Abstrak
Hudsonius hudsonius (Cypriniformes: Leuciscidae), umumnya dikenal sebagai spottail shiner, adalah spesies ikan cyprinoid kecil yang ditemukan di sebagian besar Amerika Utara. H. hudsonius secara tradisional dianggap sebagai salah satu spesies paling basal dan plesiomorfik di antara ikan notropin, yaitu, ikan yang secara tradisional ditempatkan di atau terkait dengan genus besar polifiletik Notropi s, termasuk tetapi tidak terbatas pada Alburnops , Cyprinella , Ericymba , Lythrurus , Miniellus dan Paranonotropis . Sifat basal H. hudsonius telah diperkuat lebih lanjut oleh studi molekuler baru-baru ini. Osteologi ikan notropin telah difokuskan pada wilayah kerangka tertentu, termasuk kranium, gigi faring dan rumus gigi, atau kerangka ekor. Yang lain telah bertujuan untuk menentukan hubungan evolusi menggunakan karakteristik osteologis. Namun, tidak ada penelitian yang berfokus pada osteologi H. hudsonius secara khusus atau menekankan osteologi intraspesifik dari satu spesies notropin. Dengan demikian, penelitian ini bertujuan untuk mendeskripsikan dan menganalisis karakter osteologi utama H. hudsonius , menyoroti varian osteologi utama dalam dan di antara 15 populasi dari Alberta, Northwest Territories, Manitoba dan Ontario. Perhatian khusus diberikan pada elemen rahang oral, suspensorium, seri operkular, aparatus brankial, regio hyoid, seri circumorbital, korset pektoral, korset pelvis, kerangka kaudal dan tengkorak. Analisis saya terhadap osteologi H. hudsonius mengungkapkan variasi di semua regio yang diperiksa kecuali korset pelvis. Diferensiasi regional antara populasi timur dan barat khususnya terbukti dalam derajat fusi korset pektoral, fusi parhypural dan hypural pertama dari kerangka kaudal dan bentuk urohyal. Hasil analisis ini memberikan gambaran umum tentang osteologi spesies notropin basal dan menyoroti daerah mana yang rentan terhadap variasi. Analisis ini juga menimbulkan pertanyaan lebih lanjut tentang klasifikasi taksonomi H. hudsonius saat ini berdasarkan perbedaan regional yang diamati di sini.
1. PENDAHULUAN
Hudsonius hudsonius Clinton, 1824 , umumnya dikenal sebagai spottail shiner, adalah spesies ikan cyprinoid asli Amerika Utara. Ikan ini tersebar luas dari Amerika Serikat bagian timur dan sejauh utara Northwest Territories di Kanada (Froese & Pauly, 2023 ; Page & Burr, 2011 ). Mereka adalah ikan kecil, panjangnya tidak lebih dari 147 mm (Gidmark & Simons, 2014 ). H. hudsonius ditemukan di berbagai habitat air tawar, termasuk danau, sungai dan anak sungai, dan memiliki pola makan omnivora (Gidmark & Simons, 2014 ; Nelson & Paetz, 1992 ). Ciri khas H. hudsonius meliputi tubuh yang agak terkompresi, warna biru hijau di bagian punggung dan warna perak di bagian lateral. Tanda yang paling jelas adalah titik hitam besar di pangkal sirip ekor (Page & Burr, 2011 ).
Bahasa Indonesia: Dalam analisisnya tentang osteologi Notropis atherinoides Rafinesque, 1818 dan 77 spesies Notropis Rafinesque, 1818 (sl) lainnya, Coburn ( 1982 ) menganggap H. hudsonius sebagai salah satu anggota genus yang paling plesiomorfik sebagaimana yang didefinisikan saat itu, berdasarkan beberapa karakteristik morfologi dan perilaku yang mencerminkan gaya hidup generalis. Meskipun ia melakukan analisis filogenetik untuk beberapa taksa, H. hudsonius tidak disertakan dan tidak ditetapkan sebagai spesies saudara. Sejak saat itu, studi molekuler telah memberikan dukungan filogenomik untuk sifat basal H. hudsonius . Menggunakan sekuens dari gen sitokrom b mitokondria, Mayden dkk. ( 2006 ) mengkaji hubungan filogenetik dari 168 spesies notropin shiner yang berbeda. Analisis yang dihasilkan menempatkan H. hudsonius dalam klade Hudsonius Girard, 1856 basal, monofiletik , yang mana H. hudsonius merupakan saudara dari klade yang berisi Hudsonius altipinnis Cope, 1870 dan Pteronotropis cummingsae Meyers, 1925. Hudsonius membentuk trikotomi dengan kelompok yang terdiri dari tiga spesies dari genus polifiletik Pteronotropis Fowler, 1935 , dan kelompok besar yang oleh para penulis dijuluki sebagai ‘klade notropin’, meliputi sebagian besar genus/spesies yang secara tradisional ditempatkan dalam Notropis (sl). Meskipun monofili klade Hudsonius milik Mayden et al. ( 2006 ) menghadapi beberapa tantangan (Schönhuth et al., 2018 ; Stout et al., 2022 ), penempatan basal H. hudsonius secara konsisten didukung (Mayden et al., 2006 ; Schönhuth et al., 2018 ; Stout et al., 2022 ).
Penelitian terdahulu tentang osteologi H. hudsonius telah berada dalam konteks studi filogenetik yang lebih luas (Coburn, 1982 ; Mayden, 1989 ) atau studi yang berfokus pada karakteristik osteologis spesifik di antara cyprinoids Amerika Utara lainnya, seperti kerangka kaudal atau gigi faring (Buhan, 1972 ; Eastman & Underhill, 1973 ). Meskipun sifat basal H. hudsonius , penelitian belum berfokus dan menjelaskan osteologinya secara spesifik. Lebih jauh lagi, penelitian belum berfokus pada variasi osteologis intraspesifik pada H. hudsonius atau spesies notropin lainnya. Makalah ini menjelaskan osteologi H. hudsonius dan mengidentifikasi fitur-fitur osteologis yang tunduk pada variabilitas. Dengan menguraikan osteologi takson ini, saya tidak hanya akan menguraikan osteologi spesies notropin basal plesiomorfik tetapi juga menentukan bagian kerangka H. hudsonius mana yang rentan terhadap variabilitas dan mengungkap pola variasi ini yang mungkin ada di antara populasi yang berbeda.
2 BAHAN DAN METODE
2.1 Pernyataan etika
Pertimbangan etika dan kehati-hatian tidak diperlukan untuk penelitian ini, karena penelitian ini hanya menggunakan spesimen museum yang diawetkan dalam etanol atau gliserin, yang semuanya dikumpulkan sebelum penelitian dimulai. Tidak ada hewan hidup yang ditangani atau dilibatkan dalam bagian mana pun dari penelitian ini.
2.2 Bahan
Spesimen H. hudsonius yang digunakan dalam penelitian ini dikatalogkan di University of Alberta Museum of Zoology Ichthyology Collection, Edmonton, Kanada, dan University of Manitoba Ichthyology Collection, Winnipeg, Kanada. Saya menggunakan total 72 spesimen yang diambil sampelnya dari 15 populasi berbeda di Alberta (11), Manitoba (2), Ontario bagian barat (1) dan Northwest Territories (1). Empat sampel populasi dari koleksi University of Alberta dibersihkan dan diwarnai sebelum penelitian ini dan disimpan dalam 100% gliserin. Semua spesimen yang tersisa awalnya diawetkan dalam alkohol dan kemudian dibersihkan dan diwarnai mengikuti protokol Taylor dan Van Dyke ( 1985 ).
2.3 Pengukuran dan analisis
Pengukuran ( n =54), hitungan meristik ( n= 54) dan rumus gigi ( n =43) didokumentasikan menurut Hubbs dan Lagler ( 2004 ). Hitungan sisik dikumpulkan menurut Armbruster ( 2012 ), sedangkan panjang pra-anal, pra-pektoral dan pra-panggul diukur mengikuti Habib et al. Hitungan vertebra ( n = 42 ) mencakup empat centra Weberian dan centrum gabungan dari kerangka kaudal (Fink & Fink, 1981 ). Hitungan pori ( n =24) dari kanal sensorik supraorbital, supratemporal, infraorbital, preopercular mandibular dan postocular commissure diambil dari spesimen yang dibersihkan dan diwarnai dan dihitung menurut Illick ( 1956 ).
Sebanyak 50 spesimen yang telah dibersihkan dan diwarnai dianalisis. Foto untuk gambar osteologi diambil dengan mikroskop stereo Zeiss Stereo Discovery.V8 dengan lensa okuler Carl Zeiss 44.403 6-9000 (8×) dan berbagai lensa (Zeiss Achromat S 0,3× FWD 236 mm, Zeiss Plan Apo S 0,63× FWD 81 mm dan Zeiss Plan Apo S 1,0× FWD 60 mm), dengan paket NIS-Elements F 2.20, versi 5.03. Terminologi osteologi secara umum mengikuti Coburn ( 1982 ) dan Conway ( 2011 ) kecuali dinyatakan lain. Terminologi yang berkaitan dengan sistem kanal sensorik mengikuti Reno (1966). Sebanyak 20 spesimen yang telah dibersihkan dan diwarnai dibedah lebih lanjut, dengan membuang apparatus brankialnya. Semua foramen diidentifikasi mengikuti Coburn (1989) dan Conway ( 2011 ). Analisis saya difokuskan pada rahang oral ( n = 45), suspensorium ( n = 45), seri operkular ( n = 39), aparatus brankial ( n = 22), regio hyoid ( n = 46), seri circumorbital ( n = 39), aparatus Weberian ( n = 49), korset pektoral ( n = 45), korset pelvis ( n = 45) dan kerangka kaudal ( n = 50). Saya juga menganalisis elemen tengkorak, membagi analisis saya menjadi regio etmoid ( n = 49), regio orbital ( n = 49), regio otik dan oksipital ( n = 20) dan regio basaranial ( n = 20). Analisis saya terhadap aparatus brankial, regio otik, oksipital, dan basaranial dibatasi oleh jumlah spesimen yang dapat saya bedah.
3 HASIL
3.1 Daerah etmoid
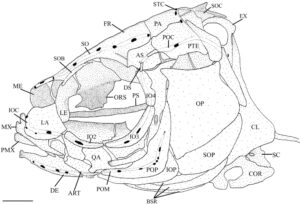
Blok atau kompleks ethmoid adalah elemen tunggal median yang membentuk bagian anterior rongga hidung. Menurut Conway ( 2011 ), blok ethmoid memiliki bagian mesetmoid (ME) dan supraetmoid (SE) yang menyatu. Ia memiliki takik anterior yang dalam yang mengakomodasi kinetmoid (K) dan dasar yang diperluas yang terletak pada strip tulang rawan, planum ethmoidale (Harrington, 1955 ), yang memisahkannya dari vomer (VO). Bagian mesetmoid membentuk bagian ventral kompleks ethmoid. Bentuknya oval dan menyempit di bagian tengah, membentuk septum hidung. Bagian supraetmoid membentuk daerah dorsal kompleks ethmoid, yang menutupi rongga hidung. Ia pipih di bagian dorsal dan tampak menyempit ke arah ujung posterior dalam pandangan dorsal (Gambar 1 ).
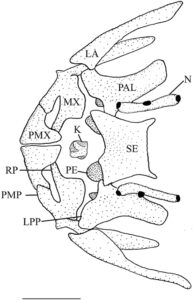
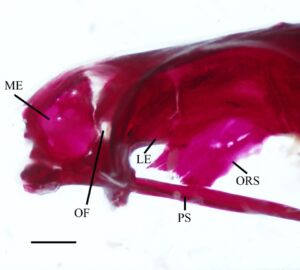
Etmoid lateral (LE) mengelilingi daerah anterior rongga mata dan membentuk bagian posterior rongga hidung. Tulang-tulang ini berartikulasi dengan blok ethmoid di bagian anterior, frontal (FR) di bagian dorsal, orbitosphenoid (ORS) di bagian posterior, dan vomer serta parasphenoid (PS) di bagian ventral. Dalam pandangan ventral (2), tulang-tulang ini memiliki margin median tebal yang menyempit menjadi satu set sayap lateral yang lebar dan datar. Permukaan ventral cekung di bagian anterior, dan margin posterior membentuk tonjolan. Ketika dilihat dari samping, ethmoid lateral menyempit ke arah daerah ventral (3). Bagian ventral dari margin anterior ethmoid lateral membentuk rak yang diarahkan ke anterior, sedangkan sudut anterodorsal dipisahkan dari bagian mesetmoid kompleks ethmoid oleh tulang rawan. Etmoid lateral membentuk margin posterior foramen olfaktorius (OF), sedangkan septum nasal dari blok ethmoid membentuk margin anterior. Pada empat spesimen yang diperiksa, foramen olfaktorius tercakup seluruhnya dalam ethmoid lateral (Gambar 2 ). Foramen orbitonasal terlihat tepat di bawah foramen olfaktorius, di sepanjang tepi posteriornya.
Preetmoid terletak di antara vomer dan kompleks etmoid. Preetmoid berbentuk bulat, nodul mengeras yang dikelilingi oleh tulang rawan dan mengarah ke lateral. Vomer adalah tulang dermal datar yang terletak di bagian ventral kompleks parasphenoid dan etmoid. Preetmoid paling lebar di bagian anterior dengan tepi lateral bulat dan satu prosesus posterior runcing yang terletak di antara etmoid lateral. Ujung anterior preetmoid memiliki lekukan yang dalam dan bulat. Nasal adalah tulang tubular yang terletak di bagian lateral dari bagian supraetmoid dari blok etmoid. Masing-masing memiliki tiga pori (Gambar 1 ).
