
Abstrak
- Pergeseran tak sinkron antara serangga herbivora dan tanaman ke daerah yang lebih tinggi setelah pemanasan iklim diperkirakan akan menghasilkan interaksi tanaman-serangga yang baru. Karena herbivora generalis yang berpindah ke daerah yang lebih tinggi dapat memasukkan spesies tanaman baru ke dalam makanan mereka, komunitas tanaman di daerah yang lebih tinggi dapat mengalami peningkatan tekanan herbivori.
- Oleh karena itu, kami menyelidiki dampak orthoptera dataran rendah terhadap komunitas tumbuhan dataran tinggi dan beberapa mekanisme yang memungkinkan orthoptera tumbuh subur di dataran tinggi. Kami melakukan percobaan kebun umum di dua dataran tinggi di Pegunungan Alpen Swiss, dengan memaparkan tumbuhan dataran tinggi ke komunitas orthoptera yang berbeda. Percobaan lapangan ini dilengkapi dengan percobaan pilihan kafetaria untuk menilai preferensi diet spesies orthoptera yang termasuk dalam percobaan pertama, beserta pengukuran sifat fungsional.
- Percobaan lapangan menyoroti bahwa tanaman dataran tinggi yang termasuk dalam klade Liliopsida [= monokotil] mengalami tingkat herbivori yang lebih tinggi ketika dimakan oleh kelompok campuran orthoptera dataran rendah dan tinggi daripada ketika tanaman diserang oleh komunitas dataran tunggal. Percobaan kafetaria mengungkap konservatisme pola makan, dengan orthoptera lebih menyukai tanaman yang memiliki sifat fungsional yang sama dengan pola makan di dataran asal mereka. Selain itu, analisis sifat kami menunjukkan bahwa orthoptera dapat dikategorikan menjadi pemakan Liliopsida atau Magnoliopsida [= dikotil] berdasarkan sifat makanan dan karakteristik fungsionalnya, yang menjelaskan komplementaritas relung makan yang diamati.
- Singkatnya, orthoptera yang berpindah ke lereng atas dapat meningkatkan tekanan herbivori pada komunitas tumbuhan di dataran tinggi. Liliopsida dataran tinggi, khususnya, sangat rentan karena sifat fungsional orthoptera dataran rendah yang mendorong preferensi makan mereka terhadap kelompok tumbuhan ini.
Baca Ringkasan Bahasa Sederhana gratis untuk artikel ini di blog Jurnal.
1. PENDAHULUAN
Perubahan iklim membentuk kembali ekosistem secara global (Feeley et al., 2020 ; Pecl et al., 2017 ). Ketika suhu meningkat dengan cepat, spesies menghadapi rezim termal baru (Berg et al., 2010 ; IPCC, 2023 ). Namun, efek langsung dari pemanasan hanyalah sebagian dari tantangan tersebut. Interaksi biotik yang berubah, didorong oleh pergeseran fisiologis (Björk et al., 2007 ; Kwatcho Kengdo et al., 2022 ), ketidaksesuaian fenologi (Gordo & Sanz, 2009 ; Menzel et al., 2020 ) dan redistribusi spesies menuju lingkungan yang sesuai (Parmesan & Yohe, 2003 ; Steinbauer et al., 2018 ) dapat menimbulkan ancaman yang lebih besar. Efek tidak langsung ini dapat mengubah ekosistem secara mendalam dengan mengganggu hubungan trofik, kompetitif, simbiosis, dan parasit, yang pada akhirnya membahayakan kelangsungan spesies (Alexander et al., 2015 ; Gilman et al., 2010 ; Nomoto et al., 2024 ).
Manifestasi mencolok dari proses ini adalah pergeseran jangkauan spesies yang tidak sinkron ke arah kutub atau ketinggian yang lebih tinggi (Parmesan et al., 1999 ; Rödder et al., 2021 ). Laju pergeseran sering kali bervariasi antara kelompok taksonomi, seperti tumbuhan sesil dan serangga ektotermik (Kerner et al., 2023 ; Urban et al., 2012 ; Vitasse et al., 2021 ), yang mengarah pada interaksi tumbuhan-serangga baru di mana distribusinya sebelumnya tidak tumpang tindih (Bovay et al., 2024 ; Kerner et al., 2023 ). Sementara pengamatan sejarah alam dan pemodelan spasial dapat memprediksi ko-kemunculan baru ini, hasil dari interaksi trofik baru ini masih belum pasti. Interaksi ini kemungkinan dibentuk oleh pergeseran dalam kompatibilitas sifat tumbuhan dan serangga yang menyimpang dari hubungan saat ini (Gilman et al., 2010 ; Schleuning et al., 2020 ).
Gradien elevasi menawarkan kerangka kerja unik untuk mengeksplorasi efek tidak langsung perubahan iklim pada interaksi tumbuhan–herbivora (Körner, 2007 ; Li et al., 2009 ; Tito et al., 2020 ). Mengikuti pergeseran suhu yang cepat dalam skala geografis yang relatif kecil, adalah mungkin untuk mensimulasikan pemanasan, dengan mentranslokasi spesies ke bawah di mana suhu lebih tinggi, dan interaksi baru, yang terjadi di lingkungan yang sama atau baru (Block et al., 2022 ; Tito et al., 2020 ). Pendekatan ini sangat berwawasan untuk interaksi tumbuhan–herbivora serangga. Herbivora serangga, yang merupakan sebagian besar herbivori tumbuhan (Huntly, 1991 ; Rosenthal & Kotanen, 1994 ), memainkan peran penting dalam mengatur komunitas tumbuhan dan dinamika ekosistem (Carson & Root, 2000 ; Schmitz, 2010 ). Perubahan dalam pola makan yang disebabkan oleh pergeseran lereng di antara serangga herbivora dengan demikian dapat merestrukturisasi komunitas tumbuhan alpen (Descombes et al., 2020 ). Pada ketinggian yang lebih tinggi, tumbuhan umumnya mengalami tekanan herbivori yang lebih rendah (Galmán et al., 2018 ; Pellissier et al., 2014 ; Rasmann, Pellissier, et al., 2014 ) dan berinvestasi lebih sedikit dalam mekanisme pertahanan (Callis-Duehl et al., 2017 ; Descombes, Marchon, et al., 2017 ; Pellissier et al., 2012 ). Akibatnya, saat serangga herbivora bergeser ke lereng atas, mereka cenderung membangun komunitas tumbuhan baru ini, mengambil manfaat dari tumbuhan dataran tinggi yang pertahanannya buruk sebagai sumber makanan (Berg et al., 2010 ; Rödder et al., 2021 ).
