
Abstrak
- Perluasan semak daun ke tundra Arktik yang didominasi tumbuhan graminoid diperkirakan akan mengubah dekomposisi serasah dengan mengubah kualitas serasah dan kondisi abiotik dan biotik setempat. Namun, masih belum jelas bagaimana perluasan semak akan memengaruhi dekomposisi serasah pada skala regional, di mana iklim makro diperkirakan menjadi pengatur dekomposisi yang dominan.
- Untuk menentukan pengaruh relatif dari iklim makro dan kontrol lokal pada pola regional kehilangan massa serasah dan pelepasan nitrogen, kami melakukan dua eksperimen dekomposisi serasah hierarkis di seluruh skala spasial. Kami menguraikan serasah daun dan akar dari sejenis rumput yang menonjol ( Eriophorum vaginatum ) dan tiga genus semak peluruh ( Betula , Alnus dan Salix ) selama 1 tahun dalam plot yang direplikasi di lima lokasi yang mencakup gradien lintang 160 km di Alaska utara.
- Dengan menggunakan serasah Eriophorum sebagai substrat, kami menemukan bahwa iklim makro adalah pengatur utama kehilangan massa tetapi memiliki efek yang berlawanan pada serasah daun dan akar. Saat suhu musim panas meningkat sepanjang gradien lintang (11,9 hingga 13,9°C), kehilangan massa serasah daun meningkat sebesar 20% sedangkan kehilangan massa serasah akar menurun sebesar 33%. Pelepasan nitrogen daun juga meningkat seiring dengan suhu musim panas. Sebaliknya, pelepasan nitrogen akar dikendalikan oleh jenis vegetasi lingkungan dekomposisi. Dengan menggunakan serasah semak yang berbeda sebagai substrat, kami menemukan bahwa kualitas serasah dan interaksinya dengan iklim mikro tanah dan iklim makro mengendalikan dekomposisi. Secara keseluruhan, serasah akar semak terurai lebih cepat daripada serasah akar Eriophorum , kehilangan 53% lebih banyak massa dan 190% lebih banyak nitrogen di semua lokasi dan lingkungan dekomposisi. Namun, untuk serasah daun, polanya bervariasi menurut genus serasah, dengan Salix kehilangan lebih banyak massa dan Betula dan Alnus kehilangan lebih sedikit massa daripada Eriophorum .
- Temuan kami menunjukkan bahwa perluasan semak di Arktik dapat mengatur dekomposisi serasah daun dan akar pada skala regional melalui dampaknya pada kontrol lokal, terutama kualitas serasah. Peningkatan tutupan semak yang berkelanjutan kemungkinan akan mempercepat pergantian kumpulan karbon dan nitrogen serasah akar di ekosistem tundra. Oleh karena itu, memasukkan proses terkait semak dalam model sistem Bumi akan meningkatkan kemampuan kita untuk memprediksi dekomposisi serasah skala regional dan dampaknya pada siklus karbon dan nutrisi di Arktik yang menghangat.
Baca Ringkasan Bahasa Sederhana gratis untuk artikel ini di blog Jurnal.
1. PENDAHULUAN
Wilayah Arktik memanas empat kali lebih cepat daripada wilayah non-Arktik (Rantanen et al., 2022 ). Sebagai respons, semak peluruh daun meningkat tutupannya dan jumlahnya di banyak bagian tundra yang didominasi graminoid (Sturm et al., 2001 ; Tape et al., 2006 ). Perluasan semak diperkirakan akan mengubah laju dekomposisi serasah, yang dapat sangat memengaruhi siklus karbon (C) dan nutrisi dan berpotensi menjadi umpan balik terhadap perubahan iklim (Mekonnen et al., 2021 ; Myers-Smith et al., 2011 ). Namun, kepentingan relatif perluasan semak dan pemanasan iklim sebagai pengendali fluks C dan N masih belum jelas.
Semak dapat memengaruhi dekomposisi serasah dengan mengubah komposisi dan kuantitas masukan serasah. Semak peluruh sering menghasilkan serasah daun dalam jumlah lebih banyak dengan rasio C:N lebih rendah daripada bentuk pertumbuhan tundra lainnya (Hobbie, 1996 ), sehingga menghasilkan masukan N biomassa lebih tinggi yang dapat merangsang aktivitas pengurai mikroba dan mempercepat dekomposisi serasah (Christiansen et al., 2018 ; DeMarco et al., 2014 ; McLaren et al., 2017 ). Selain itu, semak dapat memperkenalkan serasah akar baru (Chen et al., 2020 ; Träger et al., 2017 ). Meskipun lebih sedikit penelitian yang meneliti bagaimana masukan dan pergantian serasah akar semak berbeda dari bentuk kehidupan tundra lainnya, sebuah penelitian baru-baru ini menemukan kehilangan massa serasah akar yang lebih besar dari semak Betula nana yang berkembang dibandingkan dengan alang-alang dominan regional Eriophorum vaginatum selama satu musim tanam (Wang et al., 2017 ).
Semak belukar juga dapat memengaruhi dekomposisi serasah dengan mengubah lingkungan lokalnya. Semak belukar peluruh berkembang terutama menjadi tundra tussock, yang dicirikan oleh tajuk terbuka dengan paparan cuaca tinggi. Relatif terhadap tundra tussock, keberadaan tajuk semak belukar dapat meningkatkan suhu tanah musim dingin dan menurunkan suhu tanah musim panas dengan masing-masing meningkatkan akumulasi salju dan naungan, yang menghasilkan pemanasan tahunan bersih tanah di bawah tajuk semak belukar (Aguirre et al., 2021 ; Kropp et al., 2021 ). Vegetasi semak belukar yang lebih tinggi juga dapat meningkatkan kelembapan tanah dengan mengurangi evapotranspirasi melalui naungan (Eugster et al., 2000 ). Bersama-sama, perubahan iklim mikro ini dapat meningkatkan laju pengurai memecah serasah (Christiansen et al., 2018 ; Hicks Pries et al., 2013 ).
