
ABSTRAK
Bahasa Indonesia: Menjelajahi respons evolusioner dan fisiologis terhadap stres lingkungan sangat penting untuk menilai dampak perubahan iklim pada populasi liar. Kerang, penghuni utama benthos dengan nilai ekologis dan ekonomi yang tinggi, adalah spesies yang sangat rentan yang mungkin didorong ke batas ekologis mereka karena pemanasan mengancam kelangsungan hidup dan stabilitas populasi mereka. Spesies dalam kompleks Mytilus edulis umumnya ditemukan di wilayah beriklim sedang secara global; di Laut Baltik, populasi dibentuk oleh hibrida M. edulis dan M. trossulus dengan tingkat introgresi M. galloprovincialis yang rendah . Studi ini menyelidiki mekanisme yang melaluinya ketahanan terhadap pemanasan global dapat dilacak dengan cepat pada kerang Baltik (Kiel, Jerman). Untuk ini, kami mempelajari dua kelompok kerang muda (hewan yang baru saja menetap), satu yang terpapar pada peristiwa panas ekstrem di awal kehidupan dan satu yang naif terhadap stresor ini. Kedua kelompok kemudian diekspos ke suhu eksperimen berkisar antara 21°C hingga 26°C, dengan kinerja hewan diukur setelah 25 hari. Dampak stres termal pada komposisi genetik setiap kelompok kemudian dinilai dengan membuat genotipe 50 individu menggunakan array SNP 60 K kerang biru. Kami mengamati peningkatan signifikan dalam genotipe M. edulis bersamaan dengan penurunan M. trossulus dalam kelompok yang ditantang, dibandingkan dengan juvenil naif. Kami juga menemukan paparan suhu tinggi memengaruhi kinerja kelompok kerang, mengurangi berat jaringan kering individu yang dipilih. Hasil dari penelitian ini memberikan wawasan tentang bagaimana seleksi melalui stres termal memengaruhi kinerja dan komposisi genetik spesies pasang surut yang didistribusikan secara global, dengan implikasi penting untuk memahami dan mengelola populasi kerang dalam skenario pemanasan di masa mendatang.
1 Pendahuluan
Perubahan iklim yang sedang berlangsung dengan cepat mengubah pola abiotik dan biotik di lautan, yang memengaruhi kebugaran organisme laut dan fungsi ekosistem laut di seluruh dunia (IPCC 2022 ). Untuk mengatasi perubahan lingkungan yang cepat ini, organisme laut dapat bermigrasi ke habitat yang lebih baik atau menunjukkan plastisitas fisiologis yang cepat; namun, mereka pada akhirnya memerlukan beberapa tingkat adaptasi genetik cepat yang diperlukan untuk memastikan persistensi populasi jangka panjang—terutama ketika migrasi bukanlah pilihan yang layak atau sebagai respons terhadap peristiwa ekstrem. Karena peristiwa ekstrem ini semakin melampaui batas banyak spesies, memahami bagaimana, dan sejauh mana, spesies dapat berevolusi untuk menoleransi perubahan ini, serta konsekuensi ekologis yang lebih luas, sangat penting untuk memprediksi bagaimana keanekaragaman hayati laut, distribusi spesies, dan keseluruhan struktur ekosistem dapat dibentuk kembali di masa depan (Radchuk et al. 2019 ).
Meskipun ada bukti bahwa adaptasi dapat terjadi dalam waktu sesingkat satu generasi (Brennan et al. 2019 ; Griffiths et al. 2021 ; Campbell-Staton et al. 2021 ), kita masih memiliki pemahaman yang relatif terbatas tentang bagaimana peristiwa selektif ini bertindak pada tingkat genetik dan konsekuensi fenotipiknya. Studi yang menggabungkan fisiologi eksperimental, genomik, dan evolusi eksperimental dapat memberikan wawasan utama tentang mekanisme yang mendasari seleksi cepat terhadap kondisi ekstrem (Stillman 2019 ). Akuisisi data empiris baru yang mengeksplorasi adaptasi cepat pada spesies non-model dan populasi liar yang mungkin mengalami pergeseran ini sangat penting untuk memperluas pemahaman kita tentang proses ini dan kapasitas untuk adaptasi cepat di seluruh organisme yang beragam.