Prediksi semacam itu seharusnya secara khusus disesuaikan dengan serangga herbivora generalis, karena mereka seharusnya lebih siap untuk mengatasi pertahanan fisik dan kimia baru daripada spesialis (Berenbaum & Zangerl, 1998 ; Jones et al., 2022 ). Orthoptera, kelompok yang didominasi oleh herbivora generalis, khususnya relevan di wilayah beriklim sedang (Joern, 1979 ). Di padang rumput pegunungan, orthoptera sering kali merupakan ordo serangga yang paling melimpah, mengonsumsi hingga 30% biomassa tanaman (Blumer & Diemer, 1996 ) dan memainkan peran penting dalam mengatur komunitas tanaman dan fungsi ekosistem (Carson & Root, 2000 ; Pitteloud et al., 2021 ). Sementara herbivora serangga generalis memiliki mekanisme detoksifikasi canggih yang memungkinkan mereka memakan spesies tanaman yang beragam (Berenbaum & Schuler, 2010 ), pola makan mereka tetap dibatasi oleh hambatan fisik (Ali & Agrawal, 2012 ; Jones et al., 2022 ). Misalnya, bentuk mandibula mereka memungkinkan mereka memakan kelas fungsional tanaman tertentu, tetapi tidak semua (Patterson, 1984 ), sementara kekuatan menggigit menentukan kemampuan mereka untuk memproses daun yang lebih keras (Ibanez et al., 2013 ). Ketahanan daun juga diketahui berbeda di antara klade tanaman, terutama antara Liliopsida [= monokotil] dan Magnoliopsida [= dikotil], yang menyebabkan tingkat makan yang berbeda pada tanaman ini (Grubb et al., 2008 ). Akibatnya, saat para generalis berpindah ke daerah yang lebih tinggi, mereka diharapkan untuk menyasar tanaman dengan ciri fungsional yang mirip dengan tanaman yang dikonsumsi di dataran rendah dan terus menyasar klade tanaman favorit mereka (Descombes et al., 2020 ).
Untuk menyelidiki mekanisme fungsional yang mendasari tantangan yang ditimbulkan oleh perpindahan naik lereng yang tidak sinkron antara tanaman dan serangga herbivora, kami meneliti herbivori dan pencocokan sifat pada komunitas orthoptera dan tanaman di lokasi dataran rendah dan tinggi. Secara khusus, kami membahas tiga pertanyaan: (1) Apakah herbivori meningkat ketika tanaman alpen terpapar pada komunitas orthoptera dataran rendah? (2) Apakah spesies orthoptera dataran rendah menunjukkan konservatisme pola makan ketika berpindah ke komunitas tanaman baru di dataran tinggi? (3) Apakah spesies orthoptera dataran rendah memiliki sifat yang membuat mereka lebih suka memakan tanaman inang baru yang menunjukkan sifat fungsional yang mirip dengan inangnya di dataran rendah? Kami memperkirakan bahwa: (1) tingkat herbivori akan meningkat ketika tanaman alpen dihadapkan pada orthoptera dataran rendah, (2) orthoptera menunjukkan konservatisme pola makan dan (3) sifat fungsional orthoptera yang terkait dengan strategi makan dan sifat tanaman yang terkait dengan pertahanan dan kualitas nutrisi mendasari konservatisme ini. Untuk menjawab pertanyaan-pertanyaan ini, kami telah menggabungkan eksperimen translokasi dengan uji pilihan kafetaria dan pengukuran sifat, yang melibatkan 8 spesies orthoptera dan 35 spesies tanaman yang termasuk dalam klade Liliopsida atau Magnoliopsida untuk menangkap keragaman besar sifat fungsional. Tingkat herbivori dinilai dengan memaparkan komunitas orthoptera yang lebih tinggi dan lebih rendah ke komunitas tanaman dataran tinggi di dua lokasi yang berbeda secara iklim. Kami kemudian mengukur pola makan orthoptera dan mengeksplorasi hubungan antara preferensi makan dan sifat fungsional tanaman. Pendekatan ini memberikan pemahaman mekanistik tentang pergeseran tanaman inang yang didorong oleh iklim dengan menganalisis sifat fungsional dari sumber makanan tanaman dan herbivora mereka. Hal ini memungkinkan kami untuk menilai dampak potensial dari spesies dataran rendah yang terbentuk di ekosistem pegunungan.
