
ABSTRAK
Studi ini meneliti pola asosiasi, pengelompokan sosial, dan stabilitas temporal lumba-lumba hidung botol Indo-Pasifik di lepas North West Cape (NWC), Australia Barat. Selama 6 tahun, survei berbasis perahu mengumpulkan data identifikasi foto, yang memungkinkan pengambilan sampel berulang, konstruksi riwayat penampakan, dan identifikasi asosiasi. Di antara 344 lumba-lumba dewasa yang diidentifikasi, 116 terlihat setidaknya lima kali dan digunakan dalam analisis asosiasi. Korelasi yang kuat ( r = 0,86 ± 0,01 SE) antara indeks asosiasi sebenarnya dan yang diestimasikan, bersama dengan koefisien variasi yang tinggi ( S = 1,92 ± 0,9 SE), menunjukkan masyarakat yang sangat terdiferensiasi dengan hubungan yang beragam di antara individu. Indeks afiliasi umum, metrik jaringan sosial, dan tingkat asosiasi yang tertinggal mengungkapkan asosiasi yang sebagian besar longgar dan jangka pendek dalam jaringan sosial yang terhubung dengan baik tetapi tidak berkelompok. Pola asosiasi yang cair ini kemungkinan terkait dengan sifat populasi ini yang terbuka dan sementara, tempat individu masuk dan keluar dari area studi. Pergerakan ini memberikan peluang bagi interaksi sosial di antara individu yang berbeda, dan anggota kelompok yang berbeda dapat terpecah dalam kurun waktu yang berbeda-beda. Temuan dari penelitian ini memberikan dasar yang kuat untuk penelitian selanjutnya yang mengamati struktur sosiogenetik populasi ini dan faktor lingkungan dan sosial yang mendorong sistem sosial mereka yang cair.
1 Pendahuluan
Pola asosiasi di antara hewan-hewan individu yang hidup dalam kelompok bervariasi di seluruh ruang dan waktu dan dianggap dipengaruhi oleh berbagai faktor ekologi, fenotipik, dan perilaku (Croft et al. 2005 ). Variasi spasiotemporal dalam pola asosiasi tersebut telah menyebabkan munculnya berbagai macam struktur sosial hewan (Whitehead 1997 ). Beberapa masyarakat hewan terdiri dari komunitas-komunitas berbeda yang dicirikan oleh asosiasi yang stabil dan jangka panjang, sementara yang lain dicirikan oleh asosiasi yang longgar dan cair serta tingkat dinamika fisi-fusi yang tinggi (Couzin 2006 ). Dalam masyarakat yang dicirikan oleh tingkat dinamika fisi-fusi yang tinggi, individu-individu membentuk kelompok yang sering berubah dalam ukuran dan komposisi, dengan para anggotanya berasosiasi, disosiasi, dan kemudian berasosiasi kembali secara sering (Aureli et al. 2008 ; Conradt dan Roper 2005 ; Couzin 2006 ). Dinamika fisi–fusi ini mencirikan struktur sosial dari berbagai jenis ikan (Croft et al. 2005 ), reptil (Strickland et al. 2014 ), dan spesies mamalia (Archie et al. 2008 ; Ilany et al. 2015 ; Kerth et al. 2006 ), termasuk beberapa delphinida (Connor et al. 2000 ; Kiszka et al. 2015 ).
Delphinid hidup dalam kelompok dengan berbagai macam dinamika fisi–fusi. Ini termasuk spesies dengan ikatan stabil jangka panjang, seperti kelompok matrilineal paus pembunuh ( Orcinus orca ) (Baird dan Whitehead 2000 ), ikatan cair kelompok besar lumba-lumba Hector ( Cephalorhynchus hectori ) (Constantine 2019 ), dan asosiasi temporal yang bervariasi dari lumba-lumba hidung botol ( Tursiops spp.) (Connor et al. 2000 ). Pola asosiasi di antara masing-masing delphinid dipengaruhi oleh berbagai faktor, termasuk ketersediaan makanan, risiko predasi, tempat tinggal, demografi, riwayat hidup, dan hubungan khusus jenis kelamin yang terkait dengan persaingan intraseksual dan konflik interseksual (Connor et al. 2000 ; Gowans et al. 2007 ; Möller 2012 ; Parra et al. 2006 ). Interaksi individu dalam taksa yang hidup berkelompok ini tidak hanya memengaruhi struktur sosial mereka, tetapi juga memengaruhi proses populasi utama, termasuk kebugaran (Stanton dan Mann 2012 ), penularan penyakit (Szott et al. 2022 ; Weiss et al. 2021 ), dan struktur populasi genetik (Möller 2012 ). Dengan demikian, pengetahuan tentang pola asosiasi di antara individu dan struktur serta stabilitas temporal mereka merupakan elemen kunci untuk memahami populasi, ekologi, evolusi, dan konservasi mereka.
Lumba-lumba hidung botol Indo-Pasifik ( Tursiops aduncus ) terutama ditemukan di habitat pesisir dan muara yang dangkal dan terumbu karang dekat pantai di seluruh jangkauannya dari Australia hingga Asia Tenggara, Jepang, dan Afrika Selatan bagian timur (Braulik et al. 2019 ). Spesies ini diklasifikasikan sebagai Hampir Terancam oleh International Union for the Conservation of Nature (IUCN) karena distribusi pesisir dan intensitas ancaman di seluruh jangkauannya (Braulik et al. 2019 ). Penelitian telah mendokumentasikan variasi intraspesifik yang nyata dalam organisasi sosial dan dinamika fisi-fusi di berbagai populasi. Misalnya, asosiasi jangka panjang dan stabil telah diamati di beberapa komunitas (misalnya, Ansmann et al. 2012 ; Wiszniewski et al. 2009 ), sementara yang lain menunjukkan asosiasi yang lebih jangka pendek dan cair (misalnya, Connor et al. 1992 ). Penelitian dari Australia Barat, seperti di Shark Bay dan Bunbury, juga telah mengungkap struktur sosial yang kompleks dengan stratifikasi sosial yang jelas dan aliansi yang kompleks, yang menekankan kemampuan beradaptasi lumba-lumba hidung botol Indo-Pasifik terhadap kondisi lingkungan setempat dan pentingnya organisasi sosial dalam membentuk struktur komunitas dan dinamika ekosistem (Connor dan Krützen 2015 ; Connor et al. 2000 ; Randić et al. 2012 ; Smith 2012 ; Sprogis et al. 2016 ; Tsai dan Mann 2013 ).