Selain itu, semak dapat mendorong perubahan biotik di lingkungan lokal. Misalnya, semak dapat berasosiasi dengan dan menyukai mikroba tanah tertentu, termasuk jamur ektomikoriza (Chen et al., 2020 ; Clemmensen et al., 2006 ), yang dapat mengakibatkan pergantian C yang lebih cepat di komunitas semak Arktik (Parker et al., 2021 ). Spesies semak yang secara simbiosis mengikat N 2 , seperti Alnus sp., dapat meningkatkan kumpulan N tanah lokal, yang selanjutnya mendorong dekomposisi di lingkungan sekitar semak (Mitchell & Ruess, 2009 ). Namun, efek kumulatif dari perubahan ini dapat bervariasi. Studi terbaru menunjukkan bahwa spesies serasah, iklim mikro tanah, dan iklim memengaruhi dekomposisi serasah daun dan akar secara berbeda, yang sering kali mengakibatkan peningkatan dekomposisi serasah daun dibandingkan dengan akar (Guo et al., 2021 ; McLaren et al., 2017 ). Ada pula bukti bahwa komponen kimia yang berbeda dari kualitas serasah (yaitu lignin: N, kualitas non-lignin C) mengatur dekomposisi serasah daun dan akar (Sun et al., 2018 ).
Pada akhirnya, iklim diharapkan untuk mengatur dekomposisi pada skala regional, dengan kualitas serasah menjadi lebih penting di mana iklim kurang membatasi aktivitas pengurai (Cornelissen et al., 2007 ; Swift et al., 1979 ). Meningkatnya suhu udara telah terbukti meningkatkan laju dekomposisi di tundra dan bioma dingin lainnya (Breeuwer et al., 2008 ; Xue et al., 2016 ), dengan kelembaban juga melakukan pengendalian (Aerts, 2006 ; Hicks Pries et al., 2013 ). Karena struktur dan aktivitas komunitas pengurai dapat bervariasi dengan iklim, namun (Christiansen et al., 2017 ), perubahan yang disebabkan oleh semak dalam kualitas serasah dan kondisi abiotik dan biotik lokal dari lingkungan dekomposisi dapat menghasilkan pola dekomposisi yang berbeda dalam lokasi yang memiliki iklim yang sama. Memahami potensi ketergantungan skala ini dalam pola dan kontrol dekomposisi serasah regional memiliki implikasi yang cukup besar untuk memperkirakan bagaimana perubahan iklim dan perluasan semak akan mengubah fluks C dan N karena model biogeokimia dan sistem Bumi sering dibangun di atas kerangka teoritis bahwa iklim adalah kontrol dominan dari pergantian serasah (Bonan et al., 2013 ). Namun sedikit penelitian yang secara bersamaan menyelidiki sumber variasi dalam dan antar-situs dalam dekomposisi serasah (Bradford et al., 2016 ), terutama di tundra. Oleh karena itu, masih belum diketahui bagaimana perluasan semak akan mempengaruhi pola regional dekomposisi serasah melalui interaksi kontrol lokal dan iklim.
Untuk mengatasi kesenjangan pengetahuan ini, kami melakukan eksperimen dekomposisi serasah hierarkis di sepanjang gradien lintang di Kutub Utara Alaska. Di masing-masing dari lima lokasi tundra, kami mengurai serasah daun dan akar dari tiga taksa semak daun dan Eriophorum vaginatum , spesies alang-alang dominan pembentuk rumpun, selama 1 tahun di lingkungan penguraian yang didominasi oleh semak atau rumpun untuk mengukur efek relatif dari iklim makro, kualitas serasah, iklim mikro tanah (suhu dan kelembapan) dan jenis vegetasi lingkungan penguraian pada pola penguraian. Kami berhipotesis bahwa serasah daun dan akar akan membusuk lebih cepat di lokasi yang lebih hangat dan di bawah semak peluruh, dengan bukti bahwa pembusukan serasah meningkat seiring suhu (Breeuwer et al., 2008 ; Cornelissen et al., 2007 ; Hobbie, 1996 ) dan lingkungan pembusukan semak lebih mendukung pembusukan serasah daripada lingkungan pembusukan alang-alang (Christiansen et al., 2018 ). Kami selanjutnya berhipotesis bahwa pembusukan serasah akan meningkat seiring kualitas serasah, dengan serasah semak membusuk lebih cepat daripada serasah alang-alang dan serasah daun membusuk lebih cepat daripada serasah akar di seluruh lingkungan pembusukan karena kualitasnya yang lebih tinggi (C:N lebih rendah) (Guo et al., 2021 ; Hobbie & Gough, 2004 ; McLaren et al., 2017 ). Akhirnya, kami berhipotesis bahwa pendorong iklim dari pembusukan akan lebih memengaruhi serasah daun daripada serasah akar karena peningkatan paparan lingkungan.
2 BAHAN DAN METODE
2.1 Lokasi penelitian dan desain eksperimen
Kelima situs tersebut membentang pada gradien lintang di Alaska utara (Gambar 1 ; lintang: 67.0° N–69.3° N; bujur: 148.7° W–150.3° W; nomor izin Departemen Sumber Daya Alam Alaska LAS 31709; nomor izin Biro Pengelolaan Lahan AS F 97344) yang mencakup berbagai kondisi iklim (Tabel S1 dalam Informasi Pendukung), yang berkovariasi dengan zona vegetasi yang luas (Walker et al., 2005 ). Situs paling utara berada di Lereng Utara Pegunungan Brooks dekat tepi selatan Dataran Pantai Arktik. Wilayah ini dicirikan oleh semak rendah dan alang-alang pembentuk tussock. Situs paling selatan berada di kaki bukit selatan Pegunungan Brooks dekat tepi utara ekoton hutan-tundra boreal. Wilayah ini dicirikan oleh pepohonan jarang (terutama Picea mariana ) diselingi dengan semak rendah dan alang-alang pembentuk rumpun. Seluruh area memiliki permafrost berkelanjutan (Jorgenson et al., 2008 ) dan tanah yang berasal dari till glasial (Munroe & Bockheim, 2001 ). Selain perbedaan dalam lintang dan kondisi bioklimat terkait, lokasi dipilih untuk meminimalkan perbedaan lingkungan. Dengan demikian, semua lokasi berada di tundra asam lembap, di lereng landai (2,5–10°) dekat jalur air, dan setidaknya 100 m dari jalan untuk meminimalkan dampak jalan pada karakteristik edafik (Fraterrigo et al., 2024 ). Vegetasi di lokasi didominasi oleh alang-alang Eriophorum vaginatum dan juga mencakup graminoid Carex bigelowii , semak peluruh dan hijau Vaccinium uliginosum , V . vitis-idaea , Betula nana , B. glandulosa, Alnus viridis ssp. Fruticosa, Salix pulchra, S. glauca, S. richardsonii, Rhododendron tomentosum , Empetrum nigrum , dan berbagai herba menahun, lumut, dan lumut kerak. Dalam penelitian sebelumnya yang dilakukan di lokasi-lokasi ini, produktivitas akar lebih tinggi dan cadangan C dan N tanah lebih besar di lokasi yang lebih hangat daripada di lokasi yang lebih dingin; selain itu, semak peluruh aktif berkembang di dua lokasi paling selatan (Chen et al., 2020 ).