2 BAHAN DAN METODE
2.1 Percobaan di kebun umum
Untuk mengeksplorasi bagaimana pergeseran lereng orthoptera yang didorong oleh iklim mempengaruhi komunitas tumbuhan, kami membuat dua kebun umum pada ketinggian 1500 dan 2150 m di atas permukaan laut. Lokasi dipilih untuk memastikan paparan sinar matahari, lereng, dan orientasi yang serupa (Tabel S1 ). Suhu rata-rata selama percobaan (Juli–September) bersesuaian dengan 17,1°C dan 16,0°C untuk kebun umum dengan elevasi rendah (1500 m) dan tinggi (2150 m), masing-masing (Gambar S1 ). Dengan demikian, kebun umum dengan elevasi rendah menggambarkan skenario pemanasan +1,1°C. Di setiap kebun umum, 21 kandang didirikan dengan komunitas tumbuhan serupa yang terdiri dari sembilan spesies alpine ( n = 4 Liliopsida dan n = 5 Magnoliopsida): Anthyllis vulneraria subsp. alpestris (Schult.) Asch. & Graebn., Carex sempervirens Vill., Festuca ovina subsp. laevigata Gaudin., Helictotrichon versicolor (Vill.) Pilg., Linaria alpina (L.) Mill., Plantago alpina L., Ranunculus alpestris L., Sesleria caerulea (L.) Ard. dan Trifolium alpinum L. Semua spesies ini melimpah dan tidak terancam di area studi dan Swiss (Bornand et al., 2016 ; Lauber et al., 2018 ). Tanaman digali di sekitar lokasi dataran tinggi. Individu dipilih untuk menunjukkan ukuran yang sama dan digali dengan hati-hati untuk mencegah kerusakan akar. Semua daun yang menunjukkan tanda herbivori disingkirkan sebelum menambahkan setiap tanaman ke dalam kandang berukuran 50 × 50 × 90 cm yang ditutup dengan jaring nilon bermata halus dan kotak kayu berukuran 50 × 50 × 15 cm di bagian bawah (Gambar S2 ). Di setiap kandang, satu individu per spesies ( n = 9 tanaman per kandang) ditanam bersama dalam rancangan acak di tanah standar dengan 1/3 vermikulit dan 2/3 tanah pot (Terreau Suisse, Ricoter, Aaberg, Swiss). Tanaman dibiarkan beraklimatisasi selama 10 hingga 13 hari. Selama periode ini, tanaman disiram secara teratur, dan tanaman yang mati disingkirkan dan diganti.
Selanjutnya kami menambahkan sembilan individu orthoptera per kandang, menurut tiga perlakuan ( n = 7 ulangan per perlakuan di setiap kebun umum): (i) hanya orthoptera dataran rendah, (ii) hanya orthoptera dataran tinggi, dan (iii) campuran spesies orthoptera dataran tinggi dan rendah. Perlakuan ketinggian campuran diterapkan untuk mensimulasikan skenario di mana orthoptera dataran rendah tiba di tempat-tempat di mana orthoptera dataran tinggi tidak dapat bergeser ke atas (misalnya puncak gunung, kurangnya tanah dan padang rumput) dan dengan demikian akan menghadapi invasi spesies baru (McCain & Garfinkel, 2021 ). Individu orthoptera dikumpulkan secara acak di lapangan dan diidentifikasi hingga tingkat spesies sebelum ditambahkan ke kandang. Untuk perlakuan ketinggian campuran, kami secara acak mengambil empat hingga lima individu per ketinggian. Kepadatan sembilan individu serangga per kandang diatur untuk meniru kondisi realistis di area studi (Badenhausser, 2012 ; Deraison et al., 2015 ). Selanjutnya kami membiarkan percobaan berjalan selama sekitar 35 hari, setelah itu serangga dikeluarkan dari kandang dan dibekukan untuk analisis lebih lanjut (lihat di bawah). Pada akhir percobaan, herbivori diperkirakan secara visual pada setiap tanaman menggunakan skala tujuh tingkat: 0 = <1% (tidak ada herbivori yang terlihat), 1 = 1%–5%, 2 = 5%–13%, 3 = 13%–25%, 4 = 25%–50%, 5 = 50%–75% dan 6 = 75%–100%. Setelah itu, tanaman dikeringkan pada suhu 40°C selama 6 hari dan biomassa di atas tanah diukur (skala Sartorius, ±0,1 mg, Göttingen, Jerman). Kami kemudian memperkirakan biomassa kering yang dimakan per tanaman menggunakan nilai median kelas herbivori (yaitu 0,5%, 2,5%, 9%, 19%, 37,5%, 62,5% dan 87,5%) berdasarkan biomassa kering yang diukur.
Untuk percobaan kebun umum ini, kami memilih tujuh spesies orthopteran yang tersebar luas (Blumer & Diemer, 1996 ) dari subordo Caelifera, yang tidak terancam (Monnerat et al., 2007 ) dan yang terdapat pada ketinggian yang berbeda (Gambar S3A ). Lebih jauh lagi, lima dari spesies ini sudah berpindah ke ketinggian yang lebih tinggi di Swiss (Gambar S3B ). Individu-individu ditangkap dengan jaring kupu-kupu atau dengan kotak plastik di sekitar kebun umum, diidentifikasi dan dibagi menjadi dua kategori tergantung pada titik tengah distribusi ketinggian mereka dan ketinggian yang mereka sukai di area studi kami. Individu-individu yang termasuk spesies yang hidup di dataran rendah ( Euthystira brachyptera (Ocskay, 1826), Chorthippus parallelus (Zetterstedt, 1821), Stauroderus scalaris (Fischer von Waldheim, 1846), dan Omocestus viridulus (Linnaeus, 1758)) dimasukkan ke dalam kelompok dataran rendah, sementara individu-individu yang termasuk spesies yang hidup di dataran tinggi ( Gomphocerus sibiricus (Linnaeus, 1767), Podisma pedestris (Linnaeus, 1758), dan Miramella alpina (Kollar, 1833)) dimasukkan ke dalam kelompok dataran tinggi (Gambar S3A ).
Otorisasi terkait penangkapan serangga dan penggunaan lokasi penelitian diberikan oleh kanton Vaud (nomor otorisasi: 2364) dan kotamadya Lavey-Morcles (nomor otorisasi: 3.33.01). Persetujuan etika dari komite etika hewan tidak diperlukan untuk bekerja dengan serangga asli.
