ).
Ketidaksesuaian sitonuklir dapat memengaruhi kebugaran spesies, yang menyebabkan berkurangnya kesuburan dan/atau penyakit, atau bahkan mendorong pembentukan hibrida yang tidak dapat hidup (Burton dan Barreto 2012 ; Derr et al. 2012 ; Matosiuk et al. 2014 ). Meskipun ada beberapa kasus ketidakcocokan genetik yang dilaporkan, termasuk ketidaksesuaian sitonuklir (lihat Gagnaire et al. 2012 ), hasil kami menekankan terjadinya introgresi pada cDNA dan nDNA dalam ko-introgresi, yang menunjukkan bahwa individu dengan genom nuklir satu spesies (secara penuh atau sebagian) dan sitogenom spesies lain (secara penuh atau sebagian) seringkali dapat hidup dan mampu melakukan backcross (Pujolar et al. 2014 ; Sarver et al. 2017 ).
Lokasi introgresi dalam genom eukariota dikaitkan dengan ketidaksesuaian sitonuklir, sebuah fenomena yang lebih umum diamati dalam studi hewan daripada dalam literatur tumbuhan (Morales et al. 2018 ). Mayoritas kasus introgresi adaptif yang dilaporkan dalam tinjauan kami terjadi pada tingkat nuklir (Hulsey et al. 2013 ), meskipun beberapa penulis menganggap bahwa genom organel mengalami introgresi lebih mudah dan lebih sering daripada genom nuklir (Giannoulis et al. 2019 ; Hawkins et al. 2016 ; Wade and Goodnight 2006 ). Namun, temuan kami mendukung bahwa, sementara introgresi nuklir bertanggung jawab atas ketidaksesuaian sitonuklir hanya dalam rentang kondisi yang sempit (Bonnet et al. 2017 ), ketidaksesuaian yang kuat sering didorong oleh seleksi genom sitoplasma, khususnya dalam sistem biologis dengan kapasitas penyebaran terbatas (Brower 2018 ).
Temuan kami mendukung gagasan bahwa introgresi cDNA dapat terjadi tanpa mendorong terjadinya discordance, terutama jika introgresi tersebut tidak terhubung dengan daerah nDNA yang bertanggung jawab atas inkompatibilitas antarspesies (Di Candia dan Routman 2007 ). Pewarisan cDNA oleh satu induk juga memperkuat pentingnya penyebaran yang bias jenis kelamin (Kulikova et al. 2004 ) untuk introgresi adaptifnya, yang memengaruhi interaksi gen sitonuklir (Llopart et al. 2014 ). Meskipun demikian, setelah periode isolasi, interaksi sitonuklir dapat berkembang secara terpisah (Herrig et al. 2014 ). Oleh karena itu, ketidakseimbangan antara cDNA dan haplotipe autosomal (Oliver et al. 2002 ) dapat meminimalkan kemungkinan introgresi cDNA (Herrig et al. 2014 ), menjadikan introgresi nDNA tanpa aliran gen cDNA sebagai fenomena umum di antara studi kasus introgresi adaptif, yang dapat menjelaskan pola yang muncul dari hasil kami.