3.2 Wilayah orbit
Orbitosphenoid adalah tulang median tunggal yang berartikulasi dengan frontal di bagian dorsal, ethmoid lateral di bagian anterior dan pterosphenoid (PTS) di bagian posterior (Gambar 3 ). Margin dorsalnya terbagi menjadi lamella kiri dan kanan yang bergabung dalam satu tonjolan median yang memanjang ke bagian ventral ke parasphenoid. Sudut anteroventral tonjolan medial bersandar pada perpanjangan tulang rawan dari ethmoid lateral. Pada tiga spesimen, sudut posteroventral menyatu dengan parasphenoid (Gambar 3 ). Tepi posterior orbitosphenoid membentuk foramen oval dengan ethmoid lateral. Foramen ini tidak ada pada tiga spesimen yang diperiksa. Orbitosphenoid membentuk margin anterior foramen optik.
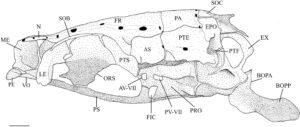
Pterosphenoid adalah elemen endochondral cekung ventral yang membentuk daerah posterior orbit (Gambar 3 ). Ini berartikulasi dengan orbitosphenoid anterior, frontal dan autosphenotic (AS) dorsolateral dan prootic (PRO) posterior. Pterosphenoid tidak berartikulasi satu sama lain medial tetapi membentuk margin posterior foramen optik dan bagian anterior foramen hypophyseal. Sudut posteroventral pterosphenoid membentuk tepi depan fossa hyomandibular anterior. Di H. hudsonius , ada empat foramen di pterosphenoid, mengakomodasi cabang vena orbitonasal (FBON), saraf troklearis (FIV), cabang oftalmik superfisial saraf wajah (FVIIS) dan cabang oftalmik saraf trigeminal (FVS) (Gambar 4 ). Identifikasi keempat foramen ini secara tentatif didasarkan pada Coburn ( 1982 ).
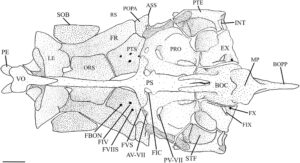
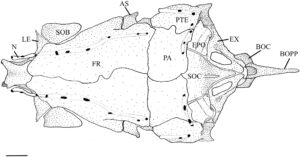
Frontal adalah tulang terbesar dari kranium, baik tumpang tindih atau tumpang tindih di bawah yang berlawanan secara medial (Gambar 5 ). Ini tumpang tindih dengan bagian supraethmoid dari blok ethmoid, ethmoid supraorbital dan lateral di anterior (Gambar 5 ) dan orbitosphenoid dan pterosphenoid di posterior (Gambar 4 ). Margin posterolateralnya berartikulasi dengan autosphenotic, dan margin posterior tumpang tindih dengan parietal (PA). Ketika dilihat secara dorsal (Gambar 5 ), ia menyempit di anterior, dengan titik terlebar berada di tempat frontal menyentuh autosphenotic. Frontal memiliki rak orbital yang ditempatkan di ventral dan menjorok ke lateral yang terletak di atas dan mengikuti kurva orbit, menyempit di anterior. Rak orbital berartikulasi di posterior dengan proses postorbital autosphenotic (POPA) (Gambar 4 ).
3.3 Daerah telinga dan oksipital
Tulang autosfenotik adalah tulang endokondral berpasangan. Tulang ini berartikulasi dengan pterosfenoid di bagian anterior, bagian frontal di bagian dorsal, bagian parietal di bagian posterodorsal, bagian pterotik (PTE) di bagian posterior, dan bagian prootik di bagian ventral. Batas ventral tulang autosfenotik membentuk setengah dari faset hiomandibula anterior dan sebagian dari faset posterior. Di bagian dorsolateral dari faset anterior terdapat prosesus postorbital yang berorientasi miring. Ujung posterior prosesus ini ditarik keluar menjadi spina autosfenotik (ASS) yang tumpul, pendek, dan melengkung ke posterior (Gambar 4 ). Pada satu spesimen, spina autosfenotik tajam dan memanjang (Gambar 6 ).
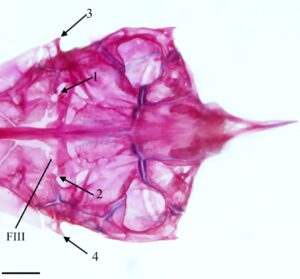
Prootik adalah tulang endokondral besar dengan bentuk heksagonal. Ia berartikulasi dengan pterosphenoid di anterior dan basioccipital (BOC) di posterior (Gambar 4 ). Ia menyentuh autosphenotik dan pterotik, berkontribusi pada faset hyomandibular anterior dan posterior. Ia membentuk dinding anterior fossa subtemporal (STF), berartikulasi dengan epiotik (EPO) dan eksoksipital (EX) di dalam fossa (Gambar 4 ). Prootik terbagi menjadi dua lamela di sepanjang tepi medial. Lamela dorsal bertemu dengan yang berlawanan di medial dan menyimpang ke anterior untuk membentuk bagian posterior foramen hipofisis. Lamela ventral membentuk bagian dari dinding miodome posterior, dengan tepi medial berartikulasi dengan parasphenoid. Bagian posterior membesar menjadi kapsul. Bukaan anterior bilik trigeminal-wajah (AV-VII) biasanya dibatasi oleh prootik di posterior dan pterosphenoid di anterior; namun, bukaan ini sepenuhnya terbungkus dalam prootik pada satu spesimen. Pada tiga spesimen, bukaan anterior bilik trigeminal-wajah terbungkus sepenuhnya dalam prootik di satu sisi dan dibagi dengan pterotik di sisi lainnya (Gambar 4 ). Pada tujuh spesimen, saya juga mengamati penopang tulang yang terbentuk di atas foramen trigeminal, tepat di lateral sayap parasphenoid, setidaknya pada satu sisi (Gambar 6 ). Bukaan posterior bilik trigeminal-wajah (PV-VII) selalu terbungkus dalam prootik.
Foramen okulomotor (FIII) ditemukan di sudut anteromedial prootik (Gambar 6 ). Dari spesimen yang daerah oksipitalnya dapat saya periksa, foramen okulomotor paling sering dibatasi oleh prootik dan pterosphenoid. Pada lima spesimen, foramen okulomotor sepenuhnya terbungkus dalam prootik di satu sisi dan berbagi dengan pterotik di sisi lain. Pada lima spesimen lain, foramen okulomotor terbungkus dalam prootik saja (seperti yang terlihat pada Gambar 6 ).
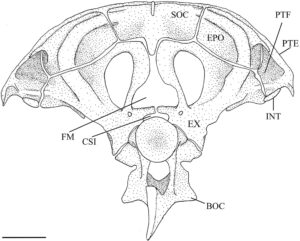
Pterotik adalah elemen besar yang tumpang tindih dengan autosfenotik di bagian anterior dan tumpang tindih dengan parietal di bagian dorsal (Gambar 5 ). Ini membentuk dinding lateral fossa subtemporal (Gambar 4 ), berartikulasi dengan epiotik dan eksoksipital di dalam fossa. Secara ventrolateral, pterotik membentuk faset hyomandibular posterior. Sebuah duri pendek menonjol dari margin posterolateral pterotik, yang bergabung secara ligamen ke posttemporal (PTT) dan suprakleithrum (SCL). Dalam pandangan posterior, ini membentuk dinding lateral fossa posttemporal (PTF) (Gambar 7 ).
Tulang parietal adalah tulang dermal persegi panjang pada permukaan dorsal tengkorak (Gambar 5 ). Tulang ini tumpang tindih dengan tulang supraoksipital (SOC) dan epiotik di bagian posterior, tulang autosfenotik dan pterotik di bagian lateral, dan tumpang tindih dengan tulang frontal di bagian anterior.
Eksoksipital adalah tulang endokondral besar yang berartikulasi dengan basioksipital di bagian medial, pterotik di bagian lateral, epiotik di bagian dorsolateral, supraoksipital di bagian dorsal, dan prootik di bagian anterior (Gambar 4 dan 7 ). Dalam pandangan ventral, ia membentuk bagian medial fossa subtemporal (Gambar 4 ). Dalam pandangan posterior, ia membentuk dinding ventromedial fossa posttemporal (Gambar 7 ). Fitur eksoksipital yang paling menonjol adalah cincin tulang yang melengkung ke arah posterior dan membungkus foramen oksipital lateral yang besar. Cincin tersebut memiliki prosesus runcing pada tepi posteriornya yang diarahkan ke takik yang terbentuk di supraneural kedua (SN2) vertebra Weberian kedua. Cincin ini bertemu dengan kebalikannya di bagian posteromedial, hampir bersentuhan untuk membentuk foramen magnum (FM) yang bulat dan berbentuk segitiga. Eksoksipital bergabung secara medial untuk membentuk dasar foramen magnum dan atap cavum sinus impar (CSI) (Gambar 7 ). Dalam pandangan posterior, eksoksipital membentuk dasar tonjolan yang dibentuk oleh kanalis semisirkularis yang berlanjut ke epiotik dan berakhir di sepanjang tepi lateral supraoksipital (Gambar 7 ). Dalam pandangan ventral, foramen vagal (FX) dan glossopharyngeal (FIX) sepenuhnya terbungkus dalam sudut posteromedial eksoksipital. Foramen vagal terletak posterior terhadap, dan jauh lebih kecil daripada, foramen glossopharyngeal (Gambar 4 ).
Epiotik adalah tulang endochondral berpasangan yang berartikulasi dengan supraoccipital di bagian medial, eksoccipital di bagian ventral dan parietal di bagian dorsal (Gambar 7 ). Ia berkontribusi pada bagian dorsal fossa subtemporal. Dalam pandangan posterior, ia membentuk dinding dorsomedial fossa posttemporal dan dasar cavum sinus impar (Gambar 7 ). Ia memiliki tonjolan menonjol yang dibentuk oleh kanalis semisirkularis di sepanjang permukaan posteriornya (Gambar 7 ). Sebuah rak yang menghadap lateral dibentuk sepanjang margin dorsolateral tonjolan ini agar posttemporal dapat berartikulasi dan membentuk atap fossa posttemporal (Gambar 7 ). Sebuah tuberkulum yang lemah juga diamati pada margin medial tonjolan.
Supraoksipital adalah tulang endokondral median yang mengartikulasikan epiotik di bagian lateral dan eksoksipital di bagian ventral (Gambar 7 ), sedangkan parietal tumpang tindih dengan tepi anterodorsalnya (Gambar 5 ). Dalam pandangan dorsal, supraoksipital memiliki bentuk segitiga dan cembung di bagian anterior dan cekung di bagian dorsomedial dengan puncak sagital kecil. Supraoksipital membentuk tepi dorsal foramen magnum. Permukaan posterolateralnya memiliki tonjolan yang dibentuk oleh kanalis semisirkularis di kedua sisi yang berakhir sebelum puncak sagital.
Interkalar (INT) adalah osifikasi dermal berpasangan. Mereka berbentuk bulan sabit dan terletak di sepanjang tepi posterior pterotik, dekat dengan sutura pterotik-eksoksipital (Gambar 4 dan 7 ). Mereka berukuran kecil pada H. hudsonius .
3.4 Daerah Basranial
Basioksipital adalah elemen tunggal yang berposisi posteroventral (Gambar 5 dan 7 ). Ia berartikulasi dengan eksoksipital di bagian dorsal, prootik di bagian anterior, dan parasphenoid di bagian ventral (Gambar 4 dan 7 ). Di bagian posterior, ia berartikulasi dengan centrum vertebra pertama. Prosesus faringeal (BOPP) ditemukan di bagian ventral dan mengarah ke posterior (Gambar 3 ). Ia terkompresi secara lateral, melebar ke bagian posterior, dan memiliki bentuk hampir persegi panjang dengan ujung posterior bulat yang tidak sepenuhnya mencapai os suspensorium (OS). Di bagian anterior prosesus faringeal terdapat lempeng pengunyah cekung yang menghadap anteroventral dengan sayap persegi yang menonjol ke bagian ventrolateral, tepi posterior yang sempit, dan tepi anterior yang berlekuk (Gambar 4 ). Di bagian dorsal lempeng terdapat lengkungan penyangga yang menghubungkan prosesus faringeal dan lempeng ke bagian ventrolateral basioksipital (Gambar 7 ). Sebuah kanal besar terbentuk di antara lengkungan. Margin anteroventral basioccipital berlekuk, membentuk sebagian dinding dan atap myodome posterior. Dua lamella ventral memanjang di atas tepi anterior basioccipital dan margin dorsal prootik (Gambar 4 ). Lamella ini tumpang tindih dengan ujung posterior parasphenoid untuk membuat lantai ventral yang membantu menutup myodome posterior bersama dengan parasphenoid. Pada salah satu spesimen yang diperiksa, lamella ventral tidak ada, tetapi penutupan myodome posterior ini masih diamati.
Parasphenoid adalah tulang tunggal dan terpanjang dari kranium. Tulang ini dapat dibagi menjadi tiga bagian. Pertama adalah batang anterior, yang melewati antara rongga mata kiri dan kanan (Gambar 3 ) dan berakhir di bagian dorsal vomer (Gambar 4 ). Kedua, ada dua sayap menaik yang diarahkan ke bagian dorsal yang menjahit tepi anteromedial prootik dan membentuk tepi anterior foramen untuk arteri karotis interna (FIC). Ketiga adalah ujung posterior yang bercabang. Tepi medial percabangan berbatasan dan membentuk sebagian atap miodome posterior tertutup (CPM) (Gambar 4 ).