2.2 Uji pilihan kafetaria
Untuk mengeksplorasi preferensi makan spesies orthoptera yang dijadikan sampel dan memperkirakan jenis makanan mereka, kami melakukan uji pilihan kafetaria dengan memaparkan masing-masing spesies serangga ke 16 spesies tanaman yang tumbuh di dataran rendah atau kerabat terdekatnya dari dataran tinggi. Untuk setiap dataran tinggi, n = 13 dan n = 3 spesies Magnoliopsida dan Liliopsida, masing-masing, diuji. Liliopsida diwakili oleh anggota famili Cyperaceae dan Poaceae, sementara Magnoliopsida diwakili oleh anggota famili Ranunculaceae, Hypericaceae, Fabaceae, Rosaceae, Geraniaceae, Cistaceae, Lamiaceae, Plantaginaceae dan Asteraceae (rincian lebih lanjut tentang spesies tanaman dapat ditemukan di Informasi Pendukung , Tabel S2 ). Kami menggunakan tujuh spesies orthoptera seperti yang dijelaskan di atas, serta spesies tambahan yang mewakili sub-ordo Ensifera ( Metrioptera saussuriana ; Frey-Gessner, 1872), yang kemudian kami temukan dalam jumlah besar di dekat lokasi percobaan di dataran tinggi (Tabel S1 ). Serangga dikumpulkan pada tahap dewasa antara Agustus dan September 2017. Karena orthoptera mampu mengoptimalkan asupan makanannya melalui pembiasaan (Dukas & Bernays, 2000 ), semua individu yang dikumpulkan dipindahkan selama 2 hingga 3 hari ke dalam kandang dalam kondisi laboratorium yang berisi semua 32 spesies tanaman percobaan (Tabel S2 ). Tanaman diambil dari lokasi percobaan paling lama 3 hari sebelum dimulainya pengujian. Tanaman digali dengan hati-hati beserta akarnya, dimasukkan ke dalam kantong plastik yang lebih lembap dan disimpan pada suhu 4°C di tempat gelap. Daun yang paling kuat dipotong dan ditempatkan dalam kotak dengan tabung Eppendorf yang diisi dengan air di bagian bawah batang untuk menjaga turgor air maksimum di daun. Daun-daun ini kemudian ditempatkan secara acak di dalam baki plastik transparan 30 × 30 cm dalam lingkaran, berjarak sama satu sama lain dengan tangkai daun di dalam (Gambar S4 ). Kami memilih untuk menyajikan daun utuh, bukan fragmen daun dengan ukuran standar untuk menghindari pengeringan yang cepat (Ibanez et al., 2013 ). Setelah periode habituasi, serangga dibiarkan kelaparan selama satu malam dan kemudian dimasukkan secara individual ke dalam kotak selama 5 jam. Pada akhir pengujian makan, semua serangga dikumpulkan dan dibekukan untuk pengukuran sifat (lihat di bawah), dan kerusakan herbivora dinilai seperti dijelaskan di atas. Percobaan ini direplikasi antara n = 3–9 per spesies orthopteran untuk setiap komunitas tumbuhan (lihat Tabel S3 ).
2.3 Karakterisasi makanan orthoptera berdasarkan sifat fungsionalnya
Untuk mengkaji ruang fungsional tanaman yang memengaruhi preferensi diet orthopteran, kami mengukur serangkaian sifat fungsional yang diketahui berkorelasi dengan herbivori dan diet serangga (Ibanez et al., 2013 ): luas daun spesifik (SLA), kandungan bahan kering daun (LDMC), ketangguhan daun, konsentrasi silika (Si), dan rasio antara karbon dan nitrogen (rasio C:N). SLA dan LDMC diukur menurut metode yang dijelaskan oleh Perez-Harguindeguy et al. ( 2013 ), dan konsentrasi Si mengikuti Hallmark et al. ( 1982 ). Untuk rasio C:N, 1–2 mg bahan daun kering digiling menjadi bubuk untuk kuantifikasi total C dan N menggunakan penganalisa unsur (Flash 2000, CHN-O Analyzer; Thermo Scientific, Waltham, AS). Ciri-ciri ini diukur pada tanaman yang digunakan untuk percobaan kafetaria, sementara ketangguhan daun diperoleh dari kumpulan data sebelumnya dari lokasi geografis yang sama (Descombes, Marchon, dkk., 2017 ).
Untuk mengkarakterisasi riwayat hidup dan sifat fungsional terkait makan pada spesies orthoptera yang berbeda, kami mengukur: kekuatan tajam (IS), rasio antara panjang femur dan tibia (rasio femur/tibia), massa tubuh kering (DW) dan rasio antara karbon dan nitrogen (rasio C:N). Sebelum pengukuran sifat, serangga dikeringkan selama 7 hari pada suhu 40°C dan ditimbang untuk mendapatkan DW. Pengukuran untuk IS dan rasio femur/tibia dilakukan dengan menggunakan gambar yang diambil menggunakan mikroskop stereo (Leica M205 C; Wetzlar, Jerman), yang diproses dengan perangkat lunak ImageJ (Schneider et al., 2012 ). IS diukur mengikuti Ibanez et al. ( 2013 ) dan rasio femur/tibia diukur sebagai panjang tibia posterior dibagi dengan panjang femur posterior. Untuk pengukuran rasio C:N, individu digiling menjadi bubuk untuk kuantifikasi total C dan N menggunakan penganalisis unsur (Flash 2000, CHN-O Analyzer; Thermo Scientific, Waltham, AS). Pengukuran dilakukan pada individu yang digunakan selama kafetaria dan percobaan translokasi, dan untuk meningkatkan jumlah replikasi, kami menambahkan n = 11 individu O. viridulus yang berasal dari Chaumont (Neuchâtel, Swiss, 47° 03′ LU, 6° 59′ BT). Untuk setiap spesies, n = 10–23 replikasi digunakan (lihat Tabel S4 ).