Kerang biru adalah model ideal untuk memahami dampak peristiwa selektif akut yang ekstrem. Kerang ini adalah spesies dasar benthos, yang menyediakan layanan ekosistem seperti siklus nutrisi dan peningkatan kompleksitas habitat melalui peningkatan struktur spasial untuk spesies terkait (Johannesson et al. 2011 ; van der Schatte Olivier et al. 2020 ), dan merupakan sumber daya akuakultur dan perikanan yang penting (Avdelas et al. 2021 ). Tiga spesies dalam kompleks kerang biru Mytilus yang berasal dari Atlantik utara, M. galloprovincialis , M. edulis , dan M. trossulus , melakukan hibridisasi di area distribusi geografis yang tumpang tindih (Fraïsse et al. 2016 ). Di Laut Baltik, populasi kerang biru merupakan kawanan yang sangat introgres yang terdiri dari ketiga spesies (Vendrami et al. 2020 ). Di wilayah ini, kerang biru beradaptasi secara lokal dengan gradien salinitas alami (Knöbel et al. 2021 ) yang beralih dari sepenuhnya laut di barat menjadi hampir air tawar di bagian timur cekungan (Meier et al. 2006 ), dengan distribusi genotipe yang selaras dengan perubahan bertahap ini. Populasi ini sudah mengatasi tekanan lingkungan, seperti pemanasan, pada tingkat yang sebagian besar wilayah pesisir diperkirakan tidak akan mengalaminya hingga akhir abad ini (Reusch et al. 2018 ). Namun, sedikit yang diketahui mengenai implikasi stres termal pada populasi ini pada tingkat genomik.
Laut Baltik dapat berfungsi sebagai model untuk memahami respons populasi terhadap perubahan global karena mengalami pergeseran lingkungan pada tingkat yang lebih cepat daripada sebagian besar wilayah lain (Reusch et al. 2018 ). Ini adalah salah satu laut yang menghangat paling cepat secara global (Belkin 2009 ) dan suhu permukaan (SST) telah meningkat lebih dari 0,3°C selama beberapa dekade terakhir, dengan tren pemanasan yang terutama terlihat selama bulan-bulan musim panas dibandingkan dengan musim dingin (Meier et al. 2022 ). Misalnya, pada tahun 2018, musim panas terhangat sejak 1990 (Naumann et al. 2018 ), gelombang panas meningkatkan suhu lebih dari 4°C di atas rata-rata 28 tahun (1990–2018) di bagian selatan cekungan (Meier et al. 2022 ). Pergeseran rezim suhu, baik melalui suhu musiman rata-rata yang dimodifikasi atau kejadian ekstrem pendek yang terisolasi, dapat berkontribusi terhadap pergeseran komunitas dan memicu reaksi berantai hilangnya keanekaragaman hayati, mendorong ekosistem ini menuju keadaan alternatif (Johannesson et al. 2011 ; Pansch et al. 2018 ; Reusch et al. 2005 ).
Suhu adalah pengatur utama fisiologi ektoterm (Somero et al. 2017 ). Kerang merespons panas akut dengan meningkatkan metabolismenya, mengurangi proses yang membutuhkan banyak energi (misalnya, pertumbuhan) dan/atau melengkapi dengan metabolisme anaerobik untuk memenuhi permintaan energi yang meningkat (Anestis et al. 2007 ; Braby dan Somero 2006 ; Tagliarolo dan McQuaid 2015 ; Vajedsamiei, Wahl, et al. 2021 ; Zittier et al. 2015 ). Sementara strategi ini dapat menjamin kelangsungan hidup jangka pendek dalam suhu yang tidak optimal, paparan yang berkepanjangan di luar CT maks menyebabkan ketidaksesuaian pasokan-permintaan metabolik, yang mengganggu kinerja dan menyebabkan kematian (Pörtner dan Farrell 2008 ; Ritchie 2018 ; Anestis et al. 2007 ; Feidantsis et al. 2020 ). Faktanya, peristiwa kematian massal telah dilaporkan di hamparan kerang Mediterania setelah gelombang panas laut (Bracchetti et al. 2024 ).
Kemampuan untuk menyesuaikan laju metabolisme sebagai respons terhadap suhu merupakan sifat penting untuk bertahan hidup dan bervariasi di antara spesies kerang (Pörtner dan Farrell 2008 ; Vajedsamiei, Wahl, et al. 2021 ). Misalnya, batas termal atas untuk mempertahankan Cakupan Pertumbuhan positif (yaitu, energi ekstra yang tersedia untuk pertumbuhan setelah memenuhi kebutuhan dasar untuk bertahan hidup) di M. trossulus , M. edulis , dan M. galloprovincialis yang diaklimatisasi dengan suhu musim panas dari pantai Pasifik barat laut dan Atlantik timur laut masing-masing adalah sekitar 17°C, 23°C, dan 30°C (Fly dan Hilbish 2013 ). Hal ini menunjukkan bahwa pada suhu yang tinggi, genotipe M. trossulus memiliki kinerja yang lebih rendah dibandingkan dengan dua spesies lainnya, dengan kemungkinan implikasi untuk pertumbuhan, kelangsungan hidup, dan hasil reproduksi. Sejauh mana genotipe spesies Mytilus yang berbeda dalam populasi hibrida memengaruhi respons terhadap stres termal masih belum jelas.