Pada tahun 2017, kami membuat plot replikasi dalam empat tipe vegetasi di masing-masing dari lima lokasi. Tipe vegetasi tersebut terdiri dari teki pembentuk tussock E. vaginatum , spesies dasar tundra asam lembap dan tiga taksa semak peluruh yang pertumbuhan, distribusi, dan kelimpahan relatifnya meningkat di tundra Arktik (Myers-Smith et al., 2011 ): Alnus ( A. viridis ssp. Fruticosa ), Betula ( B. nana ) dan Salix ( S. glauca , S. pulchra atau S. richardsonii ). Ketiga taksa semak tersebut membentuk asosiasi simbiosis dengan jamur ektomikoriza, yang dapat menguraikan bahan organik dan membantu penyerapan nutrisi tanaman (Chen et al., 2020 ). Selain itu, asosiasi Alnus dengan bakteri Frankia pemfiksasi N 2 dapat meningkatkan produktivitas tanaman dengan memperkaya tanah Arktik yang kekurangan N (Mitchell & Ruess, 2009 ). Di setiap lokasi, kami memusatkan tujuh plot melingkar pada rumpun E. vaginatum dan sembilan plot melingkar pada individu yang tidak rusak dan matang secara reproduktif dari setiap taksa semak, secara kumulatif 170 plot di semua lokasi dan taksa ( N = 34 plot/lokasi). Plot melingkar berdiameter 1 m dan disusun dalam kelompok yang mencakup rentang kondisi lingkungan mikro di setiap lokasi (Gambar S1 ). Setiap kelompok terdiri dari tiga atau empat plot (<5 m terpisah), termasuk plot yang didominasi oleh masing-masing taksa semak dan plot yang didominasi oleh E. vaginatum . Pengaturan ini memastikan bahwa plot dari setiap jenis vegetasi hadir di seluruh rentang kondisi lingkungan mikro di dalam lokasi (Gambar S1 ).
2.1.1 Pengukuran lingkungan
Bahasa Indonesia: Kami memperoleh data rata-rata curah hujan dan suhu bulanan yang diperkecil (1 km) dari data Climate Research Unit (kumpulan data CRU TS4.05; Harris et al., 2020 ) yang tumpang tindih dengan durasi eksperimen dekomposisi (Juni 2017–Juli 2018). Data curah hujan dan suhu dirangkum menjadi tiga kategori: rata-rata tahunan, rata-rata musim panas (Juni–Agustus) dan rata-rata musim dingin (Desember–Februari). Untuk mengkarakterisasi iklim mikro tanah dalam setiap plot, kami mengukur suhu tanah dengan termometer digital (kedalaman 10 cm; akurasi 0,4°C; resolusi 0,1°C) dan kadar air volumetrik tanah dengan probe TDR (kedalaman 12 cm; probe Field Scout TDR 100, Spectrum Technologies). Pengukuran dilakukan di tiga titik terpisah dalam setiap plot antara pertengahan hingga akhir Juli tahun 2017 dan 2018. Jangka waktu yang singkat ini memungkinkan kami untuk mengunjungi semua lokasi sambil menghindari fluktuasi cuaca besar yang dapat membiaskan pengukuran. Semua pengukuran lintas tahun dan titik dalam plot (enam pengukuran per plot untuk kedua tahun) dirata-ratakan untuk menghasilkan satu nilai suhu tanah dan kelembapan tanah untuk setiap plot. Mengingat banyaknya plot dan lokasi yang terpencil, kami tidak dapat menempatkan dan memelihara perekam data iklim mikro di setiap plot untuk meningkatkan resolusi temporal pengukuran iklim mikro kami. Namun, dengan memperhatikan keterbatasan ini, kami mengamati variabilitas antartahunan yang relatif rendah dalam pengukuran kami (Tabel S2 ), yang menunjukkan bahwa pengukuran tersebut berhasil menggambarkan kondisi iklim mikro relatif, bukan kondisi intermiten yang disebabkan oleh cuaca.
2.2 Percobaan dekomposisi serasah
Kami menggunakan serasah E. vaginatum dan serasah semak yang dikumpulkan di dekat Fairbanks, Alaska, selama musim semi tahun 2017 untuk percobaan dekomposisi. Kami mengumpulkan serasah dari lokasi ini karena tersedia dalam jumlah besar dan kondisi jalan yang buruk menghalangi akses ke lokasi penelitian pada musim semi. Spesies semak yang ada di lokasi penelitian tidak terwakili di area Fairbanks; namun, serasah dari spesies berikut dikumpulkan sebagai gantinya untuk mewakili tiga genus target: Alnus rubra, Betula glandulosa, Betula occidentalis, Salix lasiandra, Salix bebbiana dan Salix alaxensis . Spesies ini memiliki fraksi kimia C dan N yang mirip dengan spesies target, yang kami konfirmasikan dengan membandingkan nilai kimia serasah awal untuk bahan yang dikumpulkan (Tabel S2 ) dengan nilai yang dipublikasikan untuk spesies target (Tabel S4 ).
Serasah daun yang sudah tua dikumpulkan langsung dari tanaman dan ditambah dengan daun yang sudah tua dari lapisan serasah paling atas (kedalaman 0–1 cm) jika diperlukan bahan tambahan. Bahan akar semak dikumpulkan dengan memotong cabang-cabang akar utama yang terhubung ke tanaman dari genus target, dan bahan akar Eriophorum dikumpulkan langsung di bawah rumpun. Bahan akar yang sebelumnya mati dibuang. Di laboratorium, akar penyerap (urutan pertama–ketiga) dibedah dengan hati-hati dari cabang-cabang akar semak untuk digunakan dalam kantong serasah. Untuk akar alang-alang yang tidak memiliki arsitektur percabangan, semua segmen akar individu dianggap sebagai akar penyerap. Baik bahan daun maupun akar dikeringkan dalam oven pada suhu 55°C selama 24 jam dan kemudian dikirim ke University of Illinois Urbana-Champaign. Semua bahan akar atau daun yang diambil sampelnya dari genus yang sama dicampur dengan baik untuk memaksimalkan jumlah serasah dan untuk memastikan bahwa kantong serasah menerima sampel serasah yang homogen. Subsampel serasah dari setiap genus digiling dan dianalisis menggunakan Costech Elemental Analyser untuk menentukan nilai C dan N awal. Lihat Lampiran S1 untuk informasi tambahan tentang prosedur pemanenan serasah.