2.4 Analisis statistik
2.4.1 Percobaan di kebun umum
Untuk menguji apakah perlakuan orthopteran, elevasi kebun umum dan klade tanaman (baik Liliopsida atau Magnoliopsida) memengaruhi jumlah biomassa kering yang dimakan (yaitu herbivori), kami memasang model efek campuran linier menggunakan paket ‘lmerTest’ (Kuznetsova et al., 2017 ). Elevasi, klade, perlakuan dan interaksinya diimplementasikan sebagai variabel penjelas. Kandang tempat percobaan dilakukan dan spesies tanaman yang digunakan diimplementasikan sebagai faktor acak. Signifikansi efek utama dan interaksinya diperkirakan dengan melakukan uji Chi-kuadrat menggunakan analisis varians (ANOVA) dengan paket ‘car’ (Fox & Weisberg, 2019 ). Untuk mengeksplorasi perbedaan berpasangan antara klade, perlakuan dan interaksinya, analisis post hoc dilakukan menggunakan paket ‘emmeans’, dengan penyesuaian Tukey (Lenth, 2022 ). Kami juga menguji efek perlakuan dan elevasi terhadap jumlah herbivori yang diamati untuk setiap spesies tanaman, dengan memasang model linier untuk setiap spesies secara terpisah menggunakan paket ‘stats’ (R Core Team, 2021 ). Elevasi, perlakuan, dan interaksinya diterapkan sebagai variabel penjelas, sedangkan biomassa kering yang dimakan diterapkan sebagai variabel respons. Efek elevasi, perlakuan, dan interaksinya diperkirakan dengan melakukan uji – F menggunakan analisis varians (ANOVA) dengan paket ‘car’ (Fox & Weisberg, 2019 ). Terakhir, analisis post hoc dilakukan (perbandingan berpasangan dengan penyesuaian Tukey) menggunakan paket ‘emmeans’ (Lenth, 2022 ).
2.4.2 Uji pilihan kafetaria
Bahasa Indonesia: Untuk menguji apakah pola makan orthoptera berubah ketika dihadapkan dengan komunitas tumbuhan dataran tinggi dibandingkan dengan dataran rendah, pertama-tama kami menghitung matriks jarak Euklides untuk setiap komunitas berdasarkan intensitas kerusakan herbivori yang dibuat pada setiap spesies tumbuhan oleh setiap spesies orthoptera. Kami kemudian melakukan uji korelasi Mantel Pearson dengan 10.000 permutasi antara matriks jarak Euklides kerusakan herbivori dengan paket ‘vegan’ (Oksanen et al., 2020 ). Kami juga menguji apakah spesies orthoptera lebih suka memakan Liliopsida atau Magnoliopsida dengan memasang model campuran linier umum (GLM) dengan distribusi kuasi-Poisson menggunakan paket ‘stats’. Kami menerapkan nilai median yang ditransformasikan akar kuadrat dari kelas herbivori sebagai variabel respons. Spesies orthoptera dan klade tumbuhan diterapkan sebagai variabel penjelas. Efek spesies dan klade diperkirakan dengan melakukan uji F menggunakan analisis varians (ANOVA) dengan paket ‘car’ (Fox & Weisberg, 2019 ). Terakhir, analisis post hoc (perbandingan berpasangan dengan penyesuaian Tukey) dilakukan menggunakan paket ‘emmeans’ (Lenth, 2022 ).
2.4.3 Ciri-ciri fungsional tumbuhan dan serangga
Perbedaan dalam sifat fungsional spesies tanaman dan serangga dieksplorasi dengan melakukan analisis komponen utama (PCA), termasuk sifat-sifat yang dijelaskan di atas sebagai variabel (Gambar S5 ) menggunakan paket ‘factoextra’ (Kassambara & Mundt, 2020 ) dan ‘FactoMineR’ (Lê et al., 2008 ). Ruang diet fungsional rata-rata yang dimakan oleh orthoptera diperkirakan sebagai titik berat ruang multidimensi yang dihasilkan oleh korelasi antara hasil uji pilihan kafetaria dan koordinat setiap spesies tanaman yang diperoleh dengan PCA. Perbedaan dalam ciri fungsional pola makan spesies Orthoptera (ciri tumbuhan) dan dalam ciri morfologi spesies Orthoptera menurut preferensi makannya (baik pemakan Magnoliopsida maupun Liliopsida) dieksplorasi dengan melakukan analisis ANOVA multivariat permutasi (PERMANOVA) dengan jarak Euclidian pada model PCA menggunakan paket vegan (Oksanen et al., 2020 ). Akhirnya, untuk menguji apakah ciri Orthoptera berkorelasi dengan ciri pola makannya, kami memasang model linier menggunakan paket ‘stats’ (Tim Inti R, 2021 ). Kami menerapkan sumbu PCA pertama dari ciri Orthoptera sebagai variabel respons dan sumbu PCA pertama dari sentroid pola makan Orthoptera sebagai variabel penjelas. Pengaruh diet terhadap sifat-sifat orthopteran diperkirakan dengan melakukan uji-F menggunakan analisis varians (ANOVA) dengan paket ‘mobil’ (Fox & Weisberg, 2019 ).
Semua analisis statistik dilakukan menggunakan R (Tim Inti R, 2021 ) dan Rstudio (Tim RStudio, 2021 ).