3.5 Aparatus brankial
Faringobrankial (PCH) adalah elemen paling dorsal dari apparatus brankial (Gambar 8 ); hanya ada dua pasang faringobrankial, yang mewakili faringobrankial 2 dan 3. Ini adalah tulang-tulang kecil dengan tepi yang tidak teratur. Faringobrankial kedua (PBR2) memiliki bentuk yang kira-kira subsirkular dan disambung oleh tulang rawan ke epibranchial pertama (EB1) sepanjang tepi anterolateralnya dan epibranchial kedua sepanjang tepi posteromedial. Faringobrankial ketiga lebih memanjang daripada yang kedua, menutupi seperempat ujung posterodorsal faringobrankial kedua, dan memiliki lebar yang sama. Tepi mediannya sigmoidal, sedangkan tepi lateralnya setengah lingkaran. Pada empat apparatus brankial yang diperiksa, tepi medialnya memiliki takik yang dalam (Gambar 9a ). Faringobrankial kedua dan ketiga dihubungkan secara medial oleh tulang rawan. Tepi posterolateral dari pharyngobranchial ketiga dihubungkan oleh tulang rawan ke epibranchial ketiga, sedangkan tepi posteriornya dihubungkan ke epibranchial keempat (EB4), juga oleh tulang rawan.
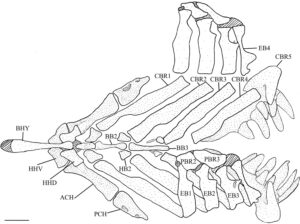
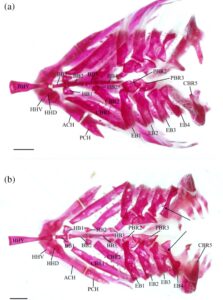
Empat pasang epibranchial hadir, menempel pada ceratobranchial (CBR) di ujung distalnya. Semua epibranchial sedikit cekung di bagian dorsal dan memiliki takik dangkal dan prosesus unsinatus yang menonjol ke posterior di sepanjang margin posterior. Tiga epibranchial pertama menyempit ke arah ujung dorsalnya di sebelah prosesus unsinatus. Epibranchial pertama lebih sempit daripada yang kedua atau ketiga, dan margin anteriornya agak cembung. Takik posterior dan prosesus unsinatusnya berkembang lemah. Epibranchial kedua memiliki margin anterior lurus, dengan takik posterior dan prosesus unsinatus yang berkembang lebih baik. Epibranchial ketiga lebih pendek daripada epibranchial pertama dan kedua. Ujung distalnya lebih lebar daripada ujung proksimalnya, dengan takik posterior yang menonjol dan prosesus unsinatus yang panjang dan ramping yang tumpang tindih dengan epibranchial keempat. Prosesus yang tipis dan lebar ditemukan pada permukaan dorsal epibranchial ketiga, yang bermula secara lateral ke prosesus uncinatus. Pada empat apparatus branchial yang diperiksa, prosesus dorsal ini hanya ada pada satu sisi. Epibranchial keempat lebih ramping daripada yang lain. Ujung distalnya lebih sempit, dan menyempit di bagian tengah, tidak seperti epibranchial lainnya (Gambar 8 ). Prosesus dorsal yang tipis dan mengarah ke medial dengan ukuran dan posisi yang bervariasi terkadang dapat ditemukan di dekat prosesus uncinatus epibranchial ketiga (Gambar 9b ). Prosesus dorsal ini hanya terlihat pada satu epibranchial pada 11 spesimen yang diperiksa dan ada pada keduanya atau sama sekali tidak ada pada 6 spesimen. Prosesus uncinatus epibranchial keempat lebar, datar, dan bundar, terletak lebih dekat ke ujung distal, dan mengarah ke posterodorsal.
Ceratobranchial merupakan elemen lengkung insang yang terbesar. Keempat yang pertama cekung di bagian ventral, sempit dan memanjang, menyentuh epibranchial masing-masing di ujung distalnya. Ceratobranchial pertama (CBR1) merupakan yang terpanjang, sedangkan ceratobranchial kedua (CBR2), ketiga (CBR3) dan keempat (CBR4) semakin pendek. Ujung proksimal lebih sempit daripada ujung distal untuk ceratobranchial pertama dan ketiga dan lebih lebar untuk ceratobranchial kedua. Kedua ujung sama untuk ceratobranchial keempat. Bagian proksimal ceratobranchial pertama sedikit menyempit sekitar sepertiga tulang sebelum melebar ke bagian distal.
Gigi faring ditopang pada ceratobranchial kelima (Gambar 10 ). Tulang ini dapat dibagi menjadi dua tungkai: tungkai anterior (ventral) dan tungkai asenden (posterior). Tungkai anterior (ANL) berjalan sejajar dengan ceratobranchial lainnya tetapi lebih berposisi ventral. Ketika dilihat dari lateral, tungkai anterior menyempit ke anterior menjadi titik tumpul dengan flens ventral (VF) yang berjalan sepanjang margin lateral dan meruncing ke ventral. Tungkai asenden (ASL) lebih lebar dan melengkung ke arah dorsal. Kedua tungkai bertemu dalam sudut anterior tumpul lancip (AAC). Tungkai asenden melengkung lemah, dengan sudut posterior yang lemah dan tidak berkembang. Dalam satu set lengkung faring yang diperiksa, sudut anterior tungkai asenden lebar dan bulat (Gambar 11 ).
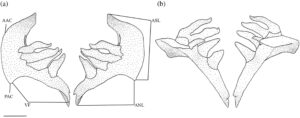

Gigi faring panjang dan berbentuk kerucut dengan ujung yang mengait. Mayoritas spesimen memiliki rumus gigi 1,4–4,1 (Tabel 3 ), yang dihitung pada 25 set lengkung seratobranchial. Rumus gigi yang paling umum yang menyimpang dari 1,4–4,1 adalah 2,4–4,2 (lima set) diikuti oleh 0,4–4,0 dan 2,4–4,1 (keduanya empat set) dan 1,4–4,0 (tiga set). Rumus 1,4–4,2 dan 0,3–4,0 hanya diamati satu kali masing-masing. Pada salah satu set yang diperiksa, tampak ada tiga baris gigi, bukan dua pada lengkung kiri. Namun, hal ini kemungkinan besar disebabkan oleh kegagalan beberapa gigi fungsional, dalam hal ini, gigi pertama dan keempat pada baris utama, untuk tanggal ketika penggantinya mengalami ankilosis dengan lengkung (Evans & Deubler, 1955 ).
Tiga pasang tulang nodular berbentuk tidak beraturan, yaitu tulang hipobranchial (HB), terletak di antara tiga tulang ceratobranchial dan basibranchial (BB) pertama. Pasangan pertama adalah yang terbesar, diikuti oleh yang kedua dan ketiga. Pasangan tulang hipobranchial pertama (HB1) dan kedua (HB2) memiliki prosesus ventral yang pendek dan tumpul. Pasangan tulang hipobranchial ketiga (HB3) memiliki serangkaian prosesus ventral yang panjang dan ramping yang mengait di bagian anterior.
Tiga basibranchial adalah tulang yang tidak berpasangan yang ditemukan di sepanjang garis tengah aparatus brankial. Basibranchial pertama (BB1) adalah yang terpendek dan meruncing ke arah posterior dalam pandangan dorsal. Dalam pandangan lateral, ia memiliki bentuk segitiga dan ujung anterior yang menyempit. Sebuah lekukan di kedua sisi di bagian tengah tulang mengakomodasi artikulasi pasangan hipobranchial pertama. Basibranchial kedua (BB2) dan ketiga (BB3) berbentuk batang. Basibranchial kedua menyempit di bagian tengah, dan ujung anterior dan posterior memiliki lebar yang sama. Basibranchial ketiga adalah yang terpanjang dan ramping, meruncing ke arah posterior.
Basihyal (BHY) adalah tulang panjang yang tidak berpasangan dan merupakan elemen paling anterior dari apparatus branchial. Tulang ini melengkung ke atas di bagian dorsal dan memiliki ujung tulang rawan anterior. Titik terdalam dari lengkungan tersebut berada di ujung anterior, sekitar sepertiga dari bagian bawah, dengan tonjolan ventral. Ujung posterior basihyal berartikulasi dengan basibranchial pertama dan memiliki dua tuberkel di kedua sisinya sebagai ligamen untuk mengikatnya ke hipohia ventral (HHV). Batang basihyal menyempit di bagian posterior. Ujung posteriornya bulat dan lebih lebar dari titik tersempit tulang tetapi masih lebih sempit dari bagian anterior.
Bahasa Indonesia : Pada salah satu apparatus branchial yang saya periksa (MZF 680), terdapat empat basibranchial, bukan tiga (Gambar 9a ). Pada spesimen ini, basibranchial keempat, paling posterior adalah yang terpanjang, diikuti oleh basibranchial kedua, ketiga, dan pertama. Hypobranchial terletak di antara basibranchial pertama dan kedua, ketiga, dan keempat, dan di ujung basibranchial keempat. Basibranchial pertama pada dasarnya sama seperti yang dijelaskan di atas. Basibranchial kedua terlebar di ujung anterior, meruncing ke posterior. Ujung anterior dan posterior basibranchial ketiga sama, dan bagian tengahnya menyempit. Basibranchial keempat memiliki bentuk keseluruhan yang seragam, dan ujung anteriornya sedikit lebih lebar.
3.6 Daerah hyoid
Tulang interhyal (IHY) merupakan tulang silindris kecil yang melekat pada permukaan medial persimpangan hyomandibula-symplectic dengan tulang rawan di salah satu ujungnya, sedangkan ujung lainnya berartikulasi pada sebuah divot yang terdapat pada tepi posterodorsal dari ceratohyal posterior (PCH) (Gambar 12 ).
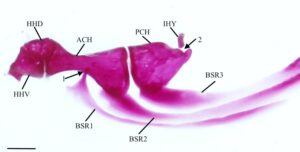
Ceratohyal posterior (Gambar 13 ) berbentuk segitiga. Ia memiliki dua foramen superfisial pada tepi dorsalnya yang terhubung di bawah jembatan tulang. Bukaan anterior lebih besar daripada bukaan posterior. Di posterior foramen terdapat lekukan tempat interhyal bersendi. Lekukan ini terlihat lebih dalam pada lima spesimen yang diperiksa (Gambar 12 ).
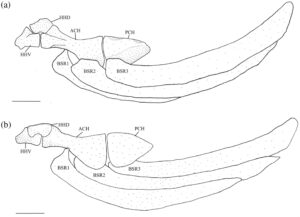
Ceratohyal anterior (ACH) paling lebar di tepinya yang bersentuhan dengan ceratohyal posterior. Bagian tengahnya meruncing, lalu melebar menjadi kepala yang sedikit bengkok di ujung anteriornya. Bagian anterodorsal berartikulasi dengan hipohyal dorsal (HHD), sedangkan bagian anteroventral berartikulasi dengan hipohyal ventral. Sebuah lekukan kecil ditemukan di sisi medial pada titik tersempit dari ceratohyal anterior tempat sinar branchiostegal (BSR) paling anterior berartikulasi.
Tulang hipohia ventral lebih besar daripada tulang hipohia dorsal. Tulang hipohia dorsal merupakan tulang bersudut yang berartikulasi dengan tulang hipohia ventral dan tulang hipohia anterior di sepanjang permukaan ventralnya. Bagian posterior tulang hipohia terletak di antara tulang hipohia dorsal kanan dan kiri dan melekat secara ligamen ke setiap elemen. Sebuah tuberkel terlihat di sepanjang tepi dorsomedial tulang hipohia dorsal. Tulang hipohia ventral berbentuk segitiga, berartikulasi dengan tulang hipohia anterior dan tulang hipohia dorsal. Sebuah tuberkel kecil terlihat di permukaan anterolateralnya.
Ceratohyal anterior dan hipohyal dorsal dan ventral secara bervariasi membentuk foramen hipohyal (Gambar 14 ). Pada 26 batang hyoid yang diperiksa, foramen hipohyal dibentuk oleh ketiga elemen (Gambar 14a ), yang memberikan ceratohyal anterior penampilan yang bercabang dangkal. Pada 18 batang hyoid yang diperiksa, foramen hipohyal dilingkupi dalam hipohyal dorsal dan ventral, tetapi ceratohyal anterior tidak berkontribusi (Gambar 14b ). Pada dua batang hyoid yang diperiksa, foramen hipohyal hanya dilingkupi dalam hipohyal ventral (Gambar 14c ).
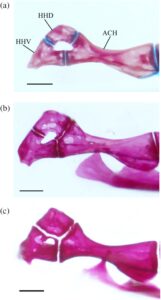
Ketiga sinar branchiostegal (Gambar 13 ) semuanya serupa bentuknya. Branchiostegal pertama (BSR1) adalah yang terkecil dan paling ramping, meruncing ke anterior menjadi kepala pendek dan tumpul yang berartikulasi dengan permukaan medial ceratohyal anterior. Pada 16 batang hyoid yang diperiksa, kepalanya memanjang (Gambar 12 ). Basis ventral ke kepala ini diperluas dengan margin anterior yang lebar dan bulat, proyeksi clupeoid (Coburn, 1982 ). Kepala branchiostegal kedua dan ketiga jauh lebih lebar daripada yang pertama, sedangkan kepala branchiostegal kedua adalah yang terbesar. Branchiostegal kedua (BSR2) berartikulasi dengan sisi lateral ceratohyal anterior, sedangkan yang ketiga (BSR3) berartikulasi dengan sisi lateral ceratohyal posterior. Sinar branchiostegal ketiga adalah yang terluas.