4.4.3 Introgresi Autosomal Versus Pulau Diferensiasi pada Kromosom Terkait Jenis Kelamin
Introgresi asimetris nuklir, yang mendukung autosom (lihat Carling dan Brumfield 2008 ), berarti kromosom seks menjadi kandidat utama untuk membentuk penghalang aliran gen dan menyimpan pulau-pulau diferensiasi (Runemark et al. 2018 ). Hasil kami mengonfirmasi pola ini, yang menunjukkan bahwa introgresi sebagian besar terjadi pada autosom, sementara pulau-pulau diferensiasi lebih umum ditemukan pada kromosom tertaut seks. Dengan demikian, kehadiran pulau-pulau diferensiasi secara bersamaan dalam kromosom seks di samping introgresi autosom adaptif diidentifikasi sebagai pola yang lazim dalam literatur. Hal ini sering kali mewakili keseimbangan antara seleksi divergen dan kekuatan homogenisasi genetik dalam nukleus (Minder dan Widmer 2008 ). Namun, pulau-pulau diferensiasi—didefinisikan sebagai daerah genomik yang menunjukkan tingkat diferensiasi yang luar biasa tinggi antara populasi atau spesies (Nachman dan Payseur 2012 )—dapat dihasilkan oleh proses yang tidak terkait dengan spesiasi (Nosil et al. 2012 ), seperti dalam kasus berkurangnya keragaman karena seleksi terkait (Cruickshank dan Hahn 2014 ). Perbedaan dalam tingkat rekombinasi pada kromosom seks dapat membantu menjelaskan sentralitas tinggi pulau-pulau diferensiasi dalam kromosom terkait seks dalam meta-analisis kami (Wadsworth et al. 2015 ). Kromosom seks memiliki karakteristik, seperti diferensiasi morfologis dan genetik antara kromosom seksual X dan Y, atau Z dan W, atau pembatasan pada rekombinasi (Storchová et al. 2004 ). Karakteristik ini bersama dengan peran mendasar yang dimainkannya dalam reproduksi seksual (misalnya, pembentukan gamet) membuatnya sangat tahan terhadap introgresi pada spesies dengan reproduksi seksual dioecious. Analisis kami menunjukkan bahwa kromosom yang tertaut dengan jenis kelamin sering kali resistan terhadap introgresi karena kendala-kendala ini, namun ketika introgresi terjadi, hal itu sering kali melibatkan daerah-daerah yang memiliki relevansi fungsional terhadap reproduksi. Keterbatasan dalam variasi perbaikan DNA dan gen dengan fungsi mitokondria (Runemark et al. 2018 ) dan aspek demografis yang berasal dari pewarisan kromosom seks oleh orang tua tunggal (dalam jenis kelamin heterogametik), (misalnya, penyebaran yang bias jenis kelamin) (lihat Kulikova et al. 2004 ), dapat menjelaskan kelangkaan atau bahkan tidak adanya introgresi dalam kromosom seks (Cahill et al. 2015 ), proses serupa yang terjadi pada spesies haplodiploid seperti semut dan lebah (lihat Ghenu et al. 2018)). Temuan ini konsisten dengan hasil kami, di mana introgresi pada kromosom seks diamati lebih jarang daripada pada autosom tetapi menunjukkan dampak fungsional yang jelas ketika terjadi. Hal ini dapat dijelaskan oleh paparan unik kromosom seks, khususnya pada jenis kelamin heterogametik, terhadap tekanan seleksi, yang dapat meningkatkan kapasitas evolusi adaptifnya meskipun terjadi kehilangan gen pada kromosom yang tidak melakukan rekombinasi (misalnya, kromosom Y) (Muirhead dan Presgraves 2016 ).
Kehadiran pulau-pulau diferensiasi pada kromosom terkait-kelamin tidak selalu mengarah pada divergensi, seperti yang terlihat pada daerah genomik dengan keragaman yang berkurang, membantu menjelaskan mengapa mereka tidak selalu mengakibatkan kemandulan atau ketidakberlangsungan (Cruickshank dan Hahn 2014 ). Meta-analisis kami mendukung hal ini, menunjukkan bahwa pulau-pulau diferensiasi dalam kromosom seks sering hidup berdampingan dengan aliran gen dalam autosom, menjaga keseimbangan antara seleksi dan kekuatan konvergen. Pengecualian dalam lokasi genomik pulau-pulau diferensiasi, baik pada autosom atau cDNA (Sun et al. 2012 ), tidak mengandung gen yang memengaruhi isolasi reproduktif tetapi masih mengalami diferensiasi yang signifikan, sering kali sebagai hasil dari proses acak, seperti pergeseran genetik (Nosil et al. 2012 ).