2.5 Pernyataan replikasi
| Percobaan | Skala inferensi | Skala penerapan faktor minat | Jumlah replikasi pada skala yang sesuai |
|---|---|---|---|
| Percobaan kebun umum di lapangan | Komunitas tumbuhan | Kurungan | Tiga komunitas orthoptera diperlakukan di dua kebun umum dengan n = 7 kandang per perlakuan dan kebun umum |
| Uji pilihan kafetaria | Jenis | Cawan petri dengan komunitas tanaman yang berbeda | Delapan spesies orthoptera diuji dengan komunitas tumbuhan dataran tinggi dan rendah, dengan n = 3–9 replikasi per spesies orthoptera dan ketinggian |
| Karakterisasi pola makan Orthoptera berdasarkan sifat fungsionalnya | Jenis | Individu | Delapan spesies orthopteran diuji dengan n = 10–23 individu untuk pengukuran sifat per spesies |
3 HASIL
3.1 Percobaan di kebun umum
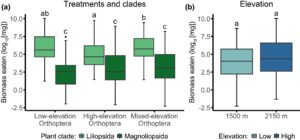
Kami tidak menemukan efek elevasi pada keseluruhan biomassa kering yang dimakan di seluruh komunitas tanaman yang diuji (Gambar 1B ; Chi 2 = 2,56, Df = 1, p = 0,109). Secara keseluruhan, Liliopsida 4,6 kali lebih banyak dimakan daripada Magnoliopsida (Gambar 1A ; Chi 2 = 13,08, Df = 1, p < 0,001). Namun, 35% lebih banyak biomassa Liliopsida dimakan ketika tanaman dihadapkan dengan komunitas orthopteran campuran dibandingkan dengan komunitas orthopteran di elevasi yang lebih tinggi (Perlakuan: Klade, Chi 2 = 10,48, Df = 2, p = 0,005, perbedaan berpasangan dengan koreksi Tukey: p = 0,044). Pada tingkat spesies (lihat Gambar S6 dan Tabel S6 ), dua spesies menghadapi tingkat herbivori yang lebih tinggi ketika dihadapkan dengan komunitas orthopteran campuran. H. versicolor (Liliopsida) menghadapi peningkatan herbivori sebesar 79% ketika terpapar komunitas orthopteran campuran dibandingkan dengan komunitas orthopteran di dataran tinggi ( F 36,2 = 6,12, p = 0,005, perbedaan berpasangan dengan koreksi Tukey: p = 0,005). Di dataran tinggi, R. alpestris (Magnoliopsida) menghadapi peningkatan sebesar 84% dan 93% ketika terpapar komunitas orthopteran campuran dibandingkan dengan komunitas orthopteran di dataran rendah dan tinggi, masing-masing (Perlakuan: Ketinggian, F 36,2 = 4,81, p = 0,014, perbedaan berpasangan dengan koreksi Tukey: p = 0,042 dan 0,012, masing-masing, lihat Gambar S7 ).
3.2 Uji pilihan kafetaria
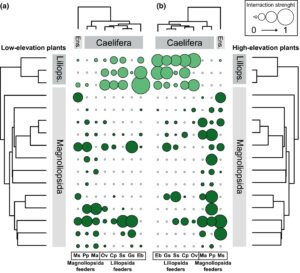
Kami menemukan bahwa spesies orthoptera yang diteliti dapat dipisahkan menjadi dua tipe makan berdasarkan preferensi mereka (Gambar 2 dan lihat Tabel S7 ): baik ‘pemakan Liliopsida’ ( O. viridulus , S. scalaris , E. brachyptera , P. parallelus dan G. sibiricus ) atau ‘pemakan Magnoliopsida’ ( M. saussuriana , M. alpina dan P. pedestris ). Selain itu, kami mengamati bahwa spesies orthoptera konservatif dalam penggunaan inangnya; ketika dihadapkan dengan komunitas tanaman inang baru, mereka selalu cenderung memakan kerabat terdekat dari tanaman inang standar mereka (Gambar 2 , statistik Mantel r = 0,46, p = 0,034).
3.3 Ciri-ciri fungsional tumbuhan dan serangga
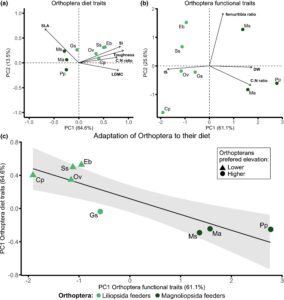
Berdasarkan analisis komponen utama sifat-sifat tanaman, kami menemukan bahwa pengumpan Magnoliopsida cenderung memakan tanaman yang menunjukkan nilai SLA tinggi, sedangkan pengumpan Liliopsida, sebaliknya, cenderung memakan tanaman dengan LDMC tinggi, kandungan Si tinggi, rasio C:N tinggi dan daun keras (Gambar 3a , F 1,6 = 9,29, p = 0,018). Selain itu, pengumpan Liliopsida dan Magnoliopsida juga memiliki sifat fungsional yang berbeda (Gambar 3b , F 1,6 = 7,30, p = 0,017). Pengumpan Liliopsida cenderung memiliki kekuatan tajam yang lebih tinggi dan lebih kecil daripada pengumpan Magnoliopsida. Ketika kita menggabungkan ciri-ciri fungsional spesies orthoptera dan ciri-ciri fungsional makanan mereka, kami mengamati bahwa kombinasi ciri-ciri khusus orthoptera terkait secara signifikan dengan ciri-ciri tanaman tertentu (Gambar 3c , F 1,6 = 20,08, p = 0,004), di mana spesies orthoptera dengan IS yang lebih tinggi dan berat kering yang lebih rendah lebih suka memakan daun yang lebih keras. Secara keseluruhan, orthoptera dengan IS yang lebih rendah dan berat kering yang lebih besar lebih suka memakan daun yang lebih lunak.