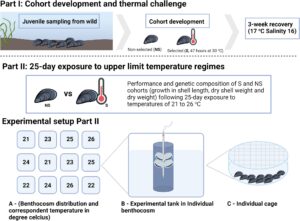
Dalam studi ini, kami mengatasi kesenjangan pengetahuan ini melalui pendekatan eksperimental dua langkah (Gambar 1 ). Pertama, untuk menilai implikasi stres termal pada susunan genetik populasi kerang, kami mengumpulkan spat (yaitu, juvenil yang baru saja menetap) dari satu peristiwa pemukiman di Kiel Fjord dan mengembangkan dua kelompok berbeda, satu dipilih untuk toleransi termal melalui paparan akut tetapi intens terhadap peristiwa stres termal (30 °C selama 47 jam), dan satu naif terhadap stresor ini. Individu dari kedua kelompok tersebut digenotipe dengan SNP-array kerang biru 60 K (Nascimento-Schulze et al. 2023 ), alat genomik yang memungkinkan genotipe individu yang konsisten dan hemat biaya di ribuan lokus yang telah ditentukan sebelumnya, memastikan reproduktifitas analisis (Robledo et al. 2018 ). Kami berhipotesis bahwa pemilihan suhu akan menciptakan kelompok yang berbeda secara genetik, yang mendukung genotipe yang lebih tahan terhadap suhu yang lebih tinggi. Secara khusus, kami memperkirakan bahwa fraksi alel M. trossulus akan menurun setelah seleksi suhu. Kedua, kami menguji dampak stres termal pasca-seleksi pada kinerja kerang. Untuk ini, kami memaparkan individu dari kedua kelompok yang dikembangkan pada skenario suhu musim panas Baltik yang konstan, mulai dari musim panas ekstrem saat ini hingga yang diperkirakan terjadi pada akhir abad ini, selama periode 25 hari, dan menilai kinerja seluruh organisme dalam pertumbuhan cangkang dan pertambahan jaringan kering. Kami berhipotesis bahwa galur yang diseleksi panas akan mengungguli galur yang tidak diseleksi pada suhu yang lebih tinggi.
2 Bahan dan Metode
2.1 Pengembangan Kelompok Remaja Terpilih dan Tidak Terpilih
2.1.1 Acara Seleksi
Sekitar 500.000 spat kerang (yaitu, individu yang menetap) dikumpulkan dengan tangan dari Kiel Fjord subtidal (54.328588, 10.148210, Laut Baltik, Jerman) pada tanggal 28 Juli 2021, dari kedalaman sekitar 0,5 m, dan disortir dengan hati-hati menggunakan saringan plastik berukuran mata jaring 1 mm yang digantung dalam ember berisi air laut 10 L. Pada hari berikutnya, sekitar 50.000 spat didistribusikan secara merata ke dalam lima tangki terpisah berukuran 10 L (~ 10.000 spat per tangki) dan disimpan pada suhu 17°C (ruangan dengan suhu terkontrol di GEOMAR) dengan 16 PSU air laut alami yang disaring melalui mata jaring 0,5 μm. Selama 8 hari berikutnya, praktik pemeliharaan melibatkan pertukaran air 50% setiap hari dan penambahan 100 mL Rhodomonas salina (~1–2 × 106 sel mL −1 ) sebagai makanan. Sisa 450.000 spat ditantang secara termal: pertama, didistribusikan secara merata di antara tangki 16 × 2 L (ca. 25.000 per tangki) yang diisi dengan 600 mL air laut alami 16 PSU, disaring dengan jaring 0,5 μm, dan diekspos selama 30 menit ke suhu tinggi yang tidak mematikan (26,5 °C), dengan aerasi konstan yang disuplai melalui ujung pipet plastik 5 mL. Tangki percobaan ditempatkan ke dalam empat penangas air baja yang mempertahankan suhu target dengan deviasi kurang dari 0,1 °C (model Haake SWB25, Thermo Scientific). Sebanyak 1 L air laut yang disaring (FSW; disaring melalui serangkaian filter 10 dan 1 μm mesh) pada suhu 30°C ditambahkan ke setiap tangki, penangas air diatur pada suhu 30°C dan suhu target tercapai dalam waktu ~ 15 menit. Spat kemudian disimpan pada suhu ini selama 47 jam.