4.4.4 Introgresi yang Berpihak pada Jenis Kelamin Dibandingkan dengan Kendala Fisiologis
Pulau-pulau diferensiasi dalam kromosom terkait-jenis kelamin biasanya hanya berukuran beberapa ratus kilobasa, sedangkan daerah yang mengalami introgresi dapat meluas hingga beberapa megabasa (Gagnaire et al. 2013 ). Asimetri ini muncul dari evolusi dan dorongan meiosis kromosom seks yang lebih cepat (Sun et al. 2012 ). Pulau-pulau diferensiasi dalam kromosom terkait-jenis kelamin dapat bertindak seperti penghalang pengembangbiakan, yang terkait dengan isolasi temporal dan perilaku (Wadsworth et al. 2015 ), yaitu memengaruhi kemandulan/ketidakmampuan hidup individu yang bias jenis kelamin (lihat Dopman et al. 2005 ). Baik pulau-pulau diferensiasi dalam kromosom terkait-jenis kelamin maupun kemandulan/ketidakmampuan hidup menunjukkan keterhubungan yang tinggi menurut hasil kami dan lebih disukai memengaruhi jenis kelamin heterogametik. Ini selaras dengan aturan Haldane, seperti yang disorot dalam hasil kami, di mana individu heterogametik mengalami pengurangan kebugaran yang lebih besar (lihat Haldane 1922 ; Kulmuni dan Pamilo 2014 ), yang sering kali memediasi pola introgresi asimetris. Literatur yang ditinjau menguatkan bahwa hasil fisiologis yang berasal dari aturan Haldane sering kali bertanggung jawab untuk mendorong pola asimetris introgresi yang bias jenis kelamin (Sackton et al. 2011 ). Pola-pola ini umumnya dimediasi oleh perempuan, terutama dalam kasus laki-laki homogametik dengan beberapa tingkat kerusakan pascazigot. Analisis kami mengidentifikasi contoh-contoh introgresi yang bias jenis kelamin di mana aturan Haldane ditegakkan, dengan alel-alel yang diintrogresi lebih sering dipertahankan pada laki-laki homogametik karena tekanan selektif yang mendukung kelangsungan hidup reproduksi. Contoh kejadian simultan antara introgres yang bias jenis kelamin dan pengecualian terhadap aturan Haldane juga dijelaskan dalam literatur, mungkin terkait dengan ciri demografi dan/atau perilaku, seperti heterosis selektif positif pada satu jenis kelamin (introgres uniparental) (Beresford et al. 2017 ). Selain itu, jenis kelamin dengan tingkat penyebaran yang lebih tinggi biasanya menunjukkan tingkat aliran gen yang lebih tinggi dengan populasi konspesifik di luar zona kontak (Kulikova et al. 2004 ; Morales et al. 2018 ). Di sisi lain, meskipun jarang diidentifikasi dalam tinjauan literatur kami, pulau diferensiasi dan/atau introgres dalam kromosom terkait jenis kelamin juga dapat mengekspresikan dampak endogen lainnya pada sel reproduksi dan pada karakteristik seksual sekunder, yang juga dapat menyebabkan penurunan kebugaran hibrida dan persilangan balik (Walsh et al. 2016 ).
4.4.5 Introgresi Adaptif Terarah Versus Pembendungan Genetik
Proses stokastik, seperti kolonisasi, kepunahan, dan kemacetan, dapat memengaruhi kelangsungan hidup spesies (Vaillant et al. 2013 ). Literatur yang diulas mendukung fakta ini, menunjukkan bahwa introgres adaptif dapat terjadi dalam konteks di mana peristiwa stokastik membentuk kembali dinamika populasi, seperti selama kolonisasi atau kemacetan (Lexer et al. 2004 ). Ketika spesies berbeda dalam kelimpahan, perkawinan cenderung menguntungkan spesies yang lebih melimpah (Lexer et al. 2010 ). Hal ini biasanya menghasilkan aliran gen dari spesies dominan ke spesies yang lebih langka, yang berpotensi menyebabkan kepunahannya melalui genangan genetik (Lexer et al. 2010 ). Namun, dikombinasikan dengan stokastisitas pembiakan, proses ini sering kali mendorong introgres adaptif arah asimetris, suatu pola yang sering diamati dalam penelitian kami yang diambil (lihat Takahashi et al. 2017 ). Meta-analisis kami mengungkap prevalensi kuat dari introgresi terarah, khususnya dalam skenario di mana spesies asli memperkenalkan alel adaptif ke dalam penjajah, konsisten dengan peristiwa perluasan spesies (lihat Currat et al. 2008 ; Excoffier et al. 2008 ). Alel adaptif yang diperkenalkan ke dalam spesies penjajah dari spesies asli yang lebih melimpah dan lebih beradaptasi dengan kondisi lokal dapat membantu spesies penjajah terbentuk di lingkungan baru (lihat Garcia-Elfring et al. 2017 ). Hasil kami mendukung hal ini, menunjukkan bahwa aliran gen asli ke penjajah memfasilitasi adaptasi ke lingkungan baru, membantu pembentukan populasi baru. Tinjauan kami juga mengidentifikasi kasus-kasus di mana pilihan pasangan mendorong introgresi adaptif dari penjajah ke penduduk asli, khususnya ketika stokastisitas bukan pendorong utama. Introgresi penjajah ke penduduk asli kurang umum tetapi tetap signifikan dalam temuan kami, sering dikaitkan dengan kasus-kasus di mana tekanan lingkungan mendukung sifat-sifat yang diperkenalkan oleh penjajah. Hasil meta-analisis menunjukkan bahwa seleksi keseimbangan terkait erat dengan asimetri demografi (penduduk asli ke penjajah atau penjajah ke penduduk asli), yang kemungkinan memungkinkan alel yang diintrogres terbentuk di kumpulan gen spesies reseptor (Fijarczyk et al. 2018 ), menangkal pergeseran genetik, penumpukan genetik, dan memungkinkan persistensi sifat adaptif, terlepas dari ketidakseimbangan kelimpahan. Temuan ini menunjukkan bahwa asimetri demografi, bersama dengan faktor ekologi, memainkan peran penting dalam membentuk pola introgresi, terutama dalam lingkungan dinamis tempat interaksi spesies secara bersamaan dipengaruhi oleh kelimpahan dan kebutuhan adaptif.