4 DISKUSI
Memahami bagaimana perubahan iklim membentuk kembali interaksi biotik sangat penting untuk mengantisipasi pergeseran dalam ekosistem pegunungan (Alexander et al., 2015 ; Block et al., 2022 ; Kerner et al., 2023 ). Dalam studi ini, kami menunjukkan bahwa tumbuhan dataran tinggi dari klad Liliopsida mengalami peningkatan herbivori ketika terpapar komunitas orthoptera dataran campuran dibandingkan dengan orthoptera dataran tinggi saja. Demikian pula, pada tingkat spesies, biomassa tumbuhan yang lebih besar dikonsumsi ketika H. versicolor dan R. alpestris berhadapan dengan orthoptera dataran tinggi dan rendah. Model realistis pergeseran lereng ke atas orthoptera di bawah perubahan iklim meramalkan bahwa kekayaan spesies orthoptera akan meningkat pada ketinggian tinggi (Descombes et al., 2020 ), yang menunjukkan bahwa perlakuan orthoptera pada ketinggian campuran dapat mensimulasikan perubahan realistis dalam interaksi tumbuhan-orthoptera ketika spesies pada ketinggian lebih tinggi dibatasi dalam pergeseran ke atas mereka. Dengan demikian, hasil kami menunjukkan bahwa jika orthoptera pada ketinggian lebih rendah dapat tumbuh subur di ketinggian tinggi, mereka kemungkinan akan meningkatkan tekanan herbivori pada beberapa tanaman pegunungan. Yang penting, perlakuan pemanasan (yaitu translokasi pada ketinggian lebih rendah) tidak memengaruhi konsumsi biomassa untuk sebagian besar spesies tanaman, yang menyoroti bahwa interaksi biotik mungkin memainkan peran yang lebih besar daripada pemanasan saja dalam menentukan persistensi spesies dalam ekosistem pegunungan (Bovay et al., 2024 ; Nomoto et al., 2024 ). Taksonomi tanaman juga tampaknya memengaruhi ketahanan spesies terhadap interaksi biotik baru. Dalam percobaan translokasi kami, tanaman Liliopsida lebih banyak dikonsumsi daripada Magnoliopsida, khususnya di hadapan komunitas orthoptera dataran campuran. Ini mengonfirmasi bahwa, meskipun bersifat generalis, orthoptera dapat menunjukkan preferensi makanan (Descombes et al., 2020 ; König et al., 2022 ; Pitteloud et al., 2021 ), karena orthoptera dataran rendah tampaknya lebih suka memakan Liliopsida. Selain itu, perbedaan preferensi makanan antara orthoptera dataran rendah dan tinggi menunjukkan komplementaritas relung ketika komunitas mereka tumpang tindih (Barry et al., 2019 ). Namun, karena kami hanya menguji sebagian kecil tanaman yang mewakili famili yang paling melimpah secara alami yang terdapat di area studi kami, maka mungkin saja temuan kami tidak berlaku untuk famili lain yang menunjukkan sifat fungsional yang berbeda, khususnya untuk Liliopsida non-graminiform.
Dengan menggunakan uji kafetaria, kami mengungkap bahwa orthoptera lebih suka memakan tanaman dataran tinggi yang terkait erat dengan makanan aslinya, yang mengonfirmasi bahwa spesies ini menunjukkan konservatisme pola makan ketika berpindah ke dataran tinggi, konsisten dengan penelitian sebelumnya (Descombes et al., 2020 ; Kuřavová et al., 2017 ). Temuan tersebut mengindikasikan bahwa orthoptera yang bermigrasi ke dataran tinggi mungkin menerima tanaman dataran tinggi dalam makanannya dan dengan demikian dapat menentukan di mana tanaman yang terkait dengan inangnya saat ini berada. Meskipun demikian, kami juga mengungkapkan bahwa spesies yang diuji dapat dibagi menjadi dua kelompok: ‘pemakan Liliopsida’ atau ‘pemakan Magnoliopsida’. Pembagian seperti itu juga kongruen dengan filogeni spesies orthopteran, karena semua pemakan Liliopsida termasuk dalam subfamili Gomphocerinae, sedangkan pemakan Magnoliopsida termasuk dalam subfamili lain (König et al., 2022 ). Namun, yang penting, klasifikasi ini mengungkapkan pemisahan fungsional berdasarkan ketinggian, di mana spesies dataran rendah sebagian besar memakan Liliopsida, sedangkan spesies dataran tinggi (kecuali G. sibiricus ) lebih menyukai Magnoliopsida. Pemisahan fungsional ini kemungkinan menjelaskan hasil percobaan translokasi, di mana Liliopsida secara umum, dan H. versicolor secara khusus, mengalami herbivori yang lebih tinggi ketika terpapar komunitas orthopteran campuran dibandingkan dengan komunitas dataran tinggi. Sebaliknya, R. alpestris lebih banyak dikonsumsi dalam perlakuan campuran dibandingkan dengan komunitas orthoptera dataran rendah, karena orthoptera dataran tinggi dalam perlakuan campuran terutama memakan Magnoliopsida. Perbedaan pola makan antara spesies orthoptera dataran tinggi dan rendah ini mungkin mencerminkan ketersediaan dan kualitas nutrisi tanaman di berbagai ketinggian. Di dataran tinggi, Magnoliopsida lebih melimpah (Descombes, Vittoz, et al., 2017 ) dan menyediakan daun yang lebih bergizi (Körner, 1989 ), yang mungkin menguntungkan bagi orthoptera di ekosistem pegunungan dengan musim tanam yang pendek. Sebaliknya, orthoptera dataran rendah mungkin lebih menyukai Liliopsida karena ketersediaannya sepanjang musim tanam, karena Magnoliopsida dapat mengering selama musim panas. Temuan ini juga dapat dihubungkan dengan hipotesis luas relung ketinggian (Rasmann, Alvarez, et al., 2014 ), karena pada ketinggian tinggi, menjadi pemakan Magnoliopsida dapat memungkinkan Orthoptera untuk memakan lebih banyak spesies tanaman dibandingkan dengan menjadi pemakan Liliopsida.