Tiga hari pasca pengumpulan, semua spat yang diolah dengan panas (hidup atau mati) dicuci dalam FSW untuk menghilangkan jaringan remis yang mati dan kemudian dipindahkan dari tangki 16 × 2 L ke tangki 4 × 10 L dengan FSW (salinitas 16 PSU, suhu 17°C ± 0,2°C, aerasi konstan), yang ditempatkan berdekatan dengan tangki yang menampung hewan naif di ruangan yang suhunya terkontrol. Selama 2 hari pertama pasca pemindahan, penggantian air (~ 95% dari volume) dilakukan setiap hari untuk menghilangkan sisa-sisa organik hewan yang mati, diikuti dengan penambahan 200 mL R. salina (~ 1–2 × 106 sel mL −1 ) ke setiap tangki. Selama 9 hari berikutnya, praktik pemeliharaan beralih ke pertukaran air harian sebesar 50% dan penambahan 100 mL R. salina (~ 1–2 × 106 sel mL −1 ). Spat yang dipilih dengan panas kemudian diwarnai dengan calcein green dan hewan yang belum pernah dipijahkan diwarnai dengan calcein blue (50 mg L −1 pada hari ke-1, dan 25 mg L −1 pada hari ke-2–9), ditambahkan segera setelah air berubah untuk memberi label fluoresensi pada individu yang hidup. Kedua bahan kimia tersebut tidak beracun bagi kerang.
Pada 14 hari pasca pengumpulan, spat yang dipilih dengan panas dari empat tangki dicampur dan tiga sub-sampel kerang sebanyak 500 μL dipipet ke dalam tabung Eppendorf 0,5 mL, dengan tingkat kelangsungan hidup dinilai. Karena prosedur pelabelan cangkang kalsein, spat hidup tampak hijau terang di bawah mikroskop stereo yang dilengkapi dengan set filter GFP standar (Leica M165 FC), dan 600 spat hidup ditemukan dan disimpan untuk langkah-langkah berikutnya. Selain itu, sekitar 600 spat disortir secara acak dari ~ 50.000 spat naif yang disimpan di lima tangki pemeliharaan pada suhu konstan 17°C. Tidak ada kematian yang diamati pada kelompok naif. Setiap kelompok yang terdiri dari 600 spat naif atau yang dipilih kemudian didistribusikan secara merata di antara empat wadah 50 mL (150 individu di setiap wadah, empat wadah per baris spat), yang ditutup dengan kasa (300 μm). Dua wadah tersebut, satu berisi spat terpilih dan satu berisi spat tak terpilih, disimpan dalam satu akuarium 10 L (‘tangki pengondisian’, total empat tangki) selama 7 hari pada suhu 17°C dan 16 PSU, diberi makan setiap hari dengan konsentrasi awal sekitar 8000 sel R. salina mL −1 untuk memastikan kondisi yang sebanding antara spat dari dua kelompok perlakuan berbeda untuk percobaan berikut.
Pada bagian berikut dari naskah ini, spat yang bertahan dari paparan tekanan termal disebut sebagai ‘terpilih’ (S), sedangkan spat yang tidak terpapar tekanan termal disebut ‘tidak terpilih’ (NS). Spat ini terpapar tantangan termal di fasilitas Kiel Indoor Benthocosm (KIB, Pansch dan Hiebenthal 2019 ).