4.4.6 Seleksi Seksual Versus Perkawinan Asortatif
Proses pemilihan pasangan memainkan peran kunci dalam membentuk pola divergensi dan introgres adaptif. Perkawinan asortatif berdampak positif pada evolusi divergen dengan mempromosikan pilihan pasangan terhadap spesies yang sama, yang menangkal introgres adaptif. Namun, meta-analisis kami menemukan introgres adaptif terjadi bersamaan dengan perkawinan asortatif (Lexer et al. 2010 ; Mavárez et al. 2006 ; Ng et al. 2017 ). Temuan-temuan ini menunjukkan bahwa introgresi dalam kasus-kasus seperti itu sering kali melibatkan alel-alel yang terkait dengan pengenalan pasangan, yang menyoroti peran kompleks perkawinan asortatif dalam introgres adaptif. Alel-alel ini dapat memediasi introgresi gen-gen tetangga yang terkait dengan sifat-sifat yang secara tidak langsung ditargetkan oleh introgresi adaptif (Mavárez et al. 2006 ).
Tinjauan kami menemukan bahwa seleksi seksual lebih menyukai pasangan dengan kebugaran yang lebih tinggi, terlepas dari spesiesnya, yang berpotensi memengaruhi arah dan tingkat introgres adaptif, sebuah pola utama dalam meta-analisis kami (Alex Sotola et al. 2019 ; Salazar et al. 2010 ). Temuan kami mengindikasikan bahwa seleksi seksual memainkan peran penting dalam membentuk introgres adaptif, khususnya ketika menyukai sifat-sifat kebugaran yang tinggi, yang memfasilitasi aliran gen melintasi batas-batas spesies. Pergaulan bebas seksual kemungkinan berkontribusi pada proses ini (Baldassarre et al. 2014 ), menyebarkan sifat-sifat baru melalui sapuan selektif (lihat Bay dan Ruegg 2017 ). Dalam skenario ekspansi, seleksi seksual cenderung menyukai spesies asli, yang beradaptasi lebih baik dengan kondisi lokal (lihat Yang et al. 2018 ). Proses ini dapat memberikan keuntungan adaptif bagi spesies penjajah dengan memfasilitasi penggabungan gen-gen adaptif yang diintrogres dari spesies asli (lihat Zielinski et al. 2013 ). Pola ini selaras dengan meta-analisis kami, yang menunjukkan bahwa alel asli yang diintrogress sering meningkatkan kemampuan adaptasi penjajah terhadap lingkungan baru. Namun, perubahan lingkungan di zona kontak dapat mendukung karakteristik spesies penjajah, yang membuka jalan bagi ekspansi. Dari perspektif teoritis, karakteristik tersebut dapat menjadi target seleksi seksual oleh spesies asli, yang memungkinkan mereka beradaptasi dengan perubahan lingkungan dan mengarahkan kembali introgres adaptif, membantu membenarkan bukti introgres adaptif yang terjadi di kedua arah dalam literatur yang ditinjau, pada spesies di bawah seleksi seksual. Temuan kami mendukung hipotesis bahwa seleksi seksual dapat secara signifikan memengaruhi introgres adaptif, terutama pada spesies dengan preferensi perkawinan yang rumit. Prevalensi seleksi seksual yang lebih besar pada spesies yang lebih kompleks menggarisbawahi pentingnya faktor perilaku dalam memfasilitasi aliran gen lintas spesies.