Kami menyiapkan kantong serasah daun dan akar yang terpisah (jaring poliester berlapis vinil berdiameter 0,5 mm yang dijahit ke dalam kantong berukuran 10 × 10 cm) dengan bahan yang dikeringkan dalam oven dari satu genus. Kantong daun diisi dengan 1,0 g bahan daun yang sudah tua, dan kantong akar diisi dengan 0,5 g bahan akar penyerap yang telah dipotong. Meskipun kantong serasah diketahui dapat membatasi dekomposisi tergantung pada ukuran mata jaring (Lecerf, 2017 ), penggunaan mata jaring berukuran seragam di seluruh percobaan memungkinkan kami untuk mengukur perbedaan yang disebabkan oleh variabel yang diinginkan dalam desain.
Kantong serasah disebarkan pada akhir Juni dan awal Juli 2017. Untuk mengendalikan kualitas serasah sambil menguji efek iklim makro, iklim mikro, dan jenis vegetasi plot, kami melakukan eksperimen substrat umum, menempatkan satu kantong daun dan akar alang-alang di setiap plot alang-alang dan semak (340 kantong). Untuk menguji efek perubahan kualitas serasah yang disebabkan oleh semak, kami melakukan eksperimen kedua di mana kami memvariasikan genus sumber serasah (selanjutnya disebut eksperimen genus serasah), menempatkan dua kantong berisi serasah daun dari genus semak target di setiap plot semak dari genus yang sama (270 kantong). Sebagian kecil plot semak (67 dari 135) juga menerima satu kantong serasah akar dari genus semak target karena bahan akar terbatas (67 kantong). Secara total, 677 kantong serasah disebarkan di lima lokasi penelitian untuk mengevaluasi faktor-faktor yang mengatur dekomposisi serasah. Kantong daun ditempatkan secara horizontal pada permukaan substrat dan kantong akar secara vertikal pada kedalaman 0–10 cm.
Pada bulan Juli 2018, setelah 390 hari inkubasi, 657 kantong serasah diambil dan dikembalikan ke Illinois untuk dianalisis; 20 kantong yang awalnya digunakan tidak dapat dipindahkan di lapangan. Serasah dibersihkan dari bahan asing, dikeringkan dalam oven pada suhu 55°C selama 48 jam, dan kemudian ditimbang untuk menentukan kehilangan massa. Subsampel serasah kering dan terurai dari setiap kantong serasah digiling, dan persentase C dan N dianalisis dengan Costech Elemental Analyzer. Nilai-nilai ini dapat melebihi 100% karena imobilisasi N atau fiksasi N yang terjadi di substrat serasah. Semua data yang digunakan dalam penelitian ini tersedia untuk umum (Fraterrigo et al., 2025 ).
2.3 Analisis Statistik
Analisis statistik dilakukan dalam R v. 4.1.2 (Tim Inti R, 2021 ). Untuk menilai perbedaan dalam karakteristik kimia dari serasah daun dan akar awal, kami menyesuaikan model linier untuk %C dan %N menggunakan fungsi lm () dalam paket stats dan melakukan analisis post-hoc dengan penyesuaian Tukey dalam paket emmeans (Lenth, 2022 ). Untuk mengevaluasi perbedaan dalam kondisi iklim mikro antara empat jenis vegetasi dominan, kami menyesuaikan model efek campuran linier untuk suhu dan kelembapan tanah berdasarkan lokasi menggunakan fungsi lmer () dari paket lme4 (Bates et al., 2015 ), yang menentukan jenis vegetasi plot sebagai efek tetap dan klaster sebagai efek acak. Kami melakukan analisis post-hoc seperti yang dijelaskan sebelumnya, menggunakan perkiraan Satterthwaite untuk derajat kebebasan efektif.
Bahasa Indonesia: Untuk mengukur kontrol pada dekomposisi serasah daun dan akar serta fluks nutrisi, kami menggunakan pendekatan teori informasi, dengan mencocokkan model efek campuran linier hierarkis kandidat menggunakan fungsi lmer () dan kemudian melakukan pemilihan model dan perata-rataan model (Burnham & Anderson, 2004 ). Kami menjelaskan pendekatan kami secara singkat di bawah ini dan memberikan deskripsi terperinci dalam Lampiran S2 . Untuk percobaan substrat umum, model kandidat termasuk sebagai efek tetap: variabel iklim mikro tanah tunggal, jenis vegetasi plot, variabel iklim makro tunggal atau kombinasi interaksi aditif dan perkalian. Semua model termasuk efek acak dari klaster yang bersarang di dalam lokasi. Untuk percobaan genus serasah, kami memodifikasi set model kandidat dengan mengganti efek tetap jenis vegetasi plot dengan genus sumber serasah karena setiap genus serasah terurai di bawah genus semak yang sama. Semua efek tetap dan acak lainnya dipertahankan untuk model akar. Untuk model daun, kami mempertahankan efek tetap tetapi mengubah efek acak ke ID plot yang bersarang di dalam klaster yang bersarang di dalam lokasi karena dua kantong daun ditempatkan di setiap plot semak. Untuk mengevaluasi dukungan untuk model kandidat, kami menghitung AICc menggunakan paket AICcmodavg di R (Mazerolle, 2020 ) diikuti dengan perata-rataan model untuk mengukur ukuran efek rata-rata, SE, interval kepercayaan 95%, dan nilai kepentingan relatif untuk setiap variabel di seluruh model yang didukung (yaitu model dengan nilai AICc dalam dua nilai AICc terendah [ΔAICc < 2,0]).
Untuk menguji perbedaan antara dekomposisi serasah Eriophorum dan semak belukar sambil mengendalikan perbedaan iklim dan lingkungan dekomposisi, kami menyesuaikan model efek campuran linier seperti yang dijelaskan di atas dan melakukan ANOVA dengan perkiraan Satterthwaite untuk derajat kebebasan efektif menggunakan paket lmerTest di R (Kuznetsova et al., 2017 ). Kami menentukan jenis vegetasi, genus serasah dan interaksinya sebagai efek tetap, dan mengelompokkan yang bersarang di lokasi sebagai efek acak.