2.1.2 Paparan Stres Kedua
Setelah pengembangan awal kohort spat terpilih dan tidak terpilih, 960 individu (480 terpilih dan 480 tidak terpilih) dipilih secara acak untuk percobaan lanjutan, di mana kami menilai dampak stres termal pada kinerja juvenil. Dimulai pada tanggal 20 Agustus, spat diekspos pada lima perlakuan suhu yang berbeda (21°C, 22°C, 23°C, 24°C, 25°C, dan 26°C) selama periode 25 hari. Perlakuan 26°C mewakili suhu gelombang panas laut harian puncak akhir abad yang diprediksi di wilayah studi, sub-mematikan bagi remis untuk periode < 1 bulan. Perlakuan 21°C mewakili suhu musim panas maksimum saat ini, yang berada dalam kisaran optimal untuk pertumbuhan remis. Suhu antara dalam kisaran eksperimen dipilih untuk mengidentifikasi perubahan halus, tetapi signifikan, dalam kinerja termal atau titik henti termal dalam kinerja larva remis dalam studi ini. Percobaan dilakukan menggunakan KIB. Di fasilitas ini, tangki polietilena 12 × 600 L (yaitu, benthocosms) yang dilengkapi dengan komputer GHL Profilux dan sensor termal (GHL GmbH), yang mengendalikan pemanas dan pendingin (Aquamedic), memungkinkan pemeliharaan dan perekaman suhu air. Dua silinder eksperimen ditugaskan untuk masing-masing dari enam suhu eksperimen. Tangki KIB digunakan sebagai penangas air dan menampung 12 silinder inkubasi hewan (dua per penangas air). Hewan ditempatkan dalam kandang plastik (OD 30 mm, panjang sekitar 10 mm) yang disegel di bagian atas dan bawah dengan kasa nilon 300 μm (Gambar 1 ). Kandang eksperimen ditempatkan dalam tangki silinder plastik 14 L, yang memungkinkan aliran air dan penyerapan makanan terjadi. Dua puluh juvenil dipelihara di setiap kandang, dengan setiap perlakuan (S dan NS) memiliki dua kandang replikasi per tangki eksperimen. Dengan demikian, total 80 hewan S dan 80 NS diinkubasi pada setiap suhu. Kandang diikatkan ke pipet 15 mL yang digunakan untuk aerasi pada ketinggian kolom air yang sama, memberikan kondisi lingkungan yang sama antara replikasi setiap perlakuan (Gambar 1 ). Panjang cangkang awal diukur dalam sub-sampel yang terdiri dari 60 individu terpilih dan 60 individu tidak terpilih, yang dikeluarkan dari tangki pengondisian sebelum didistribusikan ke dalam kandang percobaan pada hari ke-0, 1 hari sebelum dimulainya percobaan paparan termal selama 25 hari. Dampak suhu tinggi pada spat remis dinilai dengan mengukur kinerja larva melalui pertumbuhan seperti panjang cangkang, berat jaringan kering, dan berat cangkang kering pada hari ke-25, di akhir tantangan, dari sedikitnya 10 hewan per kandang. Nilai data mentah di bagian hasil disajikan sebagai rata-rata ± simpangan baku. 2.1.3 Peternakan Sepanjang durasi percobaan ini, spat diberi makan dengan monokultur Rhodomonas salina , ditambahkan pada konsentrasi ~8000 sel mL −1 (Riisgård et al., 2011) ke silinder percobaan setelah setiap pertukaran air. Sebelum memulai percobaan utama, kami mengukur konsentrasi R. Salinas setiap hari selama 72 jam di semua tangki percobaan yang diatur pada suhu masing-masing, untuk menjamin bahwa penurunan konsentrasi sel disebabkan oleh konsumsi makanan daripada kematian alga (Tabel S1 ). Konsentrasi sel diukur setiap hari (ukuran partikel 5–8 um) menggunakan penghitung Coulter (Coulter Z2, Beckman Coulter GmbH), di semua tangki percobaan. Jika konsentrasi R. salina di salah satu tangki percobaan turun ke nilai ambang batas dalam kisaran 1000 sel mL −1 , pertukaran air penuh dilakukan di semua tangki untuk memastikan bahwa R. salina terus tersedia di kolom air pada konsentrasi optimal. Pergantian air dilakukan dengan FSW setiap 3 hari (hari 1-7), setiap dua hari sekali (hari 8-20), dan setiap hari setelah hari ke-21 (Tabel S2 ). 