Perilaku lain di bawah proses selektif, seperti pengasuhan orang tua yang berbeda dan sosialitas, mungkin juga mendukung introgresi arah asimetris (Sullivan et al. 2016 ; Trigo et al. 2014 ). Akhirnya, kompetisi antarspesies intra-seksual, sebuah simpul penting dalam jaringan kami, dapat memediasi introgresi adaptif, mempromosikan pewarisan gen dari spesies dengan kapasitas kompetitif yang lebih besar (ukuran lebih besar, gigitan lebih kuat, pembungaan lebih awal, dll.) ke spesies lain, sehingga selanjutnya berkontribusi pada introgresi arah asimetris. Kompetisi antarspesies intra-seksual dianggap hanya terjadi di bawah seleksi seksual yang lemah atau tidak ada (While et al. 2015 ). Namun, hasil kami menunjukkan bahwa kompetisi intra-seksual sering melengkapi seleksi seksual, dengan keduanya bertindak bersama untuk membentuk keseimbangan antara introgresi adaptif dan kekuatan divergensi, khususnya dalam sistem perkawinan kompetitif.
4.4.7 Dampak Lingkungan terhadap Introgresi Adaptif
Introgresi adaptif dapat mendorong adaptasi terhadap kondisi lingkungan dengan meningkatkan variasi genetik, yang pada gilirannya memengaruhi kemampuan berevolusi. Seperti yang ditunjukkan oleh Grant dan Grant ( 2016 ), introgresi dapat mendorong adaptasi dengan memperkenalkan sifat-sifat yang menguntungkan sekaligus menjadi hasil dari tekanan selektif yang membentuk konvergensi morfologi dan genetik dalam kondisi alami. Hasil kami selanjutnya mendukung peran ganda introgresi ini dalam proses evolusi. Kondisi lingkungan memediasi iterasi antara periode divergensi dan konvergensi (Pauquet et al. 2018 ; Pfennig 2007 ; Staubach et al. 2012 ). Berdasarkan telaah pustaka kami, faktor lingkungan alamiah dan dampak antropogenik memengaruhi introgresi adaptif melalui, misalnya, respons adaptif terhadap antibiotik (lihat McAshan et al. 1999 ), pestisida (lihat Song et al. 2011 ), penggundulan hutan (lihat Alexander et al. 2017 ), atau perubahan iklim (lihat De La Torre et al. 2014 ; Quigley et al. 2019 ). Pada gilirannya, perubahan ketinggian (lihat Huerta-Sánchez et al. 2014 ), penyerbuk (lihat Wang et al. 2013 ), paparan patogen (lihat Jagoda et al. 2018 ), cahaya ultraviolet (lihat Ding et al. 2013 ), dan suhu (lihat Suarez-Gonzalez et al. 2016 ) dapat memodulasi kekuatan hambatan pasca-perkembangbiakan ekologis (lihat Reidenbach et al. 2012 ) dan mendorong variasi dalam kebugaran hibrida dan persilangan balik melalui introgres adaptif (Kingston et al. 2017 ). Terlepas dari tingkat dan konteks historisnya, introgres adaptif yang dimediasi oleh kondisi lingkungan membantu spesies untuk memperluas jangkauannya ke wilayah ekologi baru (Aleksic et al. 2018 ) dan/atau dapat memungkinkan mereka untuk beradaptasi dengan kondisi lingkungan baru di luar relung aslinya (De La Torre et al. 2014 ; Quigley et al. 2019 ). Perbedaan dalam kondisi lingkungan lokal selama evolusi dua garis keturunan yang berbeda dapat membantu menjelaskan perbedaan spasial dalam karakteristik introgresi adaptif antara zona kontak yang berbeda (Stuckas et al. 2009 ). Akhirnya, allele surfing memungkinkan spesies untuk menyebarkan alel yang diintrogresi ke seluruh wilayah spesies (Sarver et al. 2017 ), meningkatkan jangkauan geografis introgresi adaptif di luar batas zona kontak. Dalam tinjauan kami, kami menemukan contoh fenomena ini, misalnya, pada serangga yang dapat menyebarkan alel yang diintrogresi lebih dari 400 km (lihat Norris et al. 2015) .) dan mamalia yang berjarak lebih dari 2000 km dari zona kontak hanya dalam beberapa generasi (lihat Matosiuk et al. 2014 ). Hasil ini menunjukkan bahwa tekanan lingkungan berperan sebagai pendorong signifikan introgresi adaptif. Temuan kami menyoroti peran faktor ekologi dalam memodulasi aliran gen, mendukung hipotesis bahwa alel yang diintrogresi dapat memberikan keuntungan adaptif dalam lingkungan yang berubah.