3 HASIL
3.1 Karakteristik sampah dan iklim mikro
Karakteristik kimia dari serasah daun dan akar awal berbeda di antara keempat genus. Sebelum dekomposisi, kami mengamati rasio C:N tertinggi untuk serasah daun Eriophorum , diikuti oleh Betula, Salix dan Alnus , masing-masing ( F = 607,2, p < 0,001; Tabel S2 ). Kami mengamati pola yang sama pada akar, dengan rasio C:N akar Eriophorum kira-kira dua kali lebih besar daripada akar semak (Tabel S2 ). Pola ini terutama didorong oleh perbedaan N, karena kandungan C relatif stabil antar genus.
Pola iklim mikro tanah bulan Juli bervariasi di antara jenis vegetasi dan lokasi. Suhu tanah umumnya paling tinggi di plot Eriophorum dan sebanding di antara plot semak (Tabel S3 ). Perbedaan ini paling menonjol di lokasi selatan. Untuk kelembaban tanah, plot Betula cenderung lebih kering (kandungan air volumetrik 16,4%), sedangkan kelembaban tanah serupa di antara plot Eriophorum , Alnus dan Salix , berkisar antara 19% hingga 20% (Tabel S3 ). Perbedaan kelembaban tanah umumnya lebih besar antara plot Betula dan Salix di lokasi utara dibandingkan dengan lokasi selatan (Tabel S3 ).
3.2 Percobaan substrat umum: Dekomposisi serasah alang-alang di petak semak dan alang-alang
Dekomposisi serasah daun Eriophorum bervariasi dengan iklim makro dan iklim mikro tanah (Gambar 2a ; Tabel S5 ), dengan suhu udara musim panas lebih dari dua kali lebih penting daripada suhu tanah untuk mengatur kehilangan massa daun (Tabel S6 ). Setelah 1 tahun inkubasi, kehilangan massa serasah daun Eriophorum adalah 54,6% lebih tinggi di lokasi selatan dengan suhu udara musim panas rata-rata yang lebih tinggi daripada di lokasi utara dengan suhu musim panas yang lebih rendah. Kehilangan massa berhubungan negatif dengan suhu tanah di seluruh lokasi, tetapi interval kepercayaan 95% untuk parameter ini melewati nol (Tabel S6 ). Jenis vegetasi plot tidak mempengaruhi dekomposisi serasah daun, dengan kehilangan massa rata-rata 25,8% ± 1,9% (SE) di plot teki dan 25,9% ± 1,5% di plot semak di semua lokasi (Gambar S2 ).
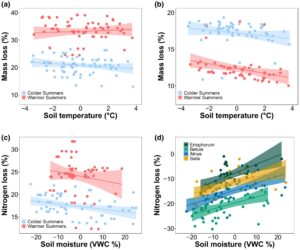
Kehilangan nitrogen dari serasah daun Eriophorum juga bervariasi menurut iklim makro dan iklim mikro, dengan kehilangan N dari daun meningkat menurut suhu udara musim panas rata-rata dan curah hujan musim panas rata-rata tetapi menurun menurut kelembaban tanah (Gambar 2c ; Tabel 1 ). Namun, hanya suhu udara musim panas rata-rata yang memiliki interval kepercayaan 95% yang tidak melewati nol (Tabel S6 ). Tidak ada pengaruh jenis vegetasi plot pada kehilangan N dari serasah daun (Gambar S2 ).
| Marga | Serasah daun | Serasah akar | ||
|---|---|---|---|---|
| Kehilangan massa | Kehilangan N | Kehilangan massa | Kehilangan N | |
| Erioforum (Erioforum spp.) | 25,6 ± 0,8 | 17,8 ± 1,5 | 15,12 ± 0,6 | -14,01 ± 2,6 |
| Betula | 21,1 ± 0,6 | -3,5 ± 1,4 | 24,11 ± 1,6 | 7,53 ± 2,7 |
| Alnus | 22,2 ± 0,7 | 11,0 ± 1,0 | 24,40 ± 2,1 | 7,80 ± 4,3 |
| Salix | 28,9 ± 0,9 | 17,7 ± 1,4 | 20,84 ± 2,1 | 22,39 ± 2,4 |
Catatan : Nilai yang dicetak tebal menunjukkan perbedaan signifikan antara Eriophorum dan serasah semak untuk respon tertentu ( p < 0,05).
Sementara makroiklim dan mikroiklim tanah memengaruhi kehilangan massa serasah akar Eriophorum , beberapa efeknya berlawanan dengan serasah daun (Gambar 2b ; Tabel 1 ). Secara khusus, kehilangan massa serasah akar menurun seiring suhu udara musim panas (Tabel S6 ). Kehilangan massa juga menurun seiring suhu tanah dan meningkat seiring kelembapan tanah, tetapi interval kepercayaan 95% untuk kedua parameter ini melewati nol (Tabel S6 ). Kami tidak menemukan bukti bahwa jenis vegetasi plot memengaruhi kehilangan massa serasah akar, yang rata-rata 14,1% ± 1,2% di plot alang-alang dan 15,1% ± 1,0% di plot semak (Gambar S2 ).
Kehilangan N serasah akar adalah satu-satunya respon yang bervariasi dengan tipe vegetasi plot (Gambar 2d ; Tabel S5 ). Dibandingkan dengan plot teki, kehilangan N secara substansial lebih rendah di plot Betula (Tabel S6 ). Plot Alnus dan Salix juga menunjukkan kehilangan N serasah akar yang lebih rendah, dan parameter-parameter ini memiliki kepentingan relatif yang tinggi; namun, interval kepercayaan 95% untuk parameter-parameter ini melewati nol (Tabel S6 ). Khususnya, kelembaban tanah dan suhu musim panas rata-rata juga memiliki kepentingan relatif yang tinggi, dengan kehilangan N akar meningkat dengan kelembaban tanah dan suhu udara musim panas. Namun, interval kepercayaan 95% untuk parameter-parameter ini melewati nol. Meskipun curah hujan musim panas rata-rata berhubungan positif dengan kehilangan N, kepentingan relatifnya rendah dan interval kepercayaan 95% melewati nol (Tabel S6 ).