2.1.4 Penilaian Pertumbuhan, Massa Tubuh Kering dan Massa Cangkang Panjang cangkang diukur menggunakan jangka sorong. Berat jaringan kering dan berat cangkang diukur seperti yang dijelaskan sebelumnya. Singkatnya, sampel spat dimatikan dalam gelombang mikro (400 watt, 30 detik), dan jaringan lunaknya dikeluarkan dari cangkangnya di bawah mikroskop stereo. Cangkang dan jaringan lunak masing-masing ditempatkan secara terpisah ke dalam wadah aluminium foil yang telah ditimbang sebelumnya, dikeringkan pada suhu 80°C, minimal selama 72 jam, dan ditimbang ulang untuk menentukan berat jaringan kering dan berat cangkang (mg) dengan mengurangi nilai wadah aluminium foil kosong dari wadah aluminium foil yang berisi cangkang atau jaringan lunak. Nilai data mentah di bagian hasil disajikan sebagai rata-rata ± simpangan baku. 2.1.5 Penilaian Struktur dan Keterkaitan Genom Dua Kohort Komposisi genetik dari kelompok yang dipilih dan tidak dipilih dianalisis menggunakan 50 spat dari setiap perlakuan, yang dikumpulkan pada hari ke-0 percobaan kedua, yaitu, sehari sebelum dimulainya tantangan paparan selama 25 hari, dengan spat yang diambil sampelnya diawetkan dalam etanol 90%. Sampel dikirim ke Identigen Ltd. untuk ekstraksi dan pengurutan DNA, menggunakan array SNP kerang biru 60 K (Nascimento-Schulze et al. 2023 ). Untuk menganalisis data sekuensing yang dihasilkan, filter kendali mutu untuk tingkat pemanggilan penanda (CR) > 95% dan CR sampel > 90% diterapkan menggunakan PLINK v1.9. Kami menerapkan filter frekuensi alel minor (MAF) < 0,01. Analisis komponen utama (PCA) diterapkan pada genotipe yang dihasilkan dengan array, memangkas lokus yang diduga terkait menggunakan ukuran jendela 50 Kb, ukuran langkah 10 Kb dan ambang r 2 sebesar 0,1 dengan PLINK v1.7 (Purcell et al. 2007 ). Kami mengeksplorasi introgresi dalam spat dengan menerapkan analisis campuran (campuran v1.3; Alexander dan Lange 2011 ). Dalam analisis ini, jumlah klaster yang paling sesuai yang mewakili leluhur suatu populasi diperkirakan di mana k memiliki nilai kesalahan validasi silang terendah. Untuk penelitian ini, kami menguji nilai k antara 2 dan 10. Untuk mengalokasikan spesies ke klaster leluhur yang dihasilkan dalam analisis campuran pertama ini, kami menjalankan analisis campuran tambahan yang mencakup 5 populasi tambahan, Kiel (Jerman Baltik, GK), Ahrenshoop (Jerman Baltik, GA), Finlandia (Finlandia Baltik, FIN), Bude (pantai Devon Utara Inggris) dan Budleigh (pantai Devon Selatan Inggris). Populasi-populasi ini telah digenotipe dengan susunan multispesies kerang biru 60 K dalam penelitian sebelumnya (Nascimento-Schulze et al. 2023 ; Nascimento-Schulze et al. data tidak dipublikasikan), dan leluhur mereka diketahui sebagai M. edulis/M. hibrida trossulus untuk populasi GK dan GA dan FIN, dengan peningkatan frekuensi genotipe M. trossulus di FIN, nenek moyang M. galloprovincialis mendominasi Bude dan M. edulis yang dominan di Budleigh. Hasil campuran dari kumpulan data ini disajikan dalam Tabel S3 dan S4 , serta divisualisasikan dalam Gambar S1 dan S2 .
2.1.6 Analisis Statistik
2.1.6.1 Kinerja Spat
Data panjang cangkang yang dikumpulkan dari individu pada hari ke-0, sebelum dimulainya tantangan termal selama 25 hari, pertama-tama diuji untuk kenormalan dan homogenitas varians menggunakan uji Shapiro–Wilk dan Levene masing-masing sebelum analisis ANOVA satu arah diterapkan untuk menguji efek pemilihan suhu.
Kami menyelidiki efek suhu pada berat jaringan kering, berat cangkang dan panjang cangkang individu terpilih dan tidak terpilih setelah tantangan termal 25 hari. Efek ini dimodelkan sebagai model campuran aditif umum (GAMM) menggunakan fungsi gam dari paket mgcv . Dalam GAMM, suhu didefinisikan sebagai prediktor efek halus (dengan efek linear dan/atau nonlinier) dan seleksi sebagai faktor terurut. Untuk menguji efek suhu di dalam dan di antara kelompok terpilih dan tidak terpilih, suhu dimodelkan sebagai penghalus dalam setiap level kelompok seleksi (terpilih vs. tidak terpilih). Lebih khusus lagi, kami menggunakan statistik-t dan statistik-f (uji Wald), intersep (atau mean) dan kemiringan atau nonlinier (derajat kebebasan efektif, edf ) dari penghalus level referensi ( tidak terpilih ), dengan masing-masing dibandingkan dengan nol, berturut-turut; dan estimasi penghalus level perlakuan ( terpilih ) kemudian dibandingkan dengan level referensi. Selain itu, efek acak dari penangas air dan sangkar , sebagai kemungkinan penyebab ketergantungan residual, disertakan, dan untuk estimasi komponen varians yang tidak bias, GAMM dipasang menggunakan kemungkinan maksimum yang dibatasi.