4.5 Keterbatasan
Mengingat sifat penelitian ini, asumsi bahwa data dalam kumpulan data kami mewakili alel atau sifat yang diintegrasikan secara adaptif didasarkan pada interpretasi penulis makalah yang diambil dari tinjauan sistematis, yang mungkin memiliki batasan tertentu.
Pertama, penyeimbangan seleksi dan introgres adaptif dapat meninggalkan jejak genomik yang serupa, seperti polimorfisme frekuensi menengah berlebih yang mengapit lokus yang diseleksi. Tumpang tindih ini mempersulit diferensiasi antara proses-proses ini dalam banyak penelitian. Sementara beberapa penelitian asli mengandalkan pola sapuan atau sinyal genomik lainnya untuk menyimpulkan introgres adaptif, yang lain mungkin tidak memperhitungkan penyeimbangan seleksi sebagai faktor pengganggu. Dengan demikian, kesimpulan dari meta-analisis kami mungkin, dalam beberapa kasus, mencakup contoh-contoh penyeimbangan seleksi daripada introgres adaptif, khususnya di wilayah genomik tempat kedua proses tersebut terjadi bersamaan. Salah tafsir lainnya dapat terjadi jika peneliti mengabaikan penjelasan alternatif untuk pembagian alel antara garis keturunan yang berbeda, seperti penyortiran garis keturunan yang tidak lengkap (terutama dalam makalah yang lebih lama). Di sisi lain, ketika menafsirkan pola introgres adaptif, penting untuk memperhitungkan peran mutasi yang merusak, khususnya dalam konteks pencampuran dari populasi besar ke populasi kecil, karena hal itu dapat mengacaukan identifikasi wilayah yang diseleksi (Kim et al. 2018 ). Sementara beberapa metode masih mengabaikan varian yang merusak, lebih dari 75% makalah yang disertakan dalam studi kami diterbitkan setelah tahun 2012, periode yang ditandai oleh kemajuan metodologis yang signifikan, termasuk perangkat genomik dan statistik, yang meningkatkan akurasi dalam membedakan introgresi adaptif dari proses genetik lainnya. Selain itu, tinjauan sistematis kami dengan cermat menyaring studi untuk ketelitian metodologis dan kekuatan bukti, yang bertujuan untuk meminimalkan penyertaan studi di mana mekanisme alternatif untuk introgresi tidak dapat dikesampingkan secara wajar atau di mana varian yang merusak tampaknya diabaikan. Ukuran sampel studi yang besar yang dianalisis semakin memperkuat kekokohan dan keandalan pola yang diamati dalam introgresi adaptif.
Selain itu, beberapa salah tafsir mungkin mencakup contoh-contoh di mana alel yang diintrogresikan secara keliru diasumsikan bersifat adaptif tanpa bukti yang memadai. Salah tafsir ini berpotensi memengaruhi kumpulan data kami dengan memperkenalkan bias atau ketidakakuratan dalam kesimpulan yang diambil mengenai mekanisme dan peran introgresi adaptif dalam proses evolusi. Namun, meskipun mungkin ada contoh-contoh di mana proses introgresi dipublikasikan sebagai adaptif tetapi kemudian ditetapkan netral, prevalensi tinggi penelitian yang menunjukkan bukti mediasi eksogen dari proses introgresi menunjukkan bahwa hal itu terutama terjadi di wilayah genom di bawah seleksi positif. Meskipun demikian, kami menekankan pentingnya bagi para peneliti untuk mengevaluasi secara kritis bukti yang mendukung hasil adaptif dari introgresi dan mempertimbangkan penjelasan alternatif untuk memastikan kekokohan temuan mereka.