3.3 Percobaan genus serasah: Dekomposisi serasah semak di bawah semak
Genus serasah semak sangat memengaruhi hilangnya massa serasah daun (Gambar 3a ; Tabel S5 ). Rata-rata, serasah daun Salix kehilangan massa 34,7% dan 28,1% lebih banyak daripada Betula dan Alnus , masing-masing, sementara hilangnya massa serasah daun Betula dan Alnus serupa (Tabel S7 ). Hilangnya massa menurun seiring dengan kelembaban tanah, tetapi ada interaksi positif antara kelembaban tanah dan genus serasah, sehingga Salix dan Alnus kehilangan lebih banyak massa seiring dengan peningkatan kelembaban tanah. Namun, interval kepercayaan untuk interaksi ini mengandung nol (Tabel S7 ).
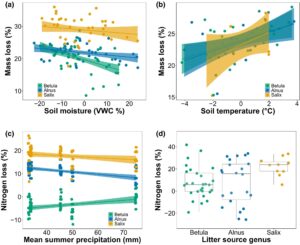
Genus serasah semak juga sangat memengaruhi hilangnya N serasah daun (Gambar 3c ; Tabel S5 dan S7 ). Dibandingkan dengan Betula , serasah Alnus dan Salix melepaskan lebih banyak N, dengan Salix melepaskan 60,9% lebih banyak N daripada Alnus secara rata-rata. Sebaliknya, serasah daun Betula melumpuhkan N, dengan peningkatan N sebesar 3,5%. Namun, terdapat interaksi positif antara genus serasah dan curah hujan musim panas rata-rata, sehingga hilangnya N dari serasah daun Alnus dan Salix menurun seiring dengan curah hujan, sementara hilangnya N dari serasah Betula meningkat seiring dengan curah hujan (Gambar 3c ; Tabel S7 ). Selain itu, terdapat interaksi negatif antara suhu udara musim panas dan kelembapan tanah, sehingga hilangnya N serasah daun menurun seiring dengan kelembapan tanah ketika suhu udara musim panas meningkat (Gambar S3 ; Tabel S7 ).
Tidak seperti serasah daun, kehilangan massa serasah akar semak terutama bervariasi dengan iklim makro dan iklim mikro dan tidak berbeda dengan genus serasah (Gambar 3b ; Tabel S5 ). Kehilangan massa serasah akar semak meningkat dengan suhu tanah, sedangkan itu menurun dengan suhu udara musim panas dan curah hujan musim panas (Tabel S7 ). Selain itu, ada interaksi positif antara suhu musim panas rata-rata dan suhu tanah, sehingga kehilangan massa meningkat lebih tajam dengan suhu tanah ketika suhu udara musim panas meningkat (Gambar S4 ). Namun, interval kepercayaan untuk semua parameter dalam model rata-rata melewati nol (Tabel S7 ).
Kehilangan N akar diatur secara eksklusif oleh genus serasah (Gambar 3d ; Tabel S5 ). Salix kehilangan lebih banyak N daripada Betula (198,7% lebih tinggi), sementara kehilangan N Alnus dan Betula sebanding (Tabel S7 ). Tidak ada pengaruh iklim makro atau iklim mikro tanah terhadap kehilangan N serasah akar.
3.4 Dekomposisi serasah berdasarkan gugus fungsi
Bahasa Indonesia: Ketika mengendalikan lokasi dan lingkungan dekomposisi, kami menemukan perbedaan yang jelas dalam dekomposisi Eriophorum dan serasah semak, tetapi polanya berlawanan untuk serasah daun dan akar (Tabel 1 ). Serasah daun Betula kehilangan 18% lebih sedikit massa dan 120% lebih sedikit N daripada serasah daun Eriophorum , sedangkan serasah akar Betula kehilangan 60% lebih banyak massa dan 154% lebih banyak N daripada serasah akar Eriophorum (semua p < 0,001). Kami menemukan perbedaan serupa antara Alnus dan Eriophorum untuk kehilangan massa daun ( p < 0,05), kehilangan massa akar ( p < 0,001) dan kehilangan N akar ( p < 0,001). Demikian pula, akar Salix kehilangan lebih banyak massa (38%) dan N (260%) daripada akar Eriophorum ( p < 0,05). Tidak seperti genus semak lainnya, bagaimanapun, kehilangan massa serasah daun Salix secara signifikan lebih besar daripada Eriophorum ( p < 0,01).
4 DISKUSI
Sementara hasil kami mengonfirmasi bahwa iklim makro adalah kontrol penting pada dekomposisi, mereka menunjukkan bahwa faktor lokal yang terkait dengan perluasan semak dapat mengatur dekomposisi serasah dengan kuat pada skala regional. Ketika mengendalikan kualitas serasah menggunakan serasah teki sebagai substrat umum, kami menemukan bahwa iklim makro adalah kontrol dominan pada hilangnya massa daun dan akar dan hilangnya N daun, meskipun itu memiliki efek yang berlawanan pada hilangnya massa serasah daun dan akar (Tabel S6 ). Namun, ketika kualitas serasah dibiarkan bervariasi, kami menemukan bahwa genus serasah umumnya memiliki efek terkuat pada dekomposisi, dengan iklim makro dan iklim mikro memodulasi efek ini (Tabel S7 ). Rata-rata, serasah akar semak terurai lebih cepat daripada serasah akar teki dan dalam koordinasi dengan kualitas serasah, sedangkan dekomposisi serasah daun semak lebih bervariasi. Hasil ini menggarisbawahi bahwa pemanasan iklim dan perluasan semak, sebagian besar melalui efeknya pada kualitas serasah, akan mengubah pola regional dekomposisi serasah di ekosistem tundra Arktik. Meskipun dekomposisi serasah semak dikoordinasikan dengan kualitas serasah sampai batas tertentu, kombinasi faktor pengatur yang berbeda cenderung melemahkan koordinasi antara dekomposisi daun dan akar pada skala spasial yang luas.
4.1 Faktor-faktor yang menentukan variasi dekomposisi serasah Eriophorum
Sesuai dengan model hierarkis tradisional dekomposisi (Swift et al., 1979 ), penelitian sebelumnya menunjukkan bahwa iklim mengendalikan dekomposisi di Arktik (Breeuwer et al., 2008 ; Xue et al., 2016 ). Hasil kami mendukung temuan ini, karena variabel makroiklim yang kami selidiki menghasilkan ukuran efek terbesar untuk kehilangan massa daun dan kehilangan N, dan kehilangan massa akar, ketika kami mengendalikan kualitas serasah dengan menggunakan substrat umum yang bersumber dari lokasi di luar lokasi (Tabel S6 ). Hubungan positif yang kami amati antara suhu udara musim panas rata-rata dan kehilangan massa daun dan kehilangan N konsisten dengan penelitian lain di mana laju dekomposisi dan mineralisasi N meningkat dengan pemanasan (Aerts, 2006 ; Hobbie, 1996 ; Schaeffer et al., 2013 ). Temuan ini menyoroti potensi pemanasan iklim untuk mempercepat siklus C dan N yang terkait dengan dekomposisi serasah daun di Arktik.