Lumba-lumba hidung botol Indo-Pasifik dikenal memiliki nilai ekologi yang cukup besar bagi Taman Laut Ningaloo (NMP) di Australia Barat (CALM dan MPRA 2005 ). Penelitian terkini menunjukkan bahwa kawasan ini merupakan habitat penting bagi spesies ini, dengan individu penghuni dan bukan penghuni menggunakan perairan dekat pantai North West Cape (NWC) sepanjang tahun (Haughey et al. 2020 , 2021 ) dan total ukuran superpopulasi sekitar 370 (95% CI: 333–407) individu. Akan tetapi, sedikit yang diketahui tentang pola asosiasi dan struktur sosial lumba-lumba hidung botol di kawasan ini dan apakah mereka mengikuti pola yang sama dengan populasi lain.
Dalam penelitian ini, kami menggunakan riwayat penampakan lumba-lumba hidung botol dewasa yang diidentifikasi melalui foto di sekitar NWC untuk menyelidiki struktur sosial mereka dan menilai kekuatan asosiasi, pengelompokan sosial, dan variabilitas temporal dalam pola asosiasi. Mengingat temuan penelitian sebelumnya (Haughey et al. 2020 ), kami berhipotesis bahwa asosiasi di antara lumba-lumba hidung botol secara individu akan bersifat nonacak dengan ikatan yang longgar secara temporal mengingat sifat demografis populasi ini yang terbuka dan sementara.
2 Metode
2.1 Lokasi Penelitian
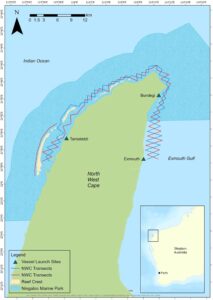
Daerah studi meliputi perairan pantai dangkal yang mengelilingi NWC, yang terletak di Australia Barat laut (Gambar 1 ). NMP meliputi sisi barat dan timur laut NWC, dengan batas tenggara di tengah-tengah antara tanjakan perahu Bundegi dan Exmouth. Daerah studi NWC seluas 130 km 2 dan membentang dari Teluk Exmouth di sisi timur, di sekitar ujung tanjung, hingga Teluk Mangrove di sisi barat. Sisi barat NWC menampilkan Ningaloo Reef, terumbu karang tepian terbesar di Australia, yang dipisahkan dari pantai oleh laguna berpasir sempit dan komunitas terumbu karang (Cassata dan Collins 2008 ). Daerah tersebut terpapar energi gelombang tinggi melintasi landas kontinen yang sempit (Cassata dan Collins 2008 ; Collins et al. 2003 ; CALM dan MPRA 2005 ). Di sisi timur NWC terdapat Teluk Exmouth, sebuah muara besar dengan kedalaman rata-rata 11,9 m (Brunskill et al. 2001 ). Teluk Exmouth merupakan tempat pembibitan penting bagi banyak spesies dan terdiri dari hutan bakau, substrat pasir dan lumpur, serta terumbu karang (Fitzpatrick et al. 2019 ).
2.2 Pengumpulan Data
Survei berbasis perahu untuk lumba-lumba hidung botol Indo-Pasifik dilakukan di seluruh wilayah studi dari bulan April hingga Oktober selama 6 tahun (2013, 2014, 2015, 2018, 2019, dan 2021). Survei tidak dilakukan dari bulan November hingga Maret karena kondisi angin yang tidak menguntungkan, risiko siklon tropis yang lebih tinggi, dan suhu ekstrem.
Survei berbasis perahu dan identifikasi foto mengikuti protokol yang dijelaskan oleh Haughey et al. ( 2020 ). Survei hanya dilakukan dalam kondisi penampakan yang baik (Beaufort Sea State
3 dan tidak ada hujan) dari kapal penelitian konsol tengah 5,8 m yang ditenagai oleh motor tempel 100 hp yang melaju dengan kecepatan 10–12 km/jam. Upaya pencarian terus menerus dari pukul 07.00 hingga 17.00, tergantung pada kondisi cuaca yang sesuai. Survei mengikuti tata letak garis transek sistematis, yang mencakup berbagai habitat dan area penggunaan manusia di dalam area studi. Tata letak transek terdiri dari dua garis “zigzag” yang berlawanan sepanjang 93 km yang mencakup 49 kaki transek dan garis tambahan sepanjang 13 km yang mencakup lima kaki transek (Gambar 1 ). Masing-masing kaki transek ini bervariasi dari 1 hingga 4 km panjangnya dan diperpanjang hingga 3,7 km dari pantai.