Urohyal memiliki dua sayap ventral horizontal dan satu sayap dorsal vertikal. Ketiga sayap meruncing bersama di anterior menjadi leher silinder yang menyempit, bercabang menjadi dua proses yang dipisahkan oleh takik yang jelas. Proses ini menempel pada hipohyal ventral melalui ligamen. Pada 27 batang hyoid yang diperiksa, sayap dorsal sedikit lebih panjang dari sayap ventral dan memiliki margin posterior setengah lingkaran yang lebar (Gambar 15a ). Pada 19 batang hyoid yang diperiksa, sayap ventral lebih panjang dari sayap dorsal, dan sayap dorsal itu sendiri memiliki bentuk segitiga dengan sudut posterodorsal bundar (Gambar 15b ). Satu urohyal memiliki bentuk yang jelas: sayap ventral jauh lebih panjang dari sayap dorsal, dan margin posterior sayap dorsal hampir lurus (Gambar 16 ).
3.7 Seri Operkular
Empat tulang membentuk seri operkular (Gambar 17 ). Operkulum (OP) adalah tulang terbesar. Operkulum cekung di bagian medial dan sudut anterodorsal ditarik menjadi lengan pendek dan tumpul, prosesus operkular, yang tumpang tindih dengan margin posterior hyomandibula (HYP). Tepi anteroventral operkulum cembung dan tumpang tindih dengan preoperkulum (POP). Tepi anterodorsal cekung dangkal dan lebih pendek dari tepi posteroventral. Tepi posterodorsal juga sigmoidal dengan sudut posterodorsal yang jelas. Faset operkular ditemukan tepat di bawah prosesus anterodorsal di sisi lateral. Ini adalah soket dalam yang menghubungkan operkulum ke hyomandibular.
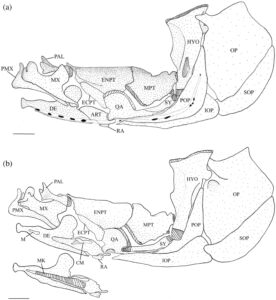
Suboperkulum (SOP) berbentuk bulan sabit dengan tepi anterodorsal yang lurus, sedangkan tepi posteroventral berbentuk cembung. Sudut anterodorsal suboperkulum ditarik ke dalam prosesus anterodorsal pendek yang berartikulasi dengan sisi medial interoperkulum (IOP). Sinar branchiostegal terletak di sepanjang sisi medial suboperkulum saat berada dalam keadaan diam.
Interoperkulum adalah tulang berbentuk baji dengan tepi dorsal cekung yang berada di bawah preoperkulum. Tepi posterior interoperkulum berbentuk cembung. Interoperkulum paling lebar di ujung posteriornya, meruncing ke anterior menjadi titik halus yang melekat secara ligamentum ke retroartikular (RA). Di bagian medial, tulang ini menyentuh interhia dan seratohia posterior, yang tumpang tindih dengan tepi dorsolateral interoperkulum.
Preoperkulum adalah tulang melengkung dengan tungkai vertikal dan horizontal yang jelas; tungkai vertikal sedikit lebih panjang. Persambungan hyomandibula-symplectic-interhyal terletak di sepanjang tepi lateral tungkai vertikal. Flensa posterior hyomandibula tumpang tindih dengan tepi anterodorsal tungkai vertikal. Tepi anterodorsal tungkai horizontal berartikulasi di bawah alur ventral kuadratus (QA).
3.8 Suspensorium
Hyomandibula (Gambar 17 ) adalah elemen paling posterior dari suspensorium. Paling lebar di ujung dorsalnya, membentuk kepala dengan kondilus anterior dan posterior, yang berartikulasi dengan tengkorak di fosa anterior dan posterior yang sesuai yang dibentuk oleh autosphenotic, pterosphenoid dan prootic. Hyomandibula meruncing ke arah ventral, berakhir di tutup tulang rawan yang berartikulasi dan membentuk sambungan dengan metapterygoid (MPT), simplektik (SY) dan interhyal. Margin anteroventral hyomandibula berartikulasi dengan metapterygoid di dua titik: titik ventral menjadi bagian dari sambungan dengan simplektik dan interhyal, dan titik dorsal berartikulasi dengan hyomandibula tepat di ventral dari ujung sayap anterior (AWH). Sayap anterior berasal dari margin anterior kepala hyomandibula dan memanjang kira-kira dua pertiga dari panjang tulang. Pada 17 suspensoria yang diperiksa, sayap anterior menyempit ke arah ujung ventral dan ditarik ke titik anteroventral yang jelas (Gambar 18a ). Sayap posterior dimulai di bawah margin posterior kepala dan berakhir ke arah ujung ventral tulang, tumpang tindih dengan lengan vertikal preoperkulum. Kondilus operkular menonjol ke posterior, berartikulasi dengan operkulum. Pada empat spesimen yang diperiksa, tonjolan adduktor yang kurang berkembang terdapat pada tingkat kondilus operkular (Gambar 18a ), sedangkan pada satu spesimen, tonjolan adduktor berkembang menjadi flensa yang jelas.
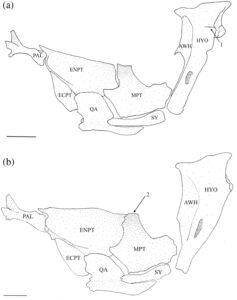
Simplektik (Gambar 17 ) adalah tulang yang terkompresi, berbentuk batang, melengkung ke arah dorsal dengan ujung anteroventral dan posterodorsal tulang rawan yang bundar. Ujung posterodorsal adalah bagian dari sambungan hyomandibula-simplektik-interhia. Tepi dorsal ujung anteroventral menyisip di bawah alur yang dibentuk oleh kuadratus. Biasanya, tepi dorsal simplektik terpisah dan tidak berartikulasi dengan metapterygoid. Namun, pada 19 suspensoria yang diperiksa, tepi dorsal simplektik berartikulasi dengan metapterygoid.
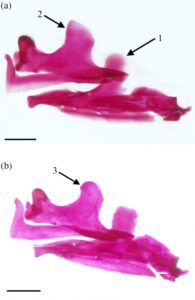
Metapterygoid adalah tulang lebar yang melengkung ke arah dorsomedial. Tulang ini biasanya tumpang tindih dengan margin anterior endopterygoid (ENPT) secara keseluruhan (Gambar 17 dan 18b ). Pada 11 suspensoria yang diperiksa, metapterygoid sebagian tumpang tindih dengan endopterygoid, hanya menutupi sudut posteroventral (Gambar 19a ). Margin posterior metapterygoid memiliki dua kepala kartilaginosa yang berbeda. Yang lebih kecil berartikulasi dengan sambungan yang dibentuk oleh simplektis, hyomandibula, dan interhia, sedangkan yang lebih besar berartikulasi dengan margin anterior hyomandibula. Tepi dorsal cekung, sedangkan sudut anterodorsal ditarik ke dalam proses yang sejajar dengan tepi dorsal ektopterigoid (ECPT). Proses anterodorsal biasanya memiliki bentuk persegi yang lebar (Gambar 18b ). Pada tujuh suspensoria yang diperiksa, prosesus anterodorsal berbentuk bulat (Gambar 19a ), sedangkan pada tujuh spesimen lainnya, prosesus anterodorsal berbentuk tipis dan runcing (Gambar 19b ). Pada satu spesimen, prosesus anterodorsal tidak jelas (Gambar 17 ). Prosesus yang kurang berkembang ditemukan di sudut posterodorsal metapterygoid.
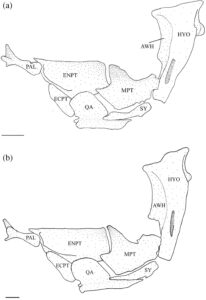
Kuadrat memiliki bilah setengah lingkaran dorsal yang tepi anterior dan dorsalnya tumpang tindih dengan ektopterigoid dan endopterygoid (Gambar 17a ). Sudut anteroventral membentuk kondilus bundar yang berartikulasi dengan anguloartikular. Di bagian ventral bilah terdapat prosesus posterior beralur ventral yang mengakomodasi simplektik dan berartikulasi dengan tepi dorsal tungkai horizontal preoperkulum (Gambar 17b ). Basis ventral kuadrat memiliki fosa yang terletak di bagian medial.
Endopterigoid lebih besar daripada metapterygoid. Ia melengkung ke arah medial. Sisi lateralnya tumpang tindih dengan ektopterigoid di bagian anterior, kuadratus di bagian ventral, dan metapterygoid di bagian posterior (Gambar 17 ). Sudut anterodorsal endopterygoid memiliki soket yang berartikulasi dengan ujung posterior palatina (PAL). Di bagian dorsal dari faset terdapat prosesus anterior yang runcing dan diarahkan ke medial yang memanjang ke atas palatina. Soket lain yang ditempatkan di bagian ventral terdapat di bagian posterior prosesus ini, yang mengakomodasi tepi posterior etmoid lateral. Pada satu spesimen, prosesus anterior tereduksi dan belum sempurna. Sebuah flens terbentuk di sepanjang tepi lateral soket ini dan memanjang ke posteroventral di sepanjang sisi lateral endopterygoid (Gambar 17a ). Tepi dorsal endopterygoid kira-kira lurus, dan tepi posterior sejajar dengan sudut paling posterior dari bilah kuadratus (Gambar 17b ).
Ektopterigoid tumpang tindih dengan endopterygoid di sepanjang tepi dorsomedialnya dan tumpang tindih dengan kuadratus di bagian posterolateral. Bentuknya kira-kira oval (Gambar 17 ).
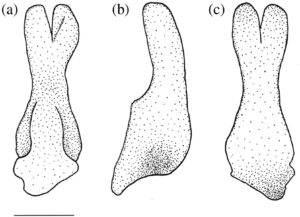
Palatina adalah tulang paling anterior dari suspensorium. Batangnya berbentuk silinder, dan ujung posteriornya memiliki tutup tulang rawan yang berartikulasi dengan endopterygoid. Tuberkulum dorsolateral diposisikan tepat di anterior tutup tulang rawan. Ujung anterior bercabang tiga menjadi tiga proses yang diarahkan ke dorsal, ventral, dan lateral (Gambar 1 dan 19 ). Proses dorsal berada di tepi medial dan menyempit menjadi ujung yang panjang, halus, dan diarahkan ke dorsal. Proses ventral juga diposisikan secara medial dan lebih pendek dan lebih lebar daripada proses dorsal, dengan bentuk yang runcing. Proses dorsal dan ventral membentuk alur yang dalam di atas bagian posterolateral preethmoid (PE). Proses lateral diposisikan secara lateral dan ditarik menjadi sayap yang lebar dan datar yang menahan proses rostral premaxilla (RP).
3.9 Rahang oral
Dentary (DE) (Gambar 17 ) adalah tulang terbesar pada rahang bawah. Ujung anteriornya melengkung ke medial untuk bertemu dengan bagian yang berlawanan. Tepi posterodorsalnya melebar ke prosesus koronoid yang datar dan bulat. Pada sembilan set rahang mulut yang diperiksa, prosesus koronoid lebih sempit dan lebih memanjang dibandingkan yang lain (Gambar 20a ). Tepi posterior dentary meruncing ke titik yang berakhir di anterior retroartikular. Tepi anteromedial menyatu dengan mentomeckelian (M), sebuah osifikasi di ujung anterior kartilago Meckel. Seluruh ujung anterior anguloartikular (ART) masuk ke ujung posterior dentary, dengan kartilago Meckel menutupi permukaan medial kedua elemen (Gambar 17b ).
Tulang anguloartikular berartikulasi dengan tulang retroartikular di sepanjang tepi ventralnya (Gambar 17 ). Tulang ini memiliki soket yang dalam di ujung posteriornya, tempat tulang kuadrata berartikulasi. Sudut posteroventral membentuk prosesus posterior tumpul yang memanjang di bawah tepi anteroventral tulang kuadrata. Tulang retroartikular adalah tulang kecil yang berartikulasi dengan tulang anguloartikular di sepanjang tepi dorsalnya. Tulang ini memiliki bentuk memanjang yang tidak teratur.
Koronomeckelian (CM) ditemukan pada tepi medial anguloartikular (Gambar 17b ). Ini adalah perpanjangan dari osifikasi ujung posterior kartilago Meckel. Koronomeckelian memiliki takik anterior yang dalam dan flens tengah yang diarahkan ke medial.
Premaksila (PMX) adalah tulang ramping, melengkung ke samping, dan berbentuk L. Tulang ini membentuk seluruh celah rahang atas. Prosesus asenden cekung posterior yang panjang menyentuh bagian yang berlawanan di garis tengah (Gambar 1 ). Prosesus asenden lebar di pangkal dan meruncing menjadi titik tumpul. Premaksila meruncing ke posterior menjadi batang pipih lateral dengan ujung bulat yang melengkung ke arah ventral.