Model-model tersebut pertama kali dipasang dengan fungsi tautan identitas dengan asumsi residual memiliki distribusi gaussian di sekitar mean yang diprediksi. Asumsi mengenai distribusi residual diperiksa melalui paket DHARMa . Karena ditentukan bahwa asumsi residual dilanggar, kami memasang model dengan asumsi distribusi t berskala ( scat ) dari residual dan menemukan asumsi ini valid. Untuk panjang shell, fungsi tautan log 10 juga diperlukan untuk memvalidasi asumsi residual. Dalam hal Kriteria Informasi Akaike (AIC), model scat juga lebih unggul daripada model gaussian. Kami menggunakan fungsi prediksi dari paket car untuk memprediksi mean dan interval kepercayaan respons terhadap prediktor. Nilai p < 0,05 diasumsikan sebagai tingkat signifikansi untuk semua analisis. Semua analisis dilakukan dalam R v4.3.1. Nilai data mentah di bagian hasil disajikan sebagai mean ± simpangan baku. 2.1.6.2 Komposisi Genetik Populasi Kami membandingkan komposisi genetik dari dua kelompok kerang (terpilih dan tidak terpilih) untuk menyelidiki perbedaan potensial dalam proporsi leluhur yang dihasilkan dari peristiwa seleksi. Untuk ini, kami menetapkan model regresi Bayesian dengan mengasumsikan distribusi Logistik-Normal untuk proporsi leluhur, menggunakan paket brms (Bürkner 2017 ) dalam R v4.3.1. Model Logistik-Normal Bayesian dipilih karena memungkinkan perbandingan proporsi rata-rata dan variabilitas dalam data komposisi, sambil memperhitungkan korelasi antara komponen, menjadikannya pilihan yang kuat untuk analisis kami. Model tersebut dipasang menggunakan No-U-Turn Sampler (NUTS) dalam kerangka Hamiltonian Monte Carlo. Deskripsi menyeluruh dari model yang diuji dan diterapkan tersedia dalam Informasi Tambahan, Bagian II ; Gambar S4–S6 ).
3 Hasil
3.1 Dampak Peningkatan Suhu terhadap Kelangsungan Hidup Kelompok dan Susunan Genetik
Pada 10 hari pasca tantangan termal, tingkat kelangsungan hidup kelompok kerang yang dipilih adalah sekitar 8%. Tidak ada kematian yang diamati pada kerang yang tidak dipilih selama periode ini. Dari 15.049 lokus yang digenotipe dengan CR > 95%, total 11.288 lokus dipertahankan setelah menerapkan MAF > 0,01. Setelah pemangkasan varian, total 10.085 lokus yang diduga tidak terkait digunakan untuk analisis PCA dan Admixture.
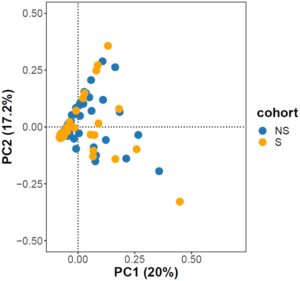
Analisis PCA tidak mengungkap adanya stratifikasi yang jelas antara kelompok yang dipilih dan yang tidak dipilih (Gambar 2 ). Dalam analisis ini, PC1 menjelaskan 20% varians, sedangkan PC2 dan PC3 menjelaskan masing-masing 17,2% dan 7,64%.
Jumlah klaster leluhur yang paling sesuai dalam analisis campuran adalah tiga (Tabel S4 ). Hasil dari koefisien campuran (Q) yang menyimpulkan tiga klaster disajikan dalam Gambar 3. Berdasarkan analisis, termasuk populasi kontrol (Gambar S1, S2 ), kami dapat menyimpulkan spesies untuk masing-masing dari tiga klaster yang ada dalam kohort S dan NS, dengan klaster biru tua mewakili leluhur M. edulis , M. trossulus biru muda , dan M. galloprovincialis kuning . Dalam analisis ini, kami mengamati bahwa sebagian besar genotipe dialokasikan ke M. edulis , diikuti oleh M. trossulus dan terakhir M. galloprovincialis .