Selama survei, dua pengamat terlatih mencari lumba-lumba dengan mata telanjang dan teropong 7 × 50 dari 0° hingga 90° dari garis transek di sisi kiri dan kanan kapal. Pengamat ketiga (nakhoda) memindai 180° di depan kapal, dan perekam data khusus mencatat data upaya survei dan setiap perubahan kondisi laut dan cuaca. Kelompok lumba-lumba, yang mewakili unit pengambilan sampel dalam penelitian kami (Syme et al. 2022 ), secara operasional didefinisikan sebagai individu dalam kedekatan spasial yang dekat (yaitu, individu dalam jarak 100 m satu sama lain) dan terlibat dalam aktivitas perilaku yang serupa (Hunt et al. 2017 ). Bahasa Indonesia: Ketika sekelompok lumba-lumba terlihat, upaya survei dihentikan, dan perahu meninggalkan garis transek untuk perlahan-lahan (< 10 km/jam) mendekati lumba-lumba untuk mengumpulkan data berikut: lokasi GPS, spesies, ukuran kelompok, komposisi usia kelompok (yaitu, jumlah dewasa, remaja, dan anak lumba-lumba), perilaku kelompok yang dominan, dan kekompakan kelompok. Perilaku kelompok dan kekompakan kelompok didasarkan pada apa yang ditunjukkan oleh lebih dari 50% lumba-lumba dalam kelompok pada awal pengamatan (Mann dan Smuts 1999 ). Tiga kategori usia untuk komposisi kelompok dibedakan berdasarkan ukuran tubuh dan kriteria perilaku sebagai berikut: (1) dewasa, individu > 2 m panjang; (2) remaja, individu antara ½ dan ¾ panjang tubuh dewasa, biasanya berenang dalam hubungan dekat dengan orang dewasa, tetapi kadang-kadang berenang secara mandiri; (3) anak lumba-lumba, individu
½ panjang ikan dewasa, ditemukan dalam hubungan dekat dengan ikan dewasa, dan berenang secara teratur di samping atau sedikit di belakang ikan dewasa (Parra et al. 2006 ). Saat data direkam, gambar identifikasi foto diambil dari semua individu dalam kelompok. Gambar sirip punggung diambil menggunakan kamera refleks lensa tunggal digital dengan lensa zoom telefoto 80–400 mm. Setelah semua individu dalam kelompok difoto atau kelompok tersebut hilang, perahu kembali ke garis transek dan melanjutkan upaya survei. Penampakan di luar upaya juga direkam dan digunakan dalam analisis.
2.3 Identifikasi Foto
Foto-foto yang dikumpulkan untuk tujuan identifikasi foto diproses mengikuti metodologi yang dijelaskan oleh Haughey et al. ( 2020 ) dan disortir, dikelola, dan disimpan menggunakan perangkat lunak manajemen data identifikasi foto DISCOVERY (Gailey dan Karczmarski 2012 ). Foto-foto diperiksa dan diberi skor menurut kriteria kualitas tertentu dan protokol kekhasan. Pertama, gambar diberi skor untuk kualitas (yaitu, fokus, kejelasan, kontras antara sirip punggung dan latar belakang, sudut relatif terhadap hewan, area sirip punggung yang terlihat, dan ukuran sirip punggung relatif terhadap bingkai) dan diklasifikasikan menjadi tiga tingkatan (yaitu, sangat baik, baik, dan buruk). Kemudian, kekhasan setiap sirip punggung dinilai berdasarkan jumlah fitur yang terlihat pada sirip dan dikategorikan ke dalam tiga kategori: D1, sirip yang sangat khas (misalnya, bagian atas hilang, takik besar, kehilangan pigmentasi, hewan mudah dikenali); D2, sirip yang cukup khas (satu ciri khas [misalnya, takik], atau beberapa torehan dan takik yang lebih kecil, atau bekas luka yang cukup besar); dan D3, sirip yang sedikit khas (tidak ada ciri khas, sirip pada dasarnya bersih kecuali bekas luka kecil, gelombang kecil, atau torehan kecil). Jika sirip punggung tidak memiliki ciri khas apa pun, maka dikategorikan sebagai “bersih”. Untuk mengembangkan riwayat penangkapan individu yang teridentifikasi melalui foto, hanya gambar individu dewasa yang khas (D1 dan D2) yang dianggap berkualitas sangat baik atau baik yang digunakan. Ikan muda dan anak ikan tidak dimasukkan dalam analisis karena tidak memiliki tanda khas.
2.4 Analisis Data
2.4.1 Mendefinisikan Asosiasi dan Memperkirakan Indeks Afiliasi
Analisis asosiasi dilakukan menggunakan SOCPROG v2.9 (Whitehead 2009 ). Semua lumba-lumba dewasa yang terlihat dan teridentifikasi melalui foto dalam kelompok yang sama dianggap terkait (Whitehead 2008a ). Untuk mencegah bias dalam indeks asosiasi, hanya kelompok dengan 50% atau lebih individu yang teridentifikasi yang disertakan (Parra et al. 2011 ). Analisis asosiasi dibatasi pada penampakan pertama per hari dari setiap lumba-lumba individu dalam suatu kelompok. Selain itu, hanya individu dewasa yang terlihat minimal lima kali selama seluruh periode studi yang disertakan dalam analisis. Ambang penampakan ini dipilih untuk memastikan keakuratan dalam indeks asosiasi, karena memasukkan individu dengan penampakan yang lebih sedikit dapat menyebabkan ketidakakuratan (Bejder et al. 1998 ). Ambang lima penampakan selaras dengan praktik dalam penelitian lain tentang struktur sosial lumba-lumba, yang memungkinkan adanya perbandingan lintas penelitian (Chilvers dan Corkeron 2002 ; Félix 1997 ; Wells et al. 1987 ; Zanardo et al. 2016 ). Ambang ini juga mencerminkan kendala lapangan praktis, yang menyeimbangkan kebutuhan akan data yang andal dengan durasi dan tingkat penangkapan kembali yang umum dalam penelitian tersebut (Methion dan Díaz López 2019 ). Lebih jauh lagi, lima penampakan adalah jumlah minimum yang diperlukan untuk menghitung tumpang tindih wilayah jelajah untuk digunakan dalam indeks afiliasi umum (generalized affiliation indexes/GAIs) (Hunt et al. 2019 ; Whitehead 2008a ). Hal ini konsisten dengan metodologi yang digunakan dalam beberapa penelitian lain (misalnya, Elliser dan Herzing 2014 ; Hartman et al. 2008 ; Hunt et al. 2019 ; Parra et al. 2011 ). Mengingat ambang batas penampakan ini, analisis kami difokuskan pada individu yang kemungkinan merupakan penduduk NWC.