Maxilla (MX) (Gambar 1 dan 17 ) adalah tulang melengkung ke samping yang tumpang tindih dan menyokong premaxilla, bertindak sebagai tuas untuk mendorong premaxilla saat mulut terbuka. Ujung anterior maxilla memiliki dua prosesus: prosesus premaxillary (PMP) dan prosesus rostral posteriornya (Gambar 1 ). Prosesus premaxillary pendek, datar dan melengkung ke medial, tumpang tindih dengan prosesus asendens premaxilla saat mulut tertutup (Gambar 1 ). Prosesus rostral panjang dan seperti kait dan melengkung ke medial di mana ia bergabung secara ligamen dengan yang berlawanan. Di posterior prosesus rostral dan premaxillary terdapat dua kondilus, yang berfungsi sebagai titik poros tempat maxilla berputar. Saat mulut terbuka, kondilus medial menahan preethmoid, sedangkan kondilus lateral menahan prosesus lateral palatina (LPP) (tidak diilustrasikan dalam Gambar 1 ). Sebuah tuberkel kecil dan runcing terlihat di sepanjang tepi anterolateral rahang atas tepat di belakang prosesus premaksila untuk insersi adduktor mandibula. Di belakang prosesus premaksila, tepi dorsal rahang atas berlekuk dalam, menciptakan leher yang mengarah ke prosesus asendens. Prosesus asendens rahang atas lebar dan datar dengan sudut bundar dan berorientasi anterodorsal (Gambar 20b ). Pada 11 rahang mulut yang diperiksa, prosesus asendens berbentuk persegi dan ditarik ke titik anterodorsal yang jelas (Gambar 20a ). Pada dua set rahang mulut yang diperiksa, prosesus asendens melebar dan persegi panjang, sedangkan pada satu set, prosesus asendens lebar dan bundar dengan sudut anterodorsal yang bengkok. Ujung posterior rahang atas meruncing menjadi ujung tumpul dengan flens posterior bundar pada tepi ventral. Flens posterior ini tumpang tindih dengan ujung posterior premaksila.
Kinethmoid (Gambar 21 ) adalah elemen endochondral tunggal di bagian tengah. Ketika mulut tertutup, ia terletak di antara tulang maksila, posterior terhadap premaksila dan anterior terhadap blok ethmoid. Kinethmoid kira-kira tiga kali lebih tinggi daripada lebarnya. Kinethmoid menyempit antara bagian dorsal dan ventral. Bagian dorsal kinethmoid bercabang dan melengkung ke anterior, sedangkan bagian ventral memiliki tuberkulum yang terletak lateral di kedua sisi dan fossa yang terletak di posterior yang mengarah ke prosesus ventral tunggal yang tumpul dan terletak di anterior.
3.10 Seri sirkumorbital 3.10
Rangkaian sirkumorbital mengelilingi mata dan meliputi supraorbital dan lima infraorbital (IO). Supraorbital adalah elemen berbentuk oval, melengkung ke arah dorsal yang terletak di sepanjang tepi dorsal mata (Gambar 4 dan 5 ). Tepi medialnya cembung, dan tepi lateralnya lurus. Dalam pandangan dorsal, tepi ini paling lebar di bagian tengah, menyempit di bagian posterior menjadi ujung yang tumpul (Gambar 5 ).
Infraorbital adalah serangkaian lima tulang pipih seperti pelat yang mengelilingi margin ventral dan posterior orbit (Gambar 22 ). Lacrimal (LA) adalah elemen pertama dan terbesar dari seri infraorbital. Ini menutupi bagian dari kapsul hidung dan maksila dan disangga oleh palatina. Tingginya kira-kira sama dengan lebarnya, dengan lima tepi: dorsal, orbital, ventral, posterior dan anterior. Secara keseluruhan, lakrimal kira-kira persegi, tetapi bulat di anterior. Pada 14 spesimen yang diperiksa, tepi anteriornya lebih dalam. Tepi ventralnya cembung dan lebih panjang dari tepi anterior. Tepi posteriornya pendek, berbatasan dengan infraorbital kedua.
Infraorbital kedua memanjang. Infraorbital ketiga lebih panjang dan lebih lebar daripada yang kedua, dengan bentuk melengkung seperti bulan sabit. Infraorbital keempat terletak di sepanjang tepi posterior orbit dan tumpang tindih dengan bagian anterior hyomandibula. Infraorbital kelima, atau dermosphenotic (DS) (Coburn, 1982 ), melapisi spina autosphenotic dan mengecil menjadi tabung tulang.
3.11 Sistem kanal sensorik
Kanal sensorik memanjang sepanjang dan hadir pada beberapa tulang tengkorak, rahang oral, seri operkular dan seri circumorbital (Gambar 22 ). Kanal supraorbital (SO) memanjang melalui nasal (N) dan tertutup di frontal, memanjang sepanjang margin dorsal orbit. Bagian nasal biasanya memiliki tiga pori, dan jumlah total pori biasanya delapan, tetapi mencapai sembilan dalam lima spesimen (Tabel 3 ). Namun, dalam tiga spesimen, semua dari sampel yang sama (MZF 2506), bagian nasal tidak ada, dan bagian supraorbital hanya memiliki enam pori. Komisura postokular (POC), kadang-kadang disebut kanal temporal lateral (Coburn, 1982 ), tertutup dalam pterotik dan memiliki satu pori. Dalam satu spesimen, ada dua pori, bukan satu (Tabel 3 ).
Kanalis supratemporal (STC) hadir di sepanjang margin posteromedial parietal, memanjang ke pterotik dan selalu memiliki tiga pori. Kanalis mandibula preoperkular (POM) tertutup dan memanjang di sepanjang dentary, anguloartikular, dan preoperkulum. Biasanya memiliki 10 pori, 4 dari bagian dentary dan 6 di sepanjang bagian preoperkular. Jumlah totalnya dapat berkisar dari 9 (empat spesimen) hingga 11 (lima spesimen) (Tabel 3 ). Pori-pori di sepanjang bagian dentary diamati serendah tiga (satu spesimen) dan hingga lima (dua spesimen), dan pori-pori di sepanjang bagian preoperkular berkisar dari serendah lima (empat spesimen) hingga setinggi tujuh (empat spesimen) (Tabel 3 ).
Kanalis infraorbitalis (IOC) memanjang sepanjang rangkaian infraorbitalis. Biasanya terdapat tujuh pori: empat pada lakrimal, satu pada infraorbital kedua, dan dua pada infraorbital ketiga. Jumlah pori infraorbital bisa serendah enam (satu spesimen) dan setinggi delapan (dua spesimen) (Tabel 3 ). Satu spesimen memiliki tiga pori pada lakrimal, bukan empat, sedangkan dua spesimen memiliki tiga pori pada infraorbital ketiga, bukan dua seperti biasanya.
3.12 Aparatus Weberian
Aparatus Weberian (Gambar 23 ) dapat dibagi menjadi dua komponen: pars sustentaculum dan pars auditum (Bird et al., 2020 ; Bird & Hernandez, 2007 ; Coburn, 1982 ). Pars sustentaculum berasal dari empat vertebra pertama, yang telah dimodifikasi untuk menopang empat tulang pendengaran Weberian, yang membentuk pars auditum (Coburn, 1982 ).
3.12.1 Bagian sustentakulum
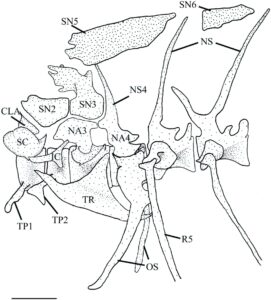
Vertebra pertama adalah centrum dengan prosesus transversal (TP1) di kedua sisi yang menjorok secara horizontal dari permukaan ventrolateral. Sisi anterior centrum berartikulasi dengan basioccipital.
Vertebra kedua sedikit lebih kecil dari yang pertama. Ia memiliki centrum amfikoel dengan prosesus transversal besar seperti bilah (TP2) yang menonjol ke arah anterolateral dan memanjang di bawah ujung anterior prosesus transversal vertebra pertama. Supraneural kedua dimodifikasi menjadi elemen berbentuk pelana yang melengkung di atas kanal saraf dan memiliki lekukan anterior tempat eksoksipital masuk.
Vertebra ketiga adalah yang terbesar dari pars sustentaculum. Ia memiliki centrum amfikoel dengan bibir ventral kecil yang menonjol di bawah vertebra kedua. Permukaan posterolateral beralur untuk menopang kaki tripus (TR). Alur ini diarahkan ke posterodorsal dan berkembang menjadi tonjolan yang memanjang sedikit di luar centrum. Lengkungan saraf ketiga (NA3) dimodifikasi dan dapat dipecah menjadi tiga komponen. Pertama adalah dasar, yang terletak pada permukaan dorsal centrum. Ia lebih lebar dari leher tetapi lebih sempit dari flens dorsal. Yang kedua adalah leher, yang menyempit dan memiliki flens yang mengembang sendiri di pangkalnya. Ketiga adalah flens dorsal, yang memanjang di atas vertebra kedua, berartikulasi dengan supraneural kedua di anterior, dengan supraneural ketiga (SN3) di dorsal dan dengan lengkung saraf keempat (NA4) di posterior.
Supraneural ketiga terdiri dari dasar yang lebar, leher yang menyempit, dan puncak dorsal yang terbagi. Dasar supraneural ketiga berartikulasi dengan supraneural kedua di bagian anterior, lengkung saraf ketiga di bagian ventral, dan spina saraf keempat (NS4) di bagian posterior. Puncak supraneural ketiga meluas ke bagian dorsal di atas sentrum vertebra kedua dan bagian posterior lengkung saraf kedua. Pada 20 aparatus Weberian yang diperiksa, puncak dorsal memanjang, menonjol lebih dari setengah lengkung saraf kedua. Puncak ganda supraneural ketiga mengecil dan memendek pada 10 aparatus Weberian yang diperiksa.
Lima supraneural datar terletak dorsal ke vertebra 3–9. Semuanya berbentuk tidak teratur dan menjadi lebih kecil di bagian posterior. Supraneural kelima (SN5) terletak dorsal ke vertebra ketiga, keempat, dan kelima, di atas puncak dorsal supraneural ketiga. Supraneural keenam (SN6) ditemukan di atas vertebra keenam. Supraneural ketujuh ditemukan di atas vertebra ketujuh; supraneural kedelapan ditemukan di atas vertebra kedelapan; dan supraneural kesembilan ditemukan di atas vertebra kesembilan. Pada lima apparatus Weberian yang diperiksa, supraneural kelima pas di antara sisi kiri dan kanan puncak ganda supraneural ketiga (Gambar 24 ).
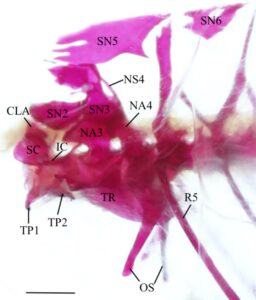
Vertebra keempat memiliki centrum amfikoel dengan lengkung saraf yang dimodifikasi dan spina neural (NS). Lengkung saraf bersifat autogen dengan dasar yang melebar dan alur lateral yang dalam yang menembus ke arah dasar. Ia berartikulasi dengan lengkung saraf ketiga di anterior dan supraneural ketiga di dorsal. Spina neural lebih pendek daripada vertebrae berikutnya dan diarahkan ke anterior. Centrum vertebra keempat menopang os suspensorium, yang terdiri dari dua lengan. Lengan luar bersifat autogen dari centrum vertebra dan berartikulasi dengan centrum vertebra keempat melalui kepala yang melebar dan melebar secara posterolateral sebelum meruncing ke belakang di anterior. Sudut paling posterior dari lengan luar terletak di bawah tulang rusuk kelima (R5). Lengan dalam os suspensorium menyatu dengan margin medial lengan luar. Ia dimulai dengan dasar yang lebar yang meruncing dan melengkung ke medial, berjalan sejajar dengan, tetapi tidak berartikulasi dengan, bagian yang berlawanan. Vertebra kelima sangat mirip dengan vertebra pasca-Weberian lainnya dan hanya berbeda pada parapofisisnya yang lebih besar dan tulang rusuk yang lebih tebal.
3.12.2 Bagian audit
Klaustrum (CLA) terletak di bagian dorsal vertebra pertama dan ditutupi oleh skaphium (SC) di bagian lateral. Klaustrum dapat dibagi menjadi faset bawah dan puncak atas. Puncak atas berbentuk segitiga, dan tepi posterior terhubung ke tepi anteroventral lengkung saraf kedua.
Skaphium terletak di sepanjang daerah posterolateral vertebra pertama. Tulang ini terdiri dari bagian yang ‘mencekung’ dan prosesus dorsal dan ventral yang terpisah dan terletak di bagian posterior. Bagian yang mencekung adalah yang terbesar, dengan lubang di sisi medial dan tuberkulum kecil di bagian tengah sisi lateral. Prosesus dorsal diarahkan ke bagian dorsal dengan ujung yang tipis dan runcing. Prosesus ventral diarahkan ke bagian anterior, lebih pendek dari lengan asenden dan melekat secara kartilaginosa ke sentrum vertebra pertama.
Intercalarium (IC) adalah tulang kecil berbentuk Y dengan satu lengan anterior dan prosesus artikulasi dan asenden posterior. Lengan anterior tumpul, melengkung ke arah anterodorsal, dan terletak lateral terhadap skaphium. Ujung posterior bercabang menjadi prosesus asenden dan artikulasi. Prosesus asenden lebih panjang dari keduanya dan mengarah ke dorsal. Lengan artikulasi mengarah ke ventral dan berartikulasi dengan permukaan lateral sentrum vertebra kedua.