Menariknya, arah efek ini berlawanan untuk kehilangan massa akar. Kami menemukan bahwa serasah akar Eriophorum menurun dengan suhu udara musim panas rata-rata (Gambar 2 , Tabel S6 ). Pola ini mungkin disebabkan oleh kovariasi antara suhu dan pola biogeografis dalam kelimpahan relatif bakteri versus jamur di tanah, serta perbedaan dalam kesesuaian substrat untuk bakteri versus jamur. Secara khusus, penelitian menunjukkan bahwa bakteri awalnya mendominasi dekomposisi serasah labil berkualitas tinggi dan jamur awalnya mendominasi dekomposisi serasah bandel berkualitas rendah (Paul & Clark, 1996 ). Sebuah studi global tentang dekomposisi daun dan akar selanjutnya menemukan bahwa biomassa jamur berkorelasi negatif dengan dekomposisi serasah daun (Guo et al., 2021 ). Pada saat yang sama, penelitian telah mengungkapkan tren lintang di mana dominasi jamur meningkat relatif terhadap bakteri di daerah dingin dan lintang tinggi dengan stok karbon lebih tinggi (Yu et al., 2022 ), yang menunjukkan bahwa jamur mungkin dominan atas bakteri di lokasi kita yang lebih dingin. Oleh karena itu, biomassa bakteri yang lebih tinggi di lokasi yang lebih hangat dapat mendukung dekomposisi serasah daun yang lebih cepat, sementara biomassa jamur yang lebih tinggi di lingkungan yang lebih dingin dapat mendukung dekomposisi serasah akar yang lebih cepat, seperti yang kami temukan di sini. Penelitian tambahan diperlukan untuk menentukan bagaimana pola biogeografis yang mapan dalam komunitas mikroba tanah memengaruhi pola dekomposisi serasah daun dan akar di tundra Arktik. Penelitian mendatang juga harus mengevaluasi apakah temuan ini kuat terhadap serasah yang bersumber dari spesies dan lokasi lain, mengingat bahwa temuan kami adalah untuk substrat yang dikumpulkan dari lokasi selain tempat mereka diinkubasi, dan pengurai mikroba dapat beradaptasi dengan serasah lokal (yaitu keuntungan dari kandang sendiri; Ayres et al., 2009 ).
Di hutan beriklim sedang, efek komunitas dekomposer pada dekomposisi dapat lebih besar daripada efek iklim makro (Bradford et al., 2017 ). Namun, sementara perluasan semak peluruh dikaitkan dengan perubahan komposisi komunitas mikroba (Clemmensen et al., 2021 ; Wallenstein et al., 2007 ), kami tidak mendeteksi efek jenis vegetasi plot pada hilangnya massa serasah daun atau akar, atau hilangnya N serasah daun. Peneliti lain sebelumnya menemukan bukti efek vegetasi pada hilangnya massa serasah daun yang lebih besar daripada efek iklim dari simulasi pemanasan satu derajat (Ward et al., 2015 ). Sebaliknya, penelitian kami mencakup gradien suhu tahunan iklim makro yang lebih besar sebesar 3,5°C, yang dapat menjelaskan perbedaan dalam hasil kami. Meskipun demikian, kami mengamati efek jenis vegetasi plot pada hilangnya N serasah akar, dengan hilangnya N menurun paling banyak di bawah Betula , diikuti oleh Alnus dan Salix . Dalam studi pendamping di lokasi yang sama, kami menemukan bahwa kolonisasi ektomikoriza pada akar paling tinggi terjadi pada Betula , diikuti oleh semak Alnus dan Salix (Chen et al., 2020 ). Hal ini menimbulkan kemungkinan bahwa jamur ektomikoriza menghambat hilangnya N serasah akar dengan menekan aktivitas jamur saprofit yang hidup bebas (yaitu ‘efek Gadgil’; Gadgil & Gadgil, 1971 ). Namun, penelitian terbaru menunjukkan bahwa efek ini bersifat sementara, hanya terjadi selama tahap awal dekomposisi (<2 tahun), dan jamur ektomikoriza malah merangsang dekomposisi serasah seiring berjalannya waktu (Jonsson et al., 2025 ). Oleh karena itu, studi yang lebih lama mungkin diperlukan untuk memisahkan berbagai efek dan interaksi yang disebabkan oleh perluasan semak.
4.2 Variabilitas dekomposisi serasah semak berdaun gugur
Kualitas serasah semakin dihargai sebagai pengatur utama dekomposisi serasah (Cornwell et al., 2008 ; Makkonen et al., 2012 ; Zhang et al., 2008 ). Hasil percobaan genus serasah kami mendukung hal ini dan menunjukkan bahwa perbedaan kualitas serasah karena perluasan semak akan menjadi pendorong utama variasi pola dekomposisi serasah di seluruh Arktik (Tabel S7 ). Banyak penelitian sebelumnya telah menyelidiki Betula nana tetapi belum membandingkan hasilnya dengan genera lain dalam kelompok fungsional semak peluruh daun. Kami mengukur laju dekomposisi relatif serasah yang dikumpulkan dari lokasi yang sama di Fairbanks, Alaska, dari spesies dalam genus yang sama dengan spesies semak peluruh daun yang meluas di Alaska utara. Sementara semua genera semak peluruh daun yang kami periksa meluas dalam jangkauannya, kami menemukan bahwa laju dekomposisi semak peluruh daun Arktik tidak dapat digeneralisasi ke tingkat kelompok fungsional. Serasah daun Salix kehilangan massa secara signifikan lebih banyak dibandingkan dengan Betula dan Alnus , sementara kami tidak menemukan perbedaan antara kehilangan massa serasah daun Betula dan Alnus . Kami berhipotesis bahwa hilangnya massa serasah Salix yang lebih cepat disebabkan oleh perbedaan kimia pada daun Salix yang membuatnya lebih labil. Misalnya, Palo ( 1984 ) menemukan spesies Salix memiliki lebih sedikit polifenol, yang merupakan senyawa yang relatif membandel, dibandingkan dengan spesies Betula . Meskipun serasah daun Salix juga memiliki rasio C:N yang relatif rendah, rasio tersebut lebih tinggi daripada Alnus (Tabel S2 ). Kami sebelumnya menemukan bahwa akar Salix memiliki tingkat kolonisasi ektomikoriza yang lebih rendah (Chen et al., 2020 ) dan nilai δ 15 N yang menunjukkan bahwa Salix lebih bergantung pada N anorganik dibandingkan dengan Betula dan Alnus (Fraterrigo et al., 2024 ). Lebih jauh lagi, spesies Salix sering kali merupakan tanaman mikoriza ganda yang berasosiasi dengan jamur mikoriza ektomikoriza dan arbuskular, yang memungkinkan mereka memperoleh N anorganik lebih cepat (Teste et al., 2020 ). Dengan demikian, serasah Salix dapat terurai lebih cepat dan melepaskan lebih banyak N anorganik untuk mendukung kebutuhan nutrisinya sendiri, sehingga terlibat dalam umpan balik serasah semak yang positif (DeMarco et al., 2014 ) sejalan dengan strategi penawaran dan permintaan nutrisi yang terkoordinasi (Guo et al., 2021 ).