Keandalan data untuk mendeteksi kompleksitas sosial yang sebenarnya dinilai menggunakan koefisien variasi (CV) diferensiasi sosial.
dan korelasi antara indeks asosiasi sebenarnya dan estimasi
(Whitehead 2008a ).
menghitung seberapa banyak variasi yang ada dalam data asosiasi dalam populasi. Nilai
~ 0,2 menunjukkan bahwa populasi berdiferensiasi buruk, nilai
> 0,5 menunjukkan sistem sosial yang terdiferensiasi dengan baik, dan nilai
> ~2 menunjukkan masyarakat yang sangat terdiferensiasi, di mana kekuatan asosiasi antar individu bervariasi (Whitehead 2009 ). Koefisien korelasi
menunjukkan jumlah daya yang dimiliki data untuk mendeteksi struktur sosial yang sebenarnya, dengan nilai mendekati 1 menunjukkan representasi yang baik dan nilai mendekati 0,4 menunjukkan representasi sedang (Whitehead 2008a , 2008b ).
Indeks asosiasi setengah bobot (HWI) digunakan untuk memperkirakan proporsi waktu yang dihabiskan oleh dua individu untuk berasosiasi, sambil memperhitungkan observasi yang hilang, seperti ketika individu relatif lebih mungkin terdeteksi ketika mereka terpisah daripada ketika mereka bersama (Cairns dan Schwager 1987 ; Godde et al. 2013 ). Nilai HWI berkisar dari 0 (individu tidak pernah terlihat bersama) hingga 1 (mereka diharapkan selalu terlihat bersama). GAI digunakan bersama dengan HWI untuk memperhitungkan efek faktor struktural yang dapat memengaruhi asosiasi sejati dan mengarah pada indeks asosiasi yang bias (Whitehead dan James 2015 ). Pengendalian terhadap tumpang tindih wilayah jelajah sangat penting karena wilayah jelajah yang tumpang tindih dapat mengarah pada asosiasi nyata yang semata-mata dihasilkan dari kedekatan spasial daripada ikatan sosial sejati. Demikian pula, memperhitungkan sifat suka berteman penting karena individu bervariasi dalam kecenderungan keseluruhan mereka untuk berasosiasi dengan orang lain. Mereka yang secara alami lebih suka berkelompok akan tampak memiliki lebih banyak koneksi sosial, yang dapat meningkatkan ukuran asosiasi sosial dan mengaburkan hubungan preferensial yang sebenarnya. GAI dihitung sebagai residual dari model linier umum, di mana indeks asosiasi setengah bobot adalah variabel dependen, dan variabel prediktor struktural yang disertakan adalah sifat suka berkelompok dan tumpang tindih wilayah jelajah (Hunt et al. 2019 ; Whitehead dan James 2015 ).
Gregariousness adalah ukuran kecenderungan satu individu untuk berasosiasi dengan individu lain dan diperkirakan menggunakan SOCPROG sebagai log dari jumlah indeks asosiasi yang melibatkan satu individu dikalikan dengan jumlah indeks yang melibatkan individu lain (Godde et al. 2013 ). Area tumpang tindih home range spasial dihitung menggunakan paket “AdehabitatHR” (Calenge 2006 ) dalam R v 4.0.4 (Tim Inti Pengembangan R 2008 ) mengikuti metode distribusi pemanfaatan (UD) berbasis kernel. UD adalah fungsi bivariat yang memberikan kepadatan probabilitas bahwa seorang individu ditemukan di suatu titik menurut koordinat geografisnya. Menggunakan metode ini memungkinkan seseorang untuk memperkirakan tumpang tindih spasial antara UD dari dua individu (berdasarkan UD 95%). Untuk memperhitungkan keberadaan NWC, kami mendefinisikannya sebagai penghalang yang tidak dapat dilintasi oleh hewan dan menyesuaikan parameter penghalusan, h , untuk memastikan tumpang tindih minimal atau tidak ada di darat dengan mengikuti metode yang dijelaskan oleh Benhamou dan Cornélis ( 2010 ). Kami menggunakan ArcMap v10.8 untuk menilai secara visual setiap wilayah jelajah per individu untuk nilai h yang berbeda , dengan nilai h 1000 ditemukan sebagai yang paling sesuai. Korelasi antara variabel prediktor struktural dan indeks asosiasi HWI diuji menggunakan prosedur penugasan kuadrat regresi berganda (MRQAP) dalam SOCPROG. Variabel prediktor struktural dengan korelasi yang tidak signifikan ( p < 0,05) dihilangkan dari perhitungan GAI (Whitehead dan James 2015 ).
Rata-rata GAI dan simpangan baku (SD) dari afiliasi yang diamati diperkirakan untuk semua individu dewasa yang melihat lima kali atau lebih selama seluruh periode studi. Kami kemudian menggunakan uji permutasi untuk menilai apakah individu berasosiasi secara acak atau memiliki afiliasi pilihan/penghindaran ( p < 0,05) (Whitehead dan James 2015 ). Permutasi dilakukan dalam periode pengambilan sampel harian untuk memperhitungkan efek demografis (misalnya, kelahiran, kematian, dan migrasi) (Whitehead 2008a ). Jumlah permutasi ditingkatkan melalui beberapa pengujian (Bejder et al. 1998 ) hingga stabil, yang terjadi pada 1000 permutasi. Individu dengan persahabatan pilihan selama beberapa studi pengambilan sampel diharapkan memiliki simpangan baku yang jauh lebih tinggi ( p < 0,05) daripada data acak (Whitehead 2009 ). Pasangan yang dihindari dianggap sebagai diade yang kurang terkait dari yang diharapkan.