Tripus adalah tulang kecil Weberian yang terbesar. Tulang ini memiliki bentuk cekung dorsal. Tripus dapat dibagi menjadi badan dengan lengan anterior dan posterior. Badannya berbentuk segitiga dengan prosesus yang diarahkan secara posterodorsal. Tripus diposisikan di sepanjang sisi lateral sentrum vertebra ketiga. Lengan tripus diratakan. Lengan anterior lebih pendek dan lebih lebar, melengkung ke arah dorsomedial dan berakhir di bagian dorsal prosesus transversal vertebra kedua dan lateral ke ujung anterior intercalarium, yang terhubung secara ligamen. Tonjolan dorsal berasal dari tepi medial lengan anterior dan berjalan ke arah dorsomedial untuk bertemu dengan sudut dorsoposterior badan. Tonjolan ini tidak ada pada dua aparatus Weberian yang diperiksa. Lengan posterior berjalan ke arah posteroventral, melengkung ke arah medial dan berakhir di antara lengan dalam dan luar os suspensorium, yang menghubungkan ke kantung renang.
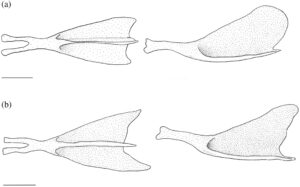
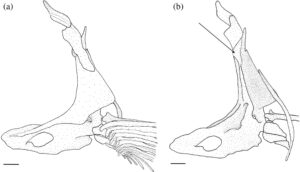
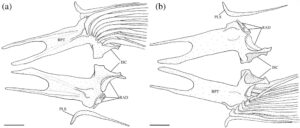
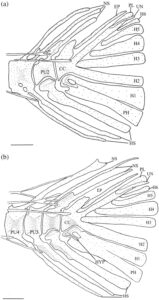
3.13 Korset dada
Posttemporal adalah elemen datar yang tumpang tindih dengan suprakleithrum di bagian ventral. Bentuknya seperti tetesan air mata, menyempit di bagian dorsal menjadi ujung yang runcing (Gambar 29 dan 30 ). Tepi anterodorsalnya menyentuh pterotik, membentuk sebagian besar atap dan tepi lateral fosa posttemporal.
Suprakleitrum adalah elemen datar yang tumpang tindih dengan posttemporal di bagian dorsal dan tumpang tindih dengan kleitrum (CL) di bagian ventral (Gambar 29 dan 30 ). Margin dorsal memiliki prosesus dorsal yang tumpul dan berkait. Pada lima korset pektoral yang diperiksa, prosesus dorsal lurus, tanpa kait. Suprakleitrum paling lebar di dasar prosesus dorsal dan meruncing menjadi ujung ventral yang tumpul dan bulat. Sudut posterodorsal operkulum tumpang tindih dengan margin anterodorsal suprakleitrum.
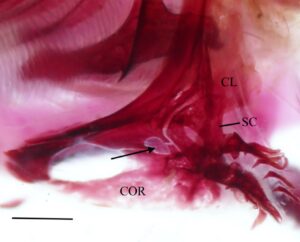
Kleithrum adalah tulang terbesar dari korset pektoral, dengan tungkai vertikal dan horizontal dengan panjang yang sama yang bertemu pada sudut tumpul (Gambar 25a ). Margin anterior tungkai horizontal bertemu dengan yang berlawanan di garis tengah. Titik tengah tungkai horizontal menyempit, sedangkan margin anterior dan posterior melebar, dan keduanya berartikulasi dengan korakoid (COR) di medial untuk membuat foramen besar. Sebuah tonjolan yang jelas membentang di sepanjang permukaan medial tungkai horizontal, meruncing ke posterior dan berakhir di dasar tungkai vertikal. Ketika dilihat dari dorsal, tepi anterolateral tungkai horizontal cekung dangkal (Gambar 26 ). Tungkai vertikal cleithrum terdiri dari pelat lateral segitiga yang lebar yang dimulai di sepanjang ujung anterior tungkai horizontal dan berakhir di ujung tungkai vertikal di tulang belakang tumpul, dan tonjolan medial yang melebar ke dorsal dan berakhir menjadi flens yang cukup berkembang tepat sebelum tulang belakang dorsal pelat lateral (Gambar 25 ). Plat lateral tungkai vertikal berartikulasi dengan tepi lateral skapula (SC), sedangkan mesocoracoid (MC) berartikulasi dengan tepi anterior punggungan medial tungkai horizontal cleithrum (Gambar 25b ).
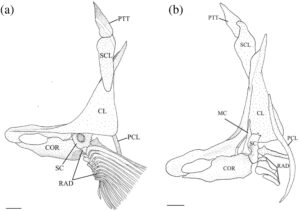
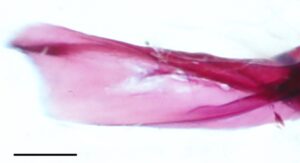
Postcleithrum (PCL) adalah tulang tipis berbentuk seperti batang, berbentuk S dengan ujung dorsal dan ventral yang runcing. Ujung dorsalnya melekat pada permukaan posteromedial lempeng lateral cleithrum. Ujung ventral postcleithrum melengkung ke arah medial (Gambar 27 ).
Korakoid melebar ke arah posterior, berartikulasi dengan kleithrum di ujung anterior dan posteriornya, membentuk sutura. Titik artikulasi posterior lebih besar daripada anterior. Di tempat artikulasi dengan kleithrum, terbentuk foramen besar. Ujung posterior korakoid berartikulasi dengan skapula, sedangkan tepi posteromedial membentuk lapisan tulang yang berartikulasi dengan mesokorakoid. Pada satu spesimen, patologi unik diamati di mana kleithrum dan korakoid bersama-sama membentuk foramen kedua yang terletak di anterior (Gambar 32 ).
Mesocoracoid (Gambar 25b ) merupakan elemen segitiga yang menyempit di bagian dorsal. Margin ventralnya berartikulasi dengan skapula di bagian posterior dan coracoid di bagian anterior. Ia memiliki prosesus bundar di sepanjang margin anterodorsal yang berartikulasi dengan tonjolan median lengan cleithrum yang menaik.
Foramen skapula besar dan diposisikan anterodorsal. Pada salah satu korset pektoral yang diperiksa, foramen skapula skapula kiri dibagi menjadi dua oleh penyangga tulang (Gambar 28 ). Margin dorsal skapula berbentuk bulat, berartikulasi dengan permukaan medial cleithrum. Tepi posterior memiliki faset yang berartikulasi dengan sinar sirip dada pertama. Sudut posterodorsal skapula berkembang menjadi flens bulat yang lemah yang berartikulasi dengan permukaan medial cleithrum. Pada tiga korset pektoral yang diperiksa, flens posterodorsal berkembang dengan kuat.
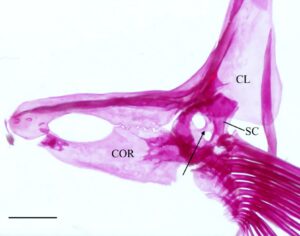
Sebanyak empat radial (RAD) menyokong jari-jari sirip dada. Dihitung dari dorsal ke ventral, radial pertama (paling dorsal) adalah yang terpendek. Bentuknya bundar dan tumpang tindih dengan tepi posterolateral skapula. Radial kedua lebar dan sedikit lebih pendek dari yang pertama dan tumpang tindih dengan tepi medial skapula. Radial ketiga adalah yang terpanjang, dan yang keempat adalah yang tertipis.
Pada sekitar 11 korset pektoral yang saya periksa, terdapat fusi lengkap cleithrum, scapula, coracoid, dan mesocoracoid menjadi satu elemen (Gambar 29 ). Spesimen-spesimen ini, selain satu yang tetap tidak menyatu, memiliki flens dorsal yang membesar pada punggung medial cleithrum (Gambar 29b ). Duri dorsal dari lempeng lateral cleithrum juga tampak memendek secara nyata pada spesimen-spesimen ini (Gambar 29b ).
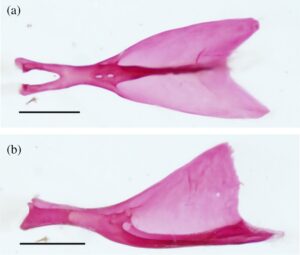
3.14 Korset panggul
Korset panggul (Gambar 30 ) terdiri dari dua basipterygia (BPT) yang disatukan oleh tulang rawan di sepanjang tepi medialnya. Kedua basipterygia cekung di bagian dorsal dan memiliki ujung anterior yang bercabang, menghasilkan dua proses yang menjorok ke anterior. Proses lateral lebih panjang dan lebih sempit daripada proses medial. Dalam pandangan dorsal, dua tonjolan dangkal membentang di sepanjang proses lateral dan medial. Tonjolan proses lateral memanjang lebih jauh ke atas. Tuberositas bulat diamati di dasar tonjolan proses medial. Ketika dilihat dari bagian ventral, tuberositas lemah diamati di ujung posterior setiap basipterygium.
Setiap basipterygium memiliki prosesus iskiadika (ISC) yang lebar, tumpul, dan melengkung ke samping yang ditemukan di posterior titik tempat sisi kiri dan kanan bergabung. Tiga radial menopang sinar setiap sirip panggul. Radius paling medial adalah elemen besar yang bengkok yang melengkung ke medial dan memiliki dasar yang melebar yang tumpang tindih dengan basipterygia di bagian ventral. Radius tengah dan lateral lebih kecil, mengartikulasikan sepanjang margin posterolateral basipterygium. Radius lateral lebih besar dari radial tengah. Belat panggul (PLS) adalah elemen berbentuk L yang bergabung erat dengan permukaan luar sinar panggul pertama. Ujung anterior belat panggul memiliki prosesus menaik yang pendek dan diarahkan ke anteroventral.
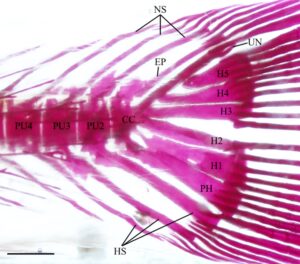
3.15 Kerangka kaudal
Seperti pada cyprinoids lain, empat vertebra terakhir dari H. hudsonius (Gambar 31 ) menopang jari-jari sirip ekor (Buhan, 1972 ). Vertebra preural pertama adalah compound centrum (CC), yang menopang epural (EP), uroneural (UN), pleurostyle (PL), enam hypural (H1–H6) dan parhypural (PH). Compound centrum memiliki duri saraf tunggal, tumpul, dan pendek. Pada salah satu kerangka ekor yang diperiksa, ada dua duri saraf pada compound centrum (Gambar 32a ). Tiga vertebra berikut adalah preural centra, masing-masing dengan satu duri saraf dan hemal (HS). Pada tujuh kerangka ekor yang diperiksa, ada dua duri saraf pada preural centrum kedua (PU2) (Gambar 33a ), sedangkan pada tiga kerangka ekor lainnya, ada dua duri saraf pada preural centrum ketiga (PU3). Pada satu kerangka kaudal, terdapat dua duri hemal pada sentrum preural ketiga, sedangkan pada kerangka lain, duri saraf dari sentrum preural kedua terbagi secara distal (Gambar 32b ). Pada spesimen lain, duri saraf dari sentrum preural kedua dan ketiga menyatu di pangkalnya (Gambar 33b ). Pada satu spesimen, sentrum preural ketiga dan keempat (PU4) tampak menyatu, sehingga tampak seperti vertebra yang membesar dengan dua duri saraf dan hemal (Gambar 34a ).
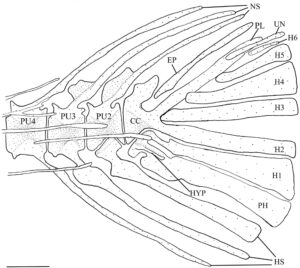
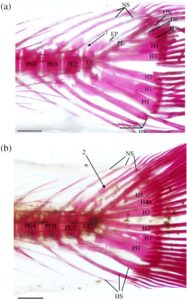
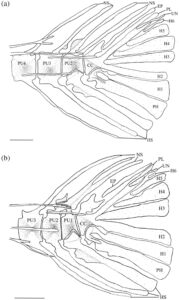
Elemen-elemen yang menyokong sinar-sinar lobus bawah sirip ekor meliputi duri-duri hemal dari centra preural kedua, ketiga dan keempat, parhypural dan hypural pertama dan kedua. Duri hemal dari centrum preural kedua bersifat autogenous. Parhypural dan hypural pertama menyatu bersama di bagian proksimal dan bersama-sama terpisah dari centrum majemuk. Pada 16 kerangka ekor yang diperiksa, parhypural dan hypural pertama sebagian menyatu bersama di bagian proksimal (Gambar 34b ), sedangkan parhypural dan hypural pertama berbatasan tetapi tidak menyatu (Gambar 34a ) pada delapan kerangka. Hypural pertama memiliki dasar yang tipis dan meruncing dan melebar di bagian distal. Parhypural memiliki hypurapophysis (HYP). Hypural kedua kira-kira berbentuk persegi panjang dan menyatu dengan centrum majemuk. Hypural dan parhypural menyokong sinar-sinar utama lobus bawah sirip ekor, sedangkan sinar prokurens ventral disokong oleh duri-duri hemal dari vertebra preural kedua, ketiga, dan keempat.
Elemen-elemen yang menyokong sinar-sinar lobus atas sirip ekor terdiri dari duri-duri saraf dari tiga centra preural ditambah epural, pleurostyle, uroneural dan hypural ketiga, keempat, kelima dan keenam, dengan hypural keenam selalu menjadi yang terkecil. Pada satu spesimen, hypural keenam tidak ada, dan hypural kelima membesar (Gambar 35 ). Epural adalah elemen ramping dan memanjang yang hampir selalu tidak menyatu dengan centrum majemuk. Pada empat kerangka ekor yang diperiksa, epural berbatasan tetapi tidak menyatu dengan centrum majemuk. Pada dua lainnya, epural menyatu dengan centrum majemuk (Gambar 35 ). Pleurostyle lebar dan kira-kira berbentuk persegi dan menyatu dengan kuat dengan centrum majemuk. Uroneural ramping dan lebih panjang dari hypural keenam. Hypurals menopang sinar-sinar utama lobus atas, sedangkan sinar-sinar prokurens disokong oleh duri-duri uroneural, epural, dan neural dari vertebra preural kedua, ketiga, dan keempat. Secara konsisten terdapat 19 sinar sirip ekor, 10 di lobus atas dan 9 di lobus bawah.