Lebih jauh lagi, analisis sifat fungsional mengungkapkan bahwa sifat-sifat tanaman yang dimakan oleh pemakan Liliopsida atau Magnoliopsida berbeda secara signifikan, dengan sifat-sifat terkait diet berkorelasi terutama dengan ukuran orthoptera (berat kering) dan kekuatan tajam. Dengan demikian, kita dapat menyimpulkan bahwa orthoptera yang memiliki nilai sifat yang berbeda memakan klade tanaman yang berbeda, yang ditentukan oleh kemampuan mereka untuk memakannya. Misalnya, kami mengungkapkan bahwa pemakan Liliopsida memiliki kekuatan tajam yang lebih besar daripada pemakan Magnoliopsida, yang memungkinkan pemakan Liliopsida untuk memakan tanaman yang lebih keras, seperti yang sebelumnya ditunjukkan oleh Ibanez et al. ( 2013 ). Demikian pula, mandibula yang lebih besar telah terbukti memfasilitasi konsumsi daun yang keras pada ulat Lepidoptera (Prasannakumar & Kodandaramaiah, 2024 ) dan bentuk mandibula berbeda di antara kelompok orthoptera yang memiliki preferensi makan yang berbeda (Stockey et al., 2022 ). Dengan demikian, kita dapat menyimpulkan bahwa sementara orthoptera dapat memakan berbagai taksa, mereka masih menunjukkan beberapa tingkat spesialisasi dalam hal sifat tanaman (Forister et al., 2012 ), yang secara efektif memisahkan pemakan Liliopsida dan Magnoliopsida. Spesialisasi ini terutama didorong oleh kekuatan tajam dan ukuran orthoptera yang memungkinkan mereka memakan daun dengan tingkat ketangguhan yang berbeda dan dapat memberikan keuntungan selama pergeseran lereng dengan meminimalkan persaingan dengan spesies orthoptera alpine. Memang, spesies dataran tinggi yang dipelajari di sini (kecuali G. sibiricus ) menunjukkan pola makan dan sifat fungsional yang berbeda dibandingkan dengan spesies dataran rendah, yang menunjukkan bahwa relung dataran tinggi masih kurang dimanfaatkan. Akibatnya, orthoptera yang menggeser lereng ke dataran rendah dapat menjajah padang rumput alpine dengan persaingan minimal (McClenaghan et al., 2015 ). Pada gilirannya, karena preferensi herbivora dapat menentukan komposisi spesies suatu ekosistem (Kempel et al., 2015 ), pergeseran ke atas pengumpan Liliopsida di dataran rendah ini dapat membentuk kembali struktur komunitas dan keanekaragaman hayati padang rumput Alpen (Descombes et al., 2020 ).
5 KESIMPULAN
Dengan menyelidiki jumlah biomassa tanaman yang dimakan dalam berbagai iklim dan skenario pergeseran lereng, preferensi orthoptera, dan sifat fungsionalnya, kami telah mengambil pendekatan multidimensi untuk mengatasi masalah biologis yang mendesak. Karena serangga dengan cepat berpindah ke ketinggian yang lebih tinggi, mereka semakin menjadi ancaman bagi spesies dan ekosistem pegunungan (Bässler et al., 2013 ). Di sini, kami mengungkapkan untuk pertama kalinya diferensiasi sifat berbasis ketinggian pada orthoptera, dengan orthoptera dataran rendah memiliki sifat yang mendorong preferensi mereka terhadap Liliopsida sementara orthoptera dataran tinggi memiliki sifat yang mendorong preferensi mereka terhadap Magnoliopsida. Kami selanjutnya mengeksplorasi bagaimana perbedaan berbasis ketinggian ini dapat memengaruhi pergeseran ke atas orthoptera dataran rendah (Vitasse et al., 2021 ), yang menyebabkan mereka berpotensi berhasil menginvasi padang rumput pegunungan dengan koeksistensi yang stabil dengan komunitas herbivora dataran tinggi saat ini karena komplementaritas relung, yang akan menjadi tantangan khusus bagi spesies tanaman dataran tinggi. Secara khusus, jika mereka berhasil menyerang padang rumput Alpen, kami berharap mereka memakan lebih banyak spesies Liliopsida, seperti H. versicolor . Klade tumbuhan ini dengan demikian mungkin mengalami tekanan herbivori yang lebih tinggi, yang memaksa spesies untuk bergeser lebih jauh ke atas lereng, beradaptasi dengan perubahan tersebut atau menghadapi kepunahan (Berg et al., 2010 ). Dengan mensimulasikan skenario perubahan iklim di masa depan dan mengukur sifat-sifat yang terkait dengan tanaman dan pola makan, kami dapat membuat prediksi yang koheren tentang bagaimana ekosistem Alpen dapat merespons efek langsung dan tidak langsung dari perubahan iklim. Namun, penelitian di masa depan harus menggabungkan interaksi biotik yang lebih kompleks untuk mengevaluasi potensi koeksistensi yang stabil antara herbivora dataran tinggi dan rendah (Chesson & Kuang, 2008 ). Lebih banyak faktor yang mengkondisikan keberhasilan pembentukan serangga dataran rendah di dataran tinggi juga harus diselidiki, seperti efek tanaman inang baru pada instar awal, yang mungkin lebih peka terhadap perubahan makanan (Bovay et al., 2024 ; Radchuk et al., 2013 ). Akhirnya, studi jangka panjang juga dianjurkan untuk menilai kapasitas tanaman di dataran tinggi untuk beradaptasi terhadap peningkatan herbivori dan untuk mengukur risiko kepunahan bagi spesies atau klade yang rentan.