2.4.2 Analisis Jaringan Sosial
Asosiasi individu divisualisasikan dalam diagram jaringan sosial menggunakan perangkat lunak NETDRAW v.2.179 dan metode spring embedding (Borgatti et al. 2002 ). Individu diidentifikasi dengan node dan label identitas. Jaringan dibuat dengan nilai asosiasi GAI sebagai tautan antara individu dan menggunakan residual dari model linear umum, di mana tautan bisa positif dan negatif (Whitehead dan James 2015 ). NETDRAW menggunakan algoritma yang diarahkan oleh gaya untuk mengatur node. Algoritma ini memposisikan node dengan asosiasi yang lebih kuat lebih dekat satu sama lain untuk kejelasan visual tetapi tidak mencerminkan ada atau tidaknya klaster sosial yang sebenarnya. Asosiasi antara individu diselidiki lebih lanjut untuk melihat apakah persahabatan dalam populasi lebih disukai, kasual, atau dihindari dengan mengubah residual mentah GAI menjadi residual deviasi. Nilai yang lebih besar dari 2 dianggap sebagai persahabatan yang disukai; skor di bawah -2 dianggap sebagai persahabatan yang dihindari, dan skor antara -2 dan 2 dianggap sebagai persahabatan biasa (Whitehead 2009 ; Whitehead dan James 2015 ). Tiga metrik jaringan sosial dihitung untuk setiap individu lumba-lumba untuk memberikan gambaran umum yang komprehensif dari struktur sosial mereka: kekuatan, koefisien pengelompokan, dan afinitas. Kekuatan adalah jumlah GAI untuk semua individu yang terhubung dengan individu tertentu (Whitehead 2009 ), dengan kekuatan yang tinggi menunjukkan bahwa individu tersebut terhubung dengan baik dengan individu lain (Whitehead 2009 ). Koefisien pengelompokan mengukur seberapa terhubung rekan-rekan individu di antara mereka sendiri dan dengan demikian mencerminkan tingkat struktur dalam suatu masyarakat (Holme et al. 2007 ). Afinitas adalah ukuran kekuatan rekan-rekan individu, yang dibobot dengan indeks asosiasi di antara mereka, di mana individu dengan afinitas tinggi memiliki asosiasi yang relatif tinggi dengan individu yang memiliki kekuatan tinggi (Whitehead 2009 ). Kekuatan dan afinitas dipilih untuk mengukur keterhubungan keseluruhan individu dalam populasi, yang mencerminkan tingkat interaksi sosial mereka. Koefisien pengelompokan disertakan untuk menilai tingkat pengelompokan lokal dalam jaringan, yang dapat menunjukkan keberadaan subkelompok. Untuk menguji signifikansi metrik jaringan sosial yang diamati, nilai acak dihasilkan menggunakan replikasi bootstrap data dengan mengambil sampel ulang dan mengubah posisi data sebanyak 1000 kali (Lusseau et al. 2006 ).
2.4.3 Pengelompokan Sosial Kami menggunakan modularitas
Newman metode yang diterapkan dalam SOCPROG v2.9 untuk menilai apakah ada kelompok sosial dalam populasi (yaitu, kelompok individu dengan asosiasi yang lebih tinggi dengan anggota lain dalam kelompok yang sama daripada yang diharapkan secara kebetulan). Metode berbasis vektor eigen bertujuan untuk memaksimalkan untuk mendeteksi komunitas yang berbeda dan mengungkapkan kelompok sosial (Newman 2006 ). Nilai-nilai > 0,3 menunjukkan pembagian jaringan sosial yang baik ke dalam kelompok-kelompok berbeda dalam populasi (Newman 2004 ).
2.4.4 Variabilitas Temporal dalam Pola Asosiasi
Kami memperkirakan tingkat asosiasi yang tertinggal dan membandingkannya dengan tingkat asosiasi nol untuk menyelidiki variabilitas temporal dalam pola asosiasi (Whitehead 1995 ). Tingkat asosiasi yang tertinggal adalah perkiraan probabilitas bahwa, jika dua individu berasosiasi pada waktu tertentu, mereka akan tetap berasosiasi dengan berbagai jeda waktu di kemudian hari (Whitehead 2008a ). Tingkat asosiasi nol adalah nilai yang diharapkan dari tingkat asosiasi yang tertinggal jika tidak ada asosiasi yang disukai (Whitehead 1995 ).
Untuk analisis ini, kami membakukan tingkat asosiasi tertinggal (SLAR) dan tingkat asosiasi nol, karena tidak semua individu dalam kelompok diidentifikasi. SLAR adalah estimasi probabilitas bahwa jika dua individu diasosiasikan pada suatu waktu, individu kedua adalah rekan yang dipilih secara acak dari yang pertama (Whitehead 2008a ). Metode temporal jackknife digunakan untuk memperoleh kesalahan standar untuk setiap SLAR (Efron dan Stein 1981 ). Untuk menghitung SLAR secara efektif dalam musim lapangan, sambil menghilangkan pertimbangan tingkat asosiasi antara periode survei tahunan, kami menetapkan jeda waktu maksimum pada 199 hari (yaitu, periode terpanjang antara asosiasi pertama dan asosiasi terakhir dari setiap pasangan individu dalam setiap periode survei tahunan).
Setelah SLAR dan tingkat asosiasi nol diperoleh, mereka kemudian dibandingkan dengan empat model asosiasi temporal yang berbeda dan dievaluasi untuk pemilihan model (Whitehead 1995 ). Model-model ini meliputi: (1) persahabatan pilihan, pasangan individu yang memiliki preferensi untuk berasosiasi terus-menerus dari waktu ke waktu; (2) kenalan biasa, asosiasi tidak teratur yang berlanjut dari waktu ke waktu; (3) persahabatan pilihan dan kenalan biasa, asosiasi antara individu dapat bertahan dari waktu ke waktu pada dua tingkat: pilihan dan biasa; dan (4) dua tingkat kenalan biasa, pasangan individu tidak berasosiasi dari waktu ke waktu, tetapi pada dua tingkat yang berbeda (Whitehead 1995 ). Pemilihan model didasarkan pada Kriteria Informasi Kuasi-Akaike (QAIC), di mana nilai QAIC terendah mewakili model dengan dukungan paling banyak (Whitehead 2007 ). Model yang berbeda dengan 0–2 QAIC (
QAIC) dianggap didukung secara substansial (Burnham dan Anderson 2002 ).
3 Hasil
3.1 Upaya Survei
Total 294 hari survei diselesaikan antara Mei 2013 dan Oktober 2021. Dari 344 lumba-lumba dewasa yang diidentifikasi, 116 terlihat
5 kali dan selanjutnya dipertimbangkan untuk analisis asosiasi. Total penampakan per individu berkisar antara 5 hingga 38 (rata-rata
SD = 11,4
9,8, median = 9,5).