4.6 Kesenjangan Pengetahuan dan Tantangan Masa Depan
Munculnya pendekatan sekuensing berthroughput tinggi menyebabkan proliferasi contoh empiris tentang pembagian alel antara taksa yang berbeda, terutama setelah 2012, selain memungkinkan identifikasi proses yang mengarah pada pembentukannya (misalnya, introgresi atau penyortiran garis keturunan yang tidak lengkap). Penggabungan pengetahuan ini tentang karakteristik umum introgresi telah memungkinkan kita untuk menafsirkan pola dan karakteristik proses yang mengarah pada adaptasi secara lebih komprehensif. Ini telah memberikan gambaran umum tentang hipotesis utama yang akan diuji mengenai peran evolusi introgresi, yang tetap menjadi tantangan penting. Misalnya, pengenalan varian genetik melalui introgresi adaptif dapat menjadi sangat penting dalam proses evolusi spesies yang menghadapi depresi perkawinan sedarah, yang memerlukan penyelidikan lebih dalam dalam literatur saat ini.
Studi kami mengidentifikasi beberapa mekanisme evolusi yang terkait dengan introgresi adaptif yang terjadi bersamaan dengan kekuatan divergensi. Hal ini menggarisbawahi bahwa proses-proses ini tidak saling eksklusif, bahkan ketika mereka bertindak dalam arah yang berlawanan. Namun, bagaimana keseimbangan ini terbentuk masih belum diketahui dalam banyak kasus. Misalnya, bagaimana pulau-pulau diferensiasi pada kromosom terkait-kelamin, selama proses introgresi adaptif, memengaruhi kinerja reproduksi jenis kelamin heterogametik? Dampak ini melampaui secara langsung mendorong kemandulan (atau bahkan ketidakmampuan total) yang berpotensi mencakup perubahan fisiologis endogen lainnya dalam sel-sel reproduksi, dan/atau karakteristik seksual sekunder (Walsh et al. 2016 ).
Studi kami juga berkontribusi dalam mengidentifikasi mekanisme utama introgresi adaptif yang dijelaskan dalam literatur, yang mencakup berbagai proses yang terjadi bersamaan dengan gaya divergensi. Namun, konsekuensi spesifik dari beberapa mekanisme ini masih belum sepenuhnya dipahami. Misalnya, bagaimana pergeseran genetik dan seleksi penyeimbangan memengaruhi terjadinya pulau diferensiasi di luar kromosom yang tertaut dengan jenis kelamin? Menjawab pertanyaan ini akan membantu memperjelas relevansinya terhadap introgresi adaptif dan divergensi, yang masih belum sepenuhnya dipahami. Selain itu, dalam kasus di mana spesies penjajah lebih suka kawin dengan spesies asli yang memiliki sifat yang lebih beradaptasi dengan kondisi lingkungan setempat, dapatkah introgresi adaptif memberikan keuntungan bagi spesies penjajah melalui seleksi seksual? Studi kami menunjukkan bahwa pola ini jauh lebih umum dalam literatur dibandingkan dengan yang sebaliknya. Namun, masih ada kelangkaan contoh yang menjelaskan hubungan antara seleksi seksual dan introgresi adaptif dari spesies asli ke spesies penjajah. Terakhir, seberapa luas penyebaran geografis alel yang diintrogresi di luar zona kontak? Meskipun ada beberapa contoh menarik, hanya sedikit bukti yang ada tentang perluasan geografis alel yang diintrogresi di luar zona kontak. Peramalan alel jarang diuji dan hanya didukung secara tidak langsung. Meskipun demikian, penting untuk mengklarifikasi tingkat spasial sebenarnya dari introgresi adaptif di luar zona kontak, karena hal ini dapat memberikan wawasan berharga untuk strategi konservasi, pengelolaan spesies, dan prediksi respons evolusioner terhadap perubahan lingkungan.
Akhirnya, sementara tinjauan kami mengidentifikasi tren utama dalam interaksi antara introgresi adaptif dan karakteristik organisme struktural, peran arsitektur genomik memerlukan eksplorasi lebih lanjut. Variasi struktural, seperti inversi, fusi, dan translokasi, dapat memfasilitasi introgresi adaptif dengan mempertahankan wilayah yang diseleksi; namun, hal itu juga bertindak sebagai penghalang aliran gen dengan membatasi rekombinasi. Demikian pula, ketidakcocokan genetik, khususnya pada kromosom seks, sangat memengaruhi pola introgresi dengan memodulasi kebugaran hibrida. Penelitian di masa mendatang harus memprioritaskan analisis genomik resolusi tinggi untuk mengukur efek ini secara sistematis di seluruh taksa dan untuk memperjelas keseimbangan antara kekuatan yang memfasilitasi dan membatasi fitur genomik struktural dalam introgresi adaptif.