4 DISKUSI
4.1 Pengukuran dan hitungan meristik
Terdapat tingkat variasi yang tinggi dalam pengukuran dan proporsi spesimen H. hudsonius yang digunakan dalam penelitian ini. Variabilitas ini dapat diamati baik di dalam maupun di antara populasi H. hudsonius . Dalam beberapa sampel (UAMZ 213, 214, 220 dan 1525), terdapat tingkat variabilitas tingkat populasi yang tinggi dalam pengukuran yang berkaitan dengan panjang tangkai, panjang predorsal dan kedalaman tubuh (Tabel 1 ).
| Pengukuran | Minimum | Maksimum | Berarti | SD |
|---|---|---|---|---|
| Panjang standar (SL) | 31.26 | 75.5 | 58.21 | 10.58 |
| Panjang pra-analSL−1 | 0.63 | 0,77 | 0.7 | 0,03 |
| Panjang predorsalSL−1 | 0.47 | 0.56 | 0.52 | 0,03 |
| Panjang pra-panggulSL−1 | 0.46 | 0.56 | 0.52 | 0,03 |
| Panjang pra-pektoralSL−1 | 0.23 | 0,35 | 0.27 | 0,02 |
| Panjang kepala (HL)SL−1 | 0.27 | 0.34 | 0.26 | 0,02 |
| Panjang moncongSL−1 | 0,03 | 0,15 | 0,07 | 0,02 |
| Panjang moncongHL−1 | 0,59 | 0.14 | 0.26 | 0,07 |
| Panjang postorbitalSL−1 | 0,08 | 0.14 | 0.11 | 0,01 |
| Panjang postorbitalHL−1 | 0.33 | 0.49 | 0.43 | 0,04 |
| Pangkal sirip punggungSL−1 | 0.10 | 0.18 | 0.14 | 0,02 |
| Dasar sirip duburSL−1 | 0,08 | 0.17 | 0.12 | 0,02 |
| Panjang tangkaiSL−1 | 0.17 | 0.30 | 0.23 | 0,03 |
| Lebar interorbitalSL−1 | 0,06 | 0.12 | 0,09 | 0,01 |
| Lebar interorbitalHL−1 | 0.23 | 0.43 | 0,35 | 0,05 |
| Diameter mataSL−1 | 0,04 | 0.10 | 0,08 | 0,01 |
| Diameter mataHL−1 | 0.18 | 0.39 | 0.30 | 0,05 |
| Kedalaman tubuhSL−1 | 0.18 | 0.34 | 0.24 | 0,03 |
| Kedalaman tangkai ekorSL−1 | 0,06 | 0.13 | 0.10 | 0,01 |
| Panjang sirip punggungSL−1 | 0,15 | 0.27 | 0.22 | 0,02 |
| Panjang sirip duburSL−1 | 0.11 | 0.21 | 0.18 | 0,02 |
| Panjang sirip dadaSL−1 | 0.13 | 0.23 | 0.18 | 0,02 |
| Panjang sirip panggulSL−1 | 0.11 | 0.19 | 0.16 | 0,02 |
Catatan : Pengukuran disajikan dalam milimeter.
Dari hitungan meristik yang diambil pada spesimen H. hudsonius yang digunakan dalam penelitian ini, gurat sisi, sisik predorsal, dan sisik postdorsal memiliki rentang yang sejauh ini paling besar (Tabel 2 ). Hitungan sisik melingkar juga memiliki rentang yang lebih besar daripada kebanyakan hitungan meristik lainnya. Hitungan skala gurat sisi H. hudsonius dilaporkan berkisar antara 38 hingga 42 (Scott & Crossman, 1973 ), 36 hingga 42 (Page & Burr, 2011 ), dan 36 hingga 40 (Nelson & Paetz, 1992 ). Namun, rentang skala gurat sisi yang dilaporkan dalam penelitian ini (35–43) masih belum pernah terjadi sebelumnya. Hitungan skala gurat sisi dapat bervariasi dalam populasi seperti halnya di antara populasi. Misalnya, dalam dua sampel, hitungan skala garis lateral bervariasi dari 36 hingga 43 (UAMZ 214) dan 37 hingga 42 (UMAZ 3215). Yang terpenting, tidak ada perbedaan regional yang jelas antara populasi timur di Manitoba dan Ontario dan populasi barat di Alberta dan Northwest Territories. Pola serupa diamati sehubungan dengan hitungan skala circumferential, predorsal dan postdorsal, yang mencerminkan tingkat variabilitas yang tinggi baik di antara maupun di dalam populasi H. hudsonius . Rentang dan frekuensi hitungan meristik lainnya umumnya mengikuti pengamatan peneliti sebelumnya (Nelson & Paetz, 1992 ; Scott & Crossman, 1973 ). Namun, hitungan sisir insang yang saya ambil biasanya berkisar antara tiga hingga enam, kontras dengan rentang yang dilaporkan oleh Nelson dan Paetz ( 1992 ) (empat hingga sembilan). Pada satu sampel (UAMZ 1525), satu spesimen tidak memiliki sisir insang sama sekali.
| Nomor | |||||||||
|---|---|---|---|---|---|---|---|---|---|
| Pari sirip punggung | 7 (3) | 8 (41) | 9 (10) | ||||||
| Ikan pari sirip dubur | 7 (3) | 8 (45) | 9 (6) | ||||||
| Jari-jari sirip perut | 7 (3) | 8 (22) | 9 (29) | ||||||
| Jari-jari sirip dada ( n = 53) | 12 (2) | 13 (6) | 14 (28) | 15 (12) | 16 (5) | ||||
| Skala garis lateral | 35 (1) | 36 (3) | 37 (5) | 38 (10) | 39 (15) | 40 (9) | 41 (5) | 42 (3) | 43 (3) |
| Sisik predorsal | 12 (1) | 13 (2) | 14 (11) | 15 (11) | 16 (11) | 17 (13) | 18 (3) | 19 (1) | 20 (1) |
| Sisik postdorsal | 13 (1) | 14 (3) | 15 (4) | 16 (8) | 17 (13) | 18 (12) | 19 (11) | 20 (1) | 21 (1) |
| Sisik di atas garis lateral | 4 (2) | 5 (12) | 6 (30) | 7 (10) | |||||
| Sisik di bawah garis lateral | 4 (9) | 5 (43) | 6 (2) | ||||||
| Skala keliling | 24 (2) | 25 (9) | 26 (18) | 27 (11) | 28 (8) | 29 (6) | |||
| Sisik tangkai | 10 (16) | 11 (23) | 12 (15) | ||||||
| Sisir insang | 0 (1) | 3 (3) | 4 (10) | 5 (32) | 6 (8) | ||||
| Vertebra predorsal ( n = 42) | 10 (2) | 11 (21) | 12 (19) | ||||||
| Jumlah total vertebra ( n = 42) | 37 (5) | 38 (22) | 39 (10) | 40 (4) | 41 (1) |
Catatan : Jumlah spesimen untuk setiap hitungan disajikan dalam tanda kurung.
4.2 Variasi osteologi intraspesifik pada H. hudsonius
Dari penelitian ini, jelas bahwa H. hudsonius menunjukkan tingkat variabilitas osteologis yang tinggi. Kecuali pada korset panggul, saya mengamati beberapa bentuk variasi intraspesifik di hampir semua daerah yang diperiksa, meskipun pada tingkat yang berbeda-beda.
Sistem kanal sensorik H. hudsonius sebagian besar sesuai dengan deskripsi Illick ( 1956 ) dan Coburn ( 1982 ) mengenai struktur dan jumlah pori (Tabel 3 ). Namun, spesimen dari satu sampel (MZF 2506) menunjukkan sedikit pengurangan pada kanal supraorbital dan infraorbital. Pada ketiga spesimen yang diperiksa, baik bagian nasal kanal infraorbital maupun tulang nasal sendiri tidak ada. Pada satu spesimen, jumlah pori infraorbital juga berkurang, dengan tiga pori lakrimal, bukan empat seperti biasanya. Pengurangan sistem kanal sensorik telah dilaporkan pada spesies Notropis (sl) lainnya sebagai adaptasi terhadap lingkungan yang tenang dan kekeruhan rendah (Reno, 1966; Swift, 1970). Pengurangan ini sering diamati pada kanal supraorbital dan infraorbital, seperti yang terlihat di sini. Akan tetapi, pada spesies lain ini, ketika kanal supraorbital mengecil, bagian nasal tetap dipertahankan (Reno, 1966; Swift, 1970), bertentangan dengan apa yang saya amati pada spesimen H. hudsonius ini , yang diambil sampelnya dari lingkungan danau. Akan tetapi, saya tidak melihat pengurangan sensori yang sama pada spesimen dari danau lain. Hilangnya bagian nasal kanal supraorbital merupakan fenomena yang memerlukan penelitian lebih lanjut.
| Saluran sensorik | ||||
|---|---|---|---|---|
| Supraorbital | 6 (3) | 8 (16) | 9 (5) | |
| supratemporal | 3 (24) | |||
| Komisura postokular | 1 (23) | 2 (1) | ||
| Preoperkular mandibula | 9 (4) | 10 (15) | 11 (5) | |
| Infraorbital | 6 (1) | 7 (21) | 8 (2) |
Catatan : Jumlah spesimen yang menunjukkan setiap hitungan disajikan dalam tanda kurung.
Baik posisi foramen hipohia dan bentuk urohia bervariasi dalam H. hudsonius . Uyeno ( 1961 ) menggunakan posisi foramen hipohia, relatif terhadap hipohia dan seratohia anterior, sebagai karakter sistematis untuk membedakan berbagai genus cyprinoid. Posisi dan bentuk foramen hipohia juga telah ditampilkan dalam deskripsi spesies dan genus notropin (Coburn, 1982 ; Mayden, 1989 ) dan dalam studi filogenetik cyprinoid Amerika Utara (Coburn & Cavender, 1992 ). Namun, analisis saya terhadap H. hudsonius menunjukkan bahwa posisi pasti foramen hipohia bervariasi. Meskipun biasanya dibentuk oleh ceratohyal anterior dan hipohyal dorsal dan ventral, hal itu terbatas pada dua elemen terakhir yang mewakili minoritas yang cukup besar, yang diamati pada 18 dari 46 set batang hyoid yang diperiksa. Jarang, foramen hipohyal hanya dibentuk oleh hipohyal ventral. Kedua status karakter ditemukan pada spesimen dari populasi barat dan timur. Saya sering mencatat kedua status karakter pada spesimen dari sampel yang sama. Posisi foramen hipohyal jelas bervariasi dalam H. hudsonius . Variabilitas ini harus dipertimbangkan ketika menganalisis spesies notropin lainnya.
Urohyal menunjukkan dua varian berbeda terkait bentuk keseluruhan dan proporsi sayap. Salah satu varian ini, di mana sayap dorsal memiliki tepi posterior setengah lingkaran bundar yang memanjang melampaui sayap ventral, hanya diamati pada spesimen yang diambil sampelnya dari Alberta dan Northwest Territories. Varian lainnya, di mana sayap dorsal berbentuk segitiga dan lebih pendek dari sayap ventral, diamati pada spesimen yang diambil sampelnya dari Alberta, Northwest Territories, Manitoba, dan Ontario. Meskipun varian kedua sejauh ini merupakan yang paling umum pada populasi barat, campuran kedua varian tersebut sering diamati pada spesimen dari satu sampel. Varian kedua lazim ditemukan pada tiga sampel yang diambil dari Manitoba dan Ontario barat. Namun, satu urohyal dari spesimen yang diambil sampelnya dari Ontario memiliki bentuk yang berbeda: sayap ventral jauh lebih panjang daripada sayap dorsal, dan tepi posterior sayap dorsal hampir lurus. Dalam penelitiannya tentang hubungan evolusi ikan mas Amerika Utara, Mayden ( 1989 ) mencatat bahwa sekitar 40% spesimen H. hudsonius yang diperiksa memiliki sepasang prosesus lateral di pangkal leher urohyal. Namun, saya tidak mengamati varian spesifik ini pada spesimen H. hudsonius yang saya periksa.
Aparatus brankial H. hudsonius unik karena epibranchial ketiga memiliki prosesus dorsal, suatu sifat yang tidak dideskripsikan pada spesies notropin lainnya (Coburn, 1982 ; Mayden, 1989 ). Demikian pula, variabel keberadaan dan ketiadaan prosesus dorsal pada epibranchial keempat bertentangan dengan deskripsi spesies notropin lainnya, dengan prosesus dorsal selalu ada (Coburn, 1982 ; Mayden, 1989 ). Meskipun didokumentasikan hanya pada satu spesimen, keberadaan empat basibranchial alih-alih tiga merupakan varian unik yang sebelumnya tidak dilaporkan dalam literatur. Mengingat kelangkaan varian ini, kemungkinan besar ini merupakan kondisi patologis atau variasi morfologi yang berevolusi hanya pada populasi tempat spesimen tersebut diambil sampelnya (MZF 680). Rumus gigi yang biasa (1,4–4,1) yang saya amati pada H. hudsonius berbeda dengan Eastman dan Underhill ( 1973 ) dan Hubbs dan Lagler ( 2004 ), yang menemukan rumus biasa 2,4–4,2. Sebaliknya, Peer ( 1961 ) dan Nelson dan Paetz ( 1992 ) melaporkan rumus gigi H. hudsonius bervariasi, antara 1,4–4,1 dan 2,4–4,2. Akan tetapi, Eastman dan Underhill ( 1973 ) mengamati variasi rumus gigi yang sama pada H. hudsonius yang saya amati sendiri (Tabel 3 ), dengan pengecualian 0,3–4,0.