Kami juga menemukan bahwa kelembapan tanah memiliki efek negatif yang kuat pada dekomposisi serasah daun semak (Tabel S7 ). Penelitian lain telah menemukan bahwa kelembapan tanah yang lebih tinggi dapat meningkatkan pertumbuhan semak Arktik (Ackerman et al., 2017 ; Myers-Smith et al., 2015 ) dan kolonisasi akar ektomikoriza (Fraterrigo et al., 2024 ) di bawah suhu yang menghangat. Pertumbuhan semak yang lebih cepat dan kelimpahan ektomikoriza yang lebih tinggi di lokasi mikro yang lebih basah berpotensi memperlambat dekomposisi dengan mengurangi jumlah N yang tersedia untuk saprotrof (Clemmensen et al., 2006 ). Mengingat pemahaman yang terbatas tentang bagaimana jamur mikoriza mempengaruhi pembusukan serasah di tundra Arktik, penelitian lebih lanjut diperlukan untuk mengevaluasi hipotesis ini. Selain itu, karena dekomposisi serasah daun lebih sensitif terhadap kelembapan tanah daripada dekomposisi serasah akar, peran kelembapan tanah dalam melemahkan koordinasi antara dekomposisi serasah daun dan akar pada skala spasial yang luas harus diselidiki.
Kami menduga bahwa semua serasah daun akan kehilangan N selama 1 tahun dan bahwa divergensi akan terkait dengan peningkatan kadar N tanah di sekitar Alnus karena asosiasinya dengan Frankia yang memfiksasi N. Namun, kami menemukan bahwa Alnus dan Salix kehilangan jumlah N yang sebanding pada serasah daunnya, sementara N daun Betula meningkat (Tabel S7 ). Perbedaan yang diamati dalam kehilangan N antara serasah semak mungkin sebagian dijelaskan oleh kandungan lignin pada serasah. Kandungan lignin dan lignin:N dilaporkan berkorelasi terbalik dengan kehilangan massa serasah (Aerts, 1997 ), dan sementara kami tidak mengukur kandungan lignin secara langsung, peneliti lain telah melaporkan variasi intraspesifik dan musiman di antara kandungan lignin daun di B. nana dan S. pulchra (Chapin et al., 1986 ). Lebih jauh, yang lain telah menemukan bahwa serasah Betula memiliki rasio lignin yang relatif tinggi (DeMarco et al., 2014 ; McLaren et al., 2017 ). Dengan demikian, N dalam serasah daun Betula mungkin lebih terlindungi oleh lignin, sehingga mengakibatkan imobilisasi N, sedangkan N dalam genera lain yang kami selidiki lebih rentan terhadap pelindian dan akses mikroba.
Secara keseluruhan, model dekomposisi akar kami tidak menggambarkan variabilitas dalam data sebaik model dekomposisi daun kami. Nilai R2 marginal untuk model akar yang didukung atas secara konsisten lebih rendah daripada model daun (Tabel S5 ). Sesuai dengan hipotesis kedua kami, satu penjelasannya adalah bahwa akar membusuk lebih lambat dan dengan demikian durasi studi yang lebih lama diperlukan untuk mengevaluasi faktor regulasi. Sebuah meta-analisis global menemukan bahwa serasah akar halus membusuk hampir dua kali lebih lambat dibandingkan dengan jaringan daun (Guo et al., 2021 ) . Hasil kami juga konsisten dengan penelitian di Arktik yang menemukan lebih sedikit variasi dalam dekomposisi antara berbagai jenis serasah akar dibandingkan dengan daun (McLaren et al., 2017 ). Yang lain telah menyarankan bahwa perbedaan antara tingkat dekomposisi di antara serasah daun dan akar terkait dengan disjungsi antara sifat daun dan akar tanaman. Misalnya, sifat-sifat yang memprediksi dekomposisi akar (misalnya diameter akar dan kandungan hemiselulosa) tidak selalu berkorelasi dengan sifat-sifat yang memprediksi dekomposisi serasah daun (misalnya lignin awal, konsentrasi Ca dan lignin:N) (Sun et al., 2018 ). Oleh karena itu, mengukur sifat-sifat di bawah tanah selain sifat-sifat di atas tanah mungkin penting untuk memprediksi pola dekomposisi kompleks dalam sistem Arktik secara akurat.
5 KESIMPULAN
Hasil kami menunjukkan bahwa kontrol lokal dapat memengaruhi pola dekomposisi regional di Kutub Utara. Kami menunjukkan bahwa perluasan semak peluruh daun dapat mengubah dekomposisi serasah secara signifikan dengan memperkenalkan serasah baru. Perbedaan intergenerik dalam kualitas serasah ini dapat berinteraksi dengan kontrol penting lainnya pada dekomposisi, seperti iklim makro dan mikro. Yang terpenting, hasil kami menunjukkan bahwa perluasan semak yang berkelanjutan akan mengurangi kumpulan karbon dan nitrogen serasah akar di ekosistem tundra. Namun, dekomposisi serasah daun dan akar menunjukkan kontrol yang berbeda, dengan efek iklim makro dan mikro yang bergeser dalam arah dan besarnya antara setiap jenis serasah. Oleh karena itu, memperhitungkan variasi iklim mikro dan kualitas serasah antara semak dalam kelompok fungsional semak peluruh daun mungkin penting untuk meningkatkan prediksi dinamika C dan N, termasuk siklus nutrisi, penyimpanan karbon, dan respirasi tanah, di Kutub Utara yang berubah.