Mengatasi bias penelitian di seluruh kelompok taksonomi, seperti representasi berlebihan mamalia, angiospermae, dan genera yang telah dipelajari dengan baik seperti Mus , Iris , Heliconius , atau Anopheles , sangat penting untuk pemahaman yang lebih seimbang tentang introgresi adaptif. Memperluas cakupan taksonomi dalam penelitian mendatang dapat mengungkap pola dan mekanisme yang saat ini kurang dieksplorasi.
4.7 Implikasi Terapan
Temuan kami menyoroti pentingnya introgresi adaptif sebagai mekanisme yang dapat meningkatkan ketahanan dan kemampuan beradaptasi spesies, khususnya di bawah tekanan lingkungan, yang sangat penting bagi biologi evolusi dan strategi konservasi. Bukti penelitian tentang fungsi introgresi adaptif di seluruh gradien kompleksitas—dari prokariota hingga mamalia—menegaskan relevansinya dalam konteks ekologi dan lapisan biologis yang beragam, dari adaptasi genomik hingga perilaku. Misalnya, pola introgresi adaptif yang teridentifikasi pada autosom di samping diferensiasi pada kromosom terkait jenis kelamin menunjukkan keseimbangan evolusi yang bernuansa antara aliran gen dan divergensi spesies. Pengetahuan ini secara langsung berlaku dalam mengelola dampak hibridisasi dalam biologi konservasi, khususnya untuk spesies di habitat yang terfragmentasi atau berubah di mana menjaga keragaman genetik adaptif sangat penting.
Studi ini juga menunjukkan potensi terapan yang signifikan dalam memprediksi respons evolusioner terhadap perubahan lingkungan yang cepat, seperti perubahan iklim dan hilangnya habitat. Prevalensi introgresi adaptif yang diamati yang dimediasi oleh seleksi penyeimbangan, terutama dalam konteks ekologi seperti ketahanan hama atau adaptasi terhadap suhu ekstrem, menggambarkan bagaimana alel yang diintrogresi dapat memungkinkan spesies untuk beradaptasi dengan cepat terhadap lingkungan baru. Wawasan ini dapat dimanfaatkan dalam genetika konservasi untuk mendukung ketahanan populasi, menawarkan kerangka kerja untuk mengantisipasi respons adaptif dan mengelola aliran gen introgresif dalam konteks konservasi.
5 Kesimpulan
Meta-analisis kami tentang tren pengetahuan tentang introgresi adaptif menunjukkan bahwa revolusi genomik, pasca-2012, membantu memperjelas karakteristik introgresi adaptif dan memberikan bukti untuk kemunculan simultan kekuatan penyeimbang divergensi. Kami menemukan bahwa jumlah karakteristik introgresi adaptif yang dipublikasikan meningkat dari bakteri ke mamalia melintasi gradien kompleksitas, yang secara progresif terjadi pada lebih banyak tingkat organisasi biologis (dari genom ke ekologi), yang dipengaruhi oleh kendala yang diberlakukan oleh kekuatan yang berbeda. Dengan menguji kecenderungan dalam literatur yang dipublikasikan, kami menyoroti beberapa karakteristik introgresi adaptif dan beberapa kekuatan divergensi yang hidup berdampingan, yang menunjukkan bahwa keduanya tidak saling eksklusif bahkan ketika keduanya bertindak dalam arah yang berlawanan. Keseimbangan ini harus dimediasi oleh kondisi lingkungan terlepas dari kompleksitas struktural organisme dan tingkat organisasi biologis tempat kekuatan penyeimbang ini beroperasi. Kesenjangan pengetahuan terus berlanjut pada mekanisme di balik keseimbangan antara kekuatan konvergen dan divergen dan bagaimana mereka mendorong kapasitas spesies untuk beradaptasi dengan perubahan lingkungan. Wawasan ini memiliki implikasi terapan yang penting, terutama untuk strategi konservasi yang bertujuan mempertahankan keragaman adaptif di bawah tekanan lingkungan.