Dalam analisis mereka terhadap 42 spesies notropin asli Minnesota, Eastman dan Underhill ( 1973 ) mencatat bahwa formula gigi H. hudsonius sejauh ini paling bervariasi di antara semua spesies yang dipelajari. Sebagian besar variasi ini disebabkan oleh gigi yang hilang pada baris minor dari satu lengkung. Para penulis menyimpulkan bahwa H. hudsonius berevolusi menuju kondisi 1,4–4,1. Hasil analisis saya sendiri mungkin mencerminkan tren ini. Namun, perlu dicatat bahwa spesimen H. hudsonius yang digunakan oleh Eastman dan Underhill ( 1973 ) diambil sampelnya di Minnesota dan South Dakota. Sebagai perbandingan, spesimen saya diambil sampelnya di Alberta, Manitoba, Ontario, dan Northwest Territories. Dengan demikian, perbedaan dalam formula gigi ini juga dapat mencerminkan perbedaan regional di antara populasi H. hudsonius .
Korset panggul tidak menunjukkan varian penting di luar jumlah jari-jari sirip panggul. Sebaliknya, korset dada menunjukkan beberapa variasi penting. Fusi korakoid, mesokorakoid, cleithrum, dan skapula menjadi satu elemen adalah varian baru yang sebelumnya tidak dilaporkan oleh peneliti lain. Varian yang menyatu ini dominan pada spesimen yang diambil sampelnya dari Manitoba dan Ontario barat, dengan delapan dari sembilan spesimen menunjukkan varian ini. Fusi ini hampir selalu disertai dengan flens punggung yang membesar pada punggung medial cleithrum dan duri punggung yang memendek pada lempeng lateral cleithrum. Tidak termasuk spesimen yang diambil sampelnya dari Manitoba dan Ontario, saya mengamati varian yang menyatu hanya tiga kali pada spesimen dari Alberta. Dua dari spesimen ini juga menunjukkan flens punggung yang membesar pada punggung medial cleithrum dan duri punggung yang memendek. Tiga spesimen lain menunjukkan fusi parsial dari korset dada, satu dari Alberta dan dua lainnya dari Northwest Territories, dan juga memiliki flens dorsal yang membesar dan duri dorsal yang memendek. Pada spesimen lain, juga dari Alberta, korset dada tidak menyatu, tetapi cleithrum memiliki flens yang membesar dan duri dorsal yang sama yang biasanya diamati pada korset dada yang menyatu. Pada spesimen lain lagi, korset dada menyatu, tetapi flens dorsal mengecil, dan duri dorsal memanjang, seperti yang biasanya terlihat pada korset dada yang tidak menyatu. Hasil saya menunjukkan bahwa varian yang menyatu ini jauh lebih umum pada populasi H. hudsonius timur , tetapi tidak terisolasi pada mereka. Namun, frekuensi varian yang menyatu ini jelas berbeda antara wilayah timur dan barat.
H. hudsonius secara konsisten memiliki 19 jari-jari sirip ekor, 10 di lobus atas dan 9 di bawah, suatu kondisi plesiomorfik pada ostariophysians dan umum di antara Cypriniformes (Coburn, 1982 ; Lundberg & Baskin, 1969 ). Namun, kerangka ekor itu sendiri menunjukkan variabilitas. Varian yang paling menonjol adalah fusi parhypural dengan hypural pertama di bagian distal. Pada satu spesimen, terdapat hilangnya hypural keenam dan hypural kelima yang membesar. Fusi dan hilangnya hypural telah dicatat pada cyprinoids Amerika Utara, biasanya sebagai sifat khusus spesies (Buhan, 1972 ; Mayden, 1989 ). Namun, fusi hypural pertama dan parhypural tampaknya unik untuk H. hudsonius (Buhan, 1972 ). Dari kerangka kaudal yang saya periksa, fusi parhypural dan hypural pertama adalah variasi yang paling umum dibandingkan dengan sebagian besar spesimen di mana kedua tulang ini hanya menyatu secara proksimal di pangkalnya. Namun, tidak jarang parhypural dan hypural pertama berbatasan secara distal, tetapi tidak menyatu. Semua spesimen yang diambil sampelnya dari Manitoba dan Ontario menunjukkan fusi atau berbatasan dengan parhypural dan hypural pertama. Spesimen-spesimen ini juga berada di sisi yang lebih besar, berkisar antara 63,2 dan 75,5 mm dalam panjang standar. Namun, saya mengamati keadaan tidak menyatu pada spesimen dengan ukuran yang sama yang diambil sampelnya dari populasi barat. Saya juga mencatat fusi parhypural dan hypural pertama pada spesimen yang lebih kecil yang panjangnya di bawah 60 mm dalam panjang standar. Jadi, saya menduga fusi parhypural dan hypural pertama terkait dengan populasi dan wilayah daripada ukuran tubuh. Variasi lain dalam kerangka ekor yang saya amati, termasuk dua duri saraf atau hemal pada sentrum preural, pembelahan duri saraf, dan fusi dua sentrum preural, berdasarkan kelangkaannya, kemungkinan besar merupakan varian individual yang tidak memiliki signifikansi taksonomi.
Variasi yang diamati dalam suspensorium, rahang oral dan seri infraorbital berhubungan dengan bentuk beberapa elemen tertentu. Untuk suspensorium, itu adalah bentuk proses anterodorsal dari metapterygoid dan sayap anterior hyomandibula. Deskripsi masa lalu dari spesies notropin lain, dan memang cyprinoid, selalu menyebutkan bahwa wahana adduktor yang berbeda dan berkembang dengan baik hadir di sisi lateral hyomandibula pada tingkat kondilus operkular (Buhan, 1970 ; Coburn, 1982 ; Mayden, 1989 ; Uyeno, 1961 ). Namun, saya mengamati punggungan adduktor yang berkembang lemah hanya pada empat suspensoria yang diperiksa dan punggungan adduktor yang berkembang dengan baik hanya pada satu spesimen. Untuk rahang oral, variasi diamati dalam bentuk proses menaik dari maksila dan proses koronoid dari dentary. Untuk seri infraorbital, ini adalah tepi anterior lakrimal. Varian-varian ini tidak terisolasi pada satu populasi atau wilayah. Akan tetapi, fakta bahwa variasi semacam ini dapat ada harus diakui saat memeriksa dan membandingkan spesies notropin lainnya.
Untuk apparatus Weberian, spesimen yang diambil dari Northwest Territories secara konsisten memiliki puncak punggung memanjang yang menonjol lebih dari setengah lengkung saraf kedua. Akan tetapi, varian ini diamati pada populasi lain, meskipun lebih jarang. Dalam sampel yang diambil dari dua populasi di Alberta timur tengah (UAMZ 213, 4950), tulang predorsal pertama pas di antara sisi kiri dan kanan puncak ganda supraneural ketiga pada lima spesimen. Jika tidak, variasi pada apparatus Weberian umumnya tidak dikaitkan dengan populasi atau wilayah tertentu.
Coburn ( 1982 ) mencatat empat foramen di pterosphenoid, berbeda dengan Harrington ( 1955 ), yang hanya melaporkan dua. Pengamatan saya sendiri selaras dengan Coburn ( 1982 ). Coburn ( 1982 ) mencatat bahwa foramen untuk cabang oftalmik saraf trigeminal sering berkesinambungan dengan foramen trigeminal alih-alih terpisah. Namun, saya tidak mengamati varian ini di H. hudsonius . Namun, saya mengamati bahwa posisi foramen olfaktorius, trigeminal, dan okulomotor tampaknya bervariasi. Meskipun jarang, foramen olfaktorius kadang-kadang terbentuk secara eksklusif di dalam etmoid lateral alih-alih dibentuk oleh mesetmoid di anterior dan etmoid lateral di posterior seperti yang umum terjadi pada spesimen yang saya analisis. Demikian pula, foramen trigeminal, meskipun biasanya dibentuk oleh pterosphenoid dan prootik, dalam satu contoh hanya dibentuk oleh prootik. Pada tiga spesimen, ia dibentuk oleh prootik hanya pada satu sisi dan pterosphenoid dan prootik pada sisi lainnya. Foramen okulomotor lebih bervariasi, terbentuk antara prootik dan pterosphenoid atau hanya prootik. Varian-varian ini juga tidak terbatas pada satu populasi atau wilayah. Namun, jenis varian ini harus dipertimbangkan saat memeriksa dan membandingkan spesies notropin lainnya.
Coburn ( 1982 ) dan Mayden ( 1989 ) mendeskripsikan apa yang kemudian disebut sebagai myodome posterior terbuka (OPM). OPM adalah bukaan yang dibatasi oleh parasphenoid di bagian anterior dan basioccipital di bagian posterior. Coburn ( 1982 ) adalah orang pertama yang mendeskripsikan OPM. Mayden ( 1989 ) melangkah lebih jauh dengan menyarankan bahwa OPM adalah sinapomorfi yang dimiliki oleh semua cyprinoids Amerika Utara. Ia juga mencatat penutupan sekunder pada genera Nocomis Girard, 1856 , Campostoma Agassiz, 1855 dan Dionda Girard, 1856 dan spesies Playygobio gracilis Richardson, 1836, Notropis boucardi Günther, 1868 dan Notropis bifrenatus Cope, ( 1867 ). Dalam taksa ini, Mayden ( 1989 ) mengamati bahwa OPM hadir pada juvenil tetapi menyatu dan tertutup pada dewasa. Ia menganggap penutupan sekunder ini sebagai sifat turunan. Semua cyprinoids Amerika Utara lainnya yang diperiksa oleh Mayden ( 1989 ) tidak memiliki penutupan sekunder ini, termasuk H. hudsonius . Bertentangan dengan Mayden ( 1989 ), saya tidak mengamati OPM pada spesimen H. hudsonius yang diperiksa. Sebaliknya, saya mengamati penutupan sekunder (Gambar 4 ), CPM.
4.3 Kesimpulan
Dari analisis ini, H. hudsonius jelas merupakan spesies dengan tingkat variasi intraspesifik yang tinggi. Beberapa varian osteologis hanya diamati pada beberapa spesimen, sedangkan yang lain diamati lebih sering pada individu dari 15 populasi yang saya periksa. Beberapa varian tampaknya mencerminkan diferensiasi regional berdasarkan varian apa yang paling umum pada spesimen dari berbagai wilayah. Secara keseluruhan, tiga karakteristik osteologis tampaknya bervariasi dalam frekuensi antara populasi barat di Alberta dan Northwest Territories dan populasi timur di Manitoba dan Ontario. Yang pertama adalah fusi cleithrum, coracoid, mesocoracoid dan scapula menjadi satu elemen, disertai dengan adanya flens dorsal yang membesar pada punggung medial cleithrum dan duri dorsal yang memendek, yang ditemukan terutama pada populasi timur dan jarang diamati pada populasi barat. Yang kedua adalah sayap dorsal urohyal, yang, dalam pandangan lateral, biasanya berbentuk segitiga dan lebih pendek dari sayap ventral. Namun, sayap ini juga bisa membulat dan sedikit lebih panjang daripada sayap ventral pada populasi barat. Yang ketiga adalah rangka kaudal. Pada populasi barat, parhypural dan hypural pertama biasanya menyatu di bagian proksimal pangkalnya. Jarang terjadi penyatuan atau perlekatan parhypural dan hypural pertama di bagian distal. Sebaliknya, penyatuan atau perlekatan parhypural dan hypural pertama secara konsisten diamati pada populasi timur.
Variasi dalam karakter osteologis yang diperiksa menimbulkan pertanyaan lebih lanjut tentang diferensiasi regional dalam H. hudsonius sebagai spesies. Lebih jauh, saya akan menyarankan bahwa temuan ini menunjukkan bahwa mungkin ada perbedaan spesifik atau subspesifik di antara populasi dan wilayah ini. Namun, spesimen yang diperiksa dalam penelitian ini berasal dari bagian barat rentang yang secara tradisional dikaitkan dengan H. hudsonius (Page & Burr, 2011 ). Untuk menentukan apakah populasi ini harus dikenali sebagai spesies atau subspesies yang berbeda, penelitian lebih lanjut perlu dilakukan. Ini mungkin termasuk penelitian yang berfokus pada osteologi populasi H. hudsonius di Kanada timur dan AS. Penelitian lebih lanjut juga perlu difokuskan pada keragaman genetik H. hudsonius untuk melihat apakah perbedaan genetik antara populasi timur dan barat berkorelasi dengan perbedaan osteologis yang saya amati. Penelitian berbasis genetik harus lebih luas berfokus pada potensi perbedaan genetik H. hudsonius di seluruh rentang aslinya. Baik analisis osteologis maupun genetika diperlukan untuk memahami evolusi dan keanekaragaman regional H. hudsonius , dan apakah spesies ini, sebagaimana diklasifikasikan saat ini, dapat dibagi lagi menjadi subspesies atau bahkan spesies.