3.2 Pola Asosiasi
Korelasi antara indeks asosiasi sebenarnya dan estimasi
, dan CV dari indeks asosiasi sebenarnya
menunjukkan bahwa kumpulan data tersebut memiliki daya yang baik untuk mendeteksi sistem sosial yang sebenarnya, yang mencerminkan masyarakat yang sangat terdiferensiasi dengan hubungan yang bervariasi di antara individu. HWI memiliki nilai rata-rata 0,056 (
) dan CV sebesar 1,955, yang mencerminkan variabilitas tinggi dalam kekuatan asosiasi di antara individu. Rata-rata ukuran wilayah jelajah lumba-lumba adalah 248,57 km 2 , dengan rata-rata tumpang tindih sebesar 10,1% (83,71 km 2 ). Tumpang tindih sangat bervariasi di antara individu, berkisar dari minimum 0 km 2 hingga maksimum 821,52 km 2 . Uji MRQAP mengungkapkan korelasi signifikan antara indeks asosiasi gregariousness ( r parsial = 0,1266, p < 0,001) dan tumpang tindih wilayah jelajah ( r parsial = 0,2956, p < 0,001), dan oleh karena itu ini dimasukkan dalam analisis GAI.
Rata-rata GAI di antara semua individu adalah 0,00
, dengan rata-rata maksimum 0,48
Nilai GAI minimum dan maksimum masing-masing adalah -0,26 dan 0,87. Ada indikasi kuat afiliasi nonacak dalam populasi, karena simpangan baku GAI (SD yang diamati = 0,101) secara signifikan lebih besar daripada data yang diubah (SD acak ≤ 0,001, p = 0,001).
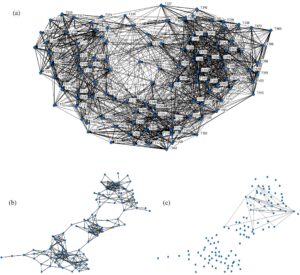
Hubungan asosiasi GAI antara lumba-lumba hidung botol Indo-Pasifik adalah 70% negatif, yang menunjukkan “hubungan longgar” di antara sebagian besar individu. Sisa hubungan menunjukkan bahwa 25% dari lumba-lumba individu dalam populasi memiliki “hubungan stabil” (Gambar 2a ), dan 5% individu tidak menunjukkan hubungan. Setelah mengubah GAI mentah menjadi residual deviasi, 357 afiliasi pilihan, 5907 afiliasi kasual, dan 22 afiliasi yang dihindari diidentifikasi (Gambar 2b,c ).
Perbandingan metrik jaringan sosial yang diamati dan acak menunjukkan bahwa kekuatan keseluruhan, koefisien pengelompokan, dan afinitas jaringan sosial lumba-lumba hidung botol Indo-Pasifik tidak berbeda dari jaringan acak (Tabel 1 ). Indeks modularitas Newman (
) menunjukkan jaringan sosial yang terhubung dengan baik tanpa bukti adanya kelompok sosial. Hasil ini menunjukkan bahwa, meskipun asosiasi individual dapat bervariasi, struktur jaringan secara keseluruhan tidak menunjukkan penyimpangan yang signifikan dari ekspektasi acak dalam hal metrik jaringan secara keseluruhan.
| Metrik jaringan sosial | Rata-rata ± SD | Acak ± SD | P |
|---|---|---|---|
| Kekuatan | 0,00 ± 1,77 | 0,00 ± 0,24 | > 0,05 |
| Koefisien pengelompokan | 2,81 ± 26,61 | -1,93 ± 11,93 | > 0,05 |
| Afinitas | -1,27 ± 29,75 | -0,81 ± 45,35 | > 0,05 |
Catatan: Statistik jaringan dihitung menggunakan residual mentah GAI, yang dirata-ratakan oleh semua individu. Perbedaan signifikan ( p < 0,05) dari nilai rata-rata acak jaringan (nilai yang diharapkan jika individu tidak memiliki preferensi asosiasi) dievaluasi menggunakan 1000 permutasi.
3.3 Pola Asosiasi Temporal
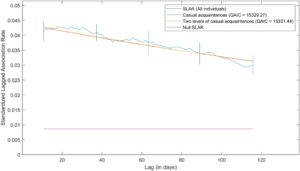
SLAR lumba-lumba hidung botol Indo-Pasifik tetap lebih tinggi daripada tingkat asosiasi nol di seluruh periode studi (Gambar 3 ), yang menunjukkan bahwa asosiasi di antara individu dari waktu ke waktu bersifat nonacak. Ada penurunan yang stabil dalam durasi tingkat asosiasi yang tertinggal. Secara keseluruhan, dua model yang paling sesuai yang menggambarkan pola asosiasi temporal lumba-lumba hidung botol Indo-Pasifik adalah kenalan biasa dan dua tingkat kenalan biasa (Tabel 2 ). Oleh karena itu, pola asosiasi lumba-lumba hidung botol Indo-Pasifik di NWC tampaknya dicirikan terutama oleh asosiasi yang longgar dan jangka pendek yang hanya berlangsung dalam waktu singkat (hari).
| Model | Persamaan model | QAIC | ∆QAIC |
|---|---|---|---|
| Persahabatan pilihan + akuisisi kasual | a2 + a3*eksp.(−a1*td) | 15.358,05 | 28.78 |
| Persahabatan yang disukai | sebuah 1 | 15.354,28 | Tanggal 25.01 |
| Dua tingkat akuisisi kasual | a3*exp(−a1*td) + a4*exp(−a2*td) | 15.331,44 | 2.17 |
| Pembelian kasual | a2*eksp(−a1*td) | 15.329,27 | 0.0 |
Catatan: Model yang dianggap memiliki dukungan terbanyak ditandai dengan huruf tebal. Dalam persamaan model, koefisien mewakili hal yang berbeda: “a1” adalah persahabatan pilihan; “a2” adalah kenalan biasa; “a3” adalah persahabatan pilihan dan kenalan biasa yang digabungkan; “a4” melibatkan dua tingkat kenalan biasa; “exp” adalah singkatan dari fungsi eksponensial; dan “td” adalah waktu tunda, yang ditetapkan pada 199 hari.
4 Diskusi
Sedikit yang diketahui tentang struktur sosial lumba-lumba hidung botol Indo-Pasifik yang menghuni perairan pesisir NWC di Australia Barat. Dalam penelitian ini, kami menggunakan data identifikasi foto, GAI, dan metrik jaringan sosial untuk menyelidiki kekuatan asosiasi, pengelompokan sosial, dan stabilitas temporal di antara lumba-lumba hidung botol Indo-Pasifik dewasa di NWC. Hasil kami menunjukkan bahwa pola asosiasi mereka dicirikan oleh asosiasi jangka pendek yang tidak acak dan longgar yang hanya berlangsung beberapa hari tanpa bukti pengelompokan sosial seperti yang ditunjukkan oleh indeks modularitas yang rendah.
Pola asosiasi ini konsisten dengan temuan dari studi lain tentang lumba-lumba hidung botol Indo-Pasifik di New South Wales utara (Hawkins dan Gartside 2008 ), lumba-lumba hidung botol biasa ( Tursiops truncatus ) di Bahamas (Rogers et al. 2004 ), lumba-lumba hidung botol ( Tursiops spp.) di Selandia Baru (Lusseau et al. 2003 ), dan lumba-lumba hidung botol biasa di Skotlandia (Lusseau et al. 2006 ). Dinamika fisi-fusi seperti itu merupakan ciri khas lumba-lumba hidung botol dan umumnya terkait dengan variabel ekologi atau demografi. Temuan ini memberikan informasi dasar tentang struktur sosial dan dinamika fisi-fusi lumba-lumba hidung botol Indo-Pasifik di NWC dan berfungsi sebagai landasan untuk studi masa depan tentang peran dan pengaruh faktor genetik, ekologi, lingkungan, dan antropogenik pada sosialitas mereka.
Metrik jaringan sosial untuk lumba-lumba di NWC menunjukkan bahwa populasi menunjukkan tingkat penataan sosial yang relatif rendah, karena tidak ada metrik yang diamati yang berbeda secara signifikan dari jaringan acak. Kekuatan rata-rata yang diamati (0,00 ± 1,77) tidak berbeda secara signifikan dari rata-rata acak, yang menunjukkan bahwa pola asosiasi lumba-lumba sebagian besar dicirikan oleh hubungan diadis acak yang didorong oleh tumpang tindih spasial-temporal dan sifat suka berteman berpasangan daripada menunjukkan diferensiasi sosial yang terstruktur. Demikian pula, afinitas juga tidak berbeda secara signifikan dari acak (−1,27 ± 29,75), yang menunjukkan bahwa kekuatan koneksi mereka longgar daripada terstruktur. Koefisien pengelompokan (2,81 ± 26,61) juga tidak berbeda secara signifikan dari nilai acak, yang menunjukkan bahwa ada pengelompokan lokal minimal, dan kelompok sosial tidak membentuk kelompok kohesif yang berbeda. Hasil-hasil ini menunjukkan bahwa sistem sosial lumba-lumba hidung botol Indo-Pasifik di NWC secara keseluruhan sangat cair dan dinamis, dengan asosiasi yang sebagian besar dipengaruhi oleh faktor spasial-temporal daripada struktur sosial yang kuat.
Penggunaan GAI mengungkapkan bahwa afiliasi di antara lumba-lumba hidung botol di NWC dipengaruhi oleh sifat suka berteman dan tumpang tindih wilayah jelajah spasial, yang menyoroti pentingnya mengendalikan pengaruh variabel sosial dalam analisis struktur sosial. Proporsi tinggi hubungan asosiasi GAI negatif (70%) menunjukkan bahwa sebagian besar lumba-lumba memiliki asosiasi yang longgar atau kasual, daripada membentuk ikatan yang kuat dan stabil. Ini menunjukkan bahwa interaksi di antara lumba-lumba lebih oportunistik dan cenderung tidak didasarkan pada hubungan jangka panjang. Hasil pada tumpang tindih wilayah jelajah lebih jauh menjelaskan dinamika ini. Lumba-lumba dengan wilayah jelajah yang tumpang tindih menunjukkan tingkat asosiasi yang lebih tinggi, yang menunjukkan bahwa kedekatan spasial memfasilitasi interaksi sosial. Korelasi signifikan antara tumpang tindih wilayah jelajah dan indeks asosiasi menunjukkan bahwa kedekatan spasial merupakan faktor kunci dalam membentuk interaksi sosial di NWC. Pola ini telah ditemukan di wilayah lain, seperti Sarasota Bay, Florida, di mana tumpang tindih wilayah jelajah telah dikaitkan dengan peningkatan afiliasi sosial (Wells 2014 ). Di NWC, interaksi antara penggunaan habitat dan asosiasi sosial kemungkinan mencerminkan strategi untuk mengoptimalkan efisiensi mencari makan dan menghindari predator.
Seperti yang diharapkan, lumba-lumba hidung botol Indo-Pasifik NWC hidup dalam masyarakat fisi–fusi, yang mungkin mencerminkan adaptasi terhadap variabel ekologi dan sosial. Dalam masyarakat yang hidup berkelompok, keputusan untuk bergabung atau memisahkan diri dari kelompok dianggap sebagian besar didorong oleh trade-off antara risiko predasi dan biaya kompetisi makan (Connor dan Krützen 2015 ; Gowans et al. 2007 ; van Schaik 1999 ). Kelompok besar dapat mengurangi risiko predasi tetapi meningkatkan kompetisi untuk sumber daya. Di Shark Bay, predator seperti hiu macan ( Galeocerdo cuvier ) memengaruhi ukuran kelompok lumba-lumba dan penggunaan habitat (Heithaus 2001 ; Heithaus dan Dill 2002 , 2006 ). Lumba-lumba menyesuaikan ukuran kelompok berdasarkan risiko predasi dan ketersediaan makanan, membentuk kelompok yang lebih besar ketika risikonya tinggi dan sumber daya berlimpah, dan kelompok yang lebih kecil ketika sumber daya langka (Gowans et al. 2007 ). Meskipun pola ini diamati pada beberapa populasi, ketersediaan sumber daya tidak secara universal menentukan perilaku sosial, karena variasi individu dan perbedaan habitat dapat memengaruhi strategi sosial (Fisher dan Cheney 2023 ; Levengood et al. 2022 ). Studi masa depan tentang ketersediaan mangsa dan risiko predasi akan membantu memperjelas dampaknya pada struktur sosial lumba-lumba hidung botol Indo-Pasifik NWC.
Keterkaitan genetik (kekerabatan) merupakan faktor tambahan yang dapat memengaruhi hubungan sosial baik jantan maupun betina, dengan ikatan sosial yang lebih disukai terbentuk di antara kerabat dengan tingkat keterkaitan genetik yang tinggi (Smith 2014 ). Kekerabatan telah terbukti menjadi faktor penting dalam pengembangan dan pemeliharaan ikatan sosial pada banyak mamalia, termasuk lumba-lumba hidung botol (Chabanne et al. 2022 ; Connor dan Krützen 2015 ; Diaz-Aguirre et al. 2018 , 2019 , 2020 ; Foroughirad et al. 2023 ; Frère et al. 2010 ; Gerber et al. 2019 ; Marfurt et al. 2022 ; Möller 2012 ; Smolker et al. 1992 ). Di Shark Bay, lumba-lumba hidung botol betina membentuk asosiasi pilihan dengan betina lain meskipun sedikit tumpang tindih wilayah jelajah, sementara beberapa betina menghindari satu sama lain meskipun tumpang tindih penuh. Asosiasi pilihan ini lebih umum di antara betina yang lebih dekat hubungan biparental daripada yang diharapkan secara kebetulan (Frère et al. 2010 ). Sebaliknya, lumba-lumba jantan lebih menyukai ikatan sosial dengan teman sebaya daripada kerabat dan berasosiasi dengan jantan lain di awal kehidupan, yang memperkirakan pasangan aliansi dewasa (Gerber et al. 2019 ). Hubungan jenis kelamin dan genetik di antara individu-individu dalam populasi ini tidak diketahui. Investigasi di masa depan, yang melibatkan teknik molekuler, diperlukan untuk menilai pengaruh kekerabatan dan hubungan khusus jenis kelamin pada pola pengelompokan dan asosiasi lumba-lumba hidung botol Indo-Pasifik di lepas pantai NWC.
Variabilitas temporal yang diamati dalam pola asosiasi di antara lumba-lumba hidung botol Indo-Pasifik terutama mencerminkan asosiasi yang longgar dan jangka pendek. Pergeseran demografis, seperti kedatangan individu baru atau perubahan komposisi kelompok, dapat memengaruhi pola-pola ini dengan memperkenalkan dinamika baru dan mengubah ikatan sosial yang ada. Pergerakan lumba-lumba ke dalam dan ke luar area studi menyebabkan proporsi individu sementara yang signifikan (Haughey et al. 2020 , 2021 ). Jenis masyarakat terbuka dan sementara ini mirip dengan lumba-lumba hidung botol biasa di kepulauan samudra Madeira, Portugal, di mana komposisi kelompok tetap cair dengan perekrutan individu baru secara teratur (Dinis et al. 2018 ). Variabilitas demografis tersebut dapat berkontribusi pada kurangnya pengelompokan sosial yang diamati dan menggarisbawahi sifat dinamis masyarakat lumba-lumba.
Struktur sosial yang dijelaskan dalam studi ini sebagian besar mencerminkan lumba-lumba penghuni karena pengecualian sebagian besar populasi dengan ukuran sampel yang rendah. Dari 344 individu yang teridentifikasi, hanya 116 lumba-lumba yang terlihat lima kali atau lebih yang disertakan dalam analisis. Pengecualian ini kemungkinan kurang mewakili individu yang berpindah tempat, yang mungkin memiliki lebih sedikit penampakan, dan akibatnya, lebih sedikit data yang tersedia untuk analisis asosiasi. Oleh karena itu, meskipun temuan tersebut memberikan wawasan berharga tentang dinamika sosial populasi penghuni, temuan tersebut mungkin tidak sepenuhnya menangkap interaksi dan afiliasi lumba-lumba yang berpindah tempat di area studi. Keterbatasan ini menunjukkan bahwa dinamika sosial keseluruhan semua lumba-lumba di NWC mungkin lebih kompleks dan bervariasi daripada yang dilaporkan, sehingga memerlukan kehati-hatian saat menggeneralisasi hasil ini ke seluruh populasi. Studi masa depan yang menggabungkan kumpulan data yang lebih luas, termasuk individu yang berpindah tempat, akan sangat penting untuk pemahaman yang lebih komprehensif tentang struktur sosial lumba-lumba hidung botol Indo-Pasifik di wilayah ini.
Sebagai kesimpulan, penelitian ini memungkinkan kita untuk memperoleh pemahaman dasar tentang dinamika sosial lumba-lumba hidung botol Indo-Pasifik di NWC. Hasil penelitian mengungkapkan bahwa komunitas ini hidup dalam masyarakat fisi-fusi terbuka, dengan hubungan diadik non-acak dan asosiasi fluida jangka pendek. Kemampuan beradaptasi dalam struktur sosial ini diyakini mendukung keuntungan adaptif, yang memungkinkan lumba-lumba untuk merespons faktor ekologi dan perubahan lingkungan, sehingga memengaruhi struktur komunitas dan dinamika ekosistem. Penelitian mendatang yang melibatkan risiko predasi, variabilitas spasiotemporal dalam sumber daya mangsa, teknik molekuler, dan preferensi habitat akan membantu kita lebih memahami sosioekologi populasi ini.