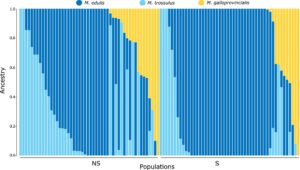
Komposisi genetik populasi terpilih vs. populasi tak terpilih menunjukkan tren berbeda untuk M. edulis dan M. galloprovincialis . Proporsi mentah dari analisis campuran menunjukkan bahwa populasi tak terpilih terdiri dari M. edulis (0,58), M. trossulus (0,29), dan M. galloprovincialis (0,13), sedangkan populasi terpilih memiliki proporsi M. edulis (0,74), M. trossulus (0,15), dan M. galloprovincialis (0,11). Pemodelan efek campuran Bayesian pada skala logit menunjukkan bukti kuat untuk peningkatan proporsi M. edulis pada populasi terpilih dibandingkan dengan populasi tak terpilih, dengan estimasi intersep tak terpilih sebesar 2,40 (IK 95%: 0,38 hingga 4,38), dan efek populasi terpilih 3,75 (IK 95%: 0,74 hingga 6,42) ( Informasi Pendukung, Bagian II ). Peningkatan genotipe M. edulis mengorbankan M. trossulus , karena meskipun model menunjukkan tren positif terhadap peningkatan proporsi M. galloprovincialis pada populasi terpilih, interval kredibel mencakup nilai negatif dan karenanya tidak signifikan (1,93,IK 95%: −0,74 hingga 4,62) ( Informasi Pendukung, Bagian II ), sehingga peningkatan signifikan M. edulis disebabkan oleh penurunan signifikan timbal balik M. trossulus . Parameter khusus famili menunjukkan variabilitas signifikan dalam proporsi M. edulis ( σ q2 = 7,24, 95% CI: 6,32–8,33) dan M. galloprovincialis ( σ q3 = 6,73, 95% CI: 5,87–7,74) di antara individu, yang menunjukkan heterogenitas yang cukup besar dalam populasi. Selain itu, model menemukan korelasi positif sedang antara leluhur M. edulis dan M. galloprovincialis (korelasi = 0,48, 95% CI: 0,33–0,62), yang menunjukkan bahwa peningkatan proporsi leluhur M. edulis dalam populasi cenderung disertai dengan peningkatan leluhur M. galloprovincialis .
3.2 Dampak Peningkatan Suhu terhadap Panjang Cangkang, Berat Jaringan Kering, dan Massa Cangkang
Tidak ada perbedaan signifikan dalam panjang cangkang yang diamati antara kelompok yang dipilih (2,02 ± 0,33 mm) dan yang tidak dipilih (1,99 ± 0,32 mm) pada hari ke-0, sebelum dimulainya tantangan termal 25 hari (Gambar S3 , Tabel S5 , p > 0,05).
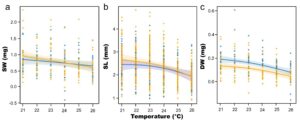
Pemaparan spat terhadap suhu eksperimen yang berbeda selama periode 25 hari tidak menghasilkan perbedaan yang signifikan pada berat cangkang masing-masing individu (Gambar 4a , Tabel 1 ), dengan berat cangkang spat yang dipilih sebesar 0,79 ± 0,5 mg, sedangkan spat yang tidak dipilih sebesar 0,77 ± 0,36 mg.
| SW 25 hari | Memperkirakan | Kesalahan Standar | T | P |
|---|---|---|---|---|
| A. Koefisien parametrik | ||||
| Intercept (tidak dipilih) | 0.7492 | 0,0392 | 19.0895 | < 0,0001 |
| Intercept (dipilih) | -0,0315 | 0,0510 | -0,6189 | 0.5360 |
| SW 25 hari | pendidikan | Ref.df | F | P |
|---|---|---|---|---|
| B. Istilah halus | ||||
| s(suhu, tidak dipilih) | 1.4395 | 1.6167 | 2.0979 | 0.1726 |
| s(suhu, dipilih) | 1.0001 | 1.0002 | 0.6174 | 0.4320 |
| kandang | 6.0661 | 22.0000 | 8.6526 | 0,0832 tahun |
| SL 25 hari | Memperkirakan | Kesalahan Standar | T | P |
|---|---|---|---|---|
| A. Koefisien parametrik | ||||
| Intercept (tidak dipilih) | 0,3777 tahun | 0,0081 | 46.3824 | < 0,0001 |
| Intercept (dipilih) | 0,0126 | 0,0068 tahun | 1.8364 | 0,0663 tahun |
| SL 25 hari | pendidikan | Ref.df | F | P |
|---|---|---|---|---|
| B. Istilah halus | ||||
| s(suhu, tidak dipilih) | 1.7442 | 1.7886 | 13.3916 | 0,0006 |
| s(suhu, dipilih) | 1.0005 | 1.0010 | 2.3425 | 0.1260 |
| kandang | 16.8297 | 23.0000 | 80.1398 | < 0,0001 |
| DW 25 hari | Memperkirakan | Kesalahan Standar | T | P |
|---|---|---|---|---|
| A. Koefisien parametrik | ||||
| Intercept (tidak dipilih) | 0.1418 | 0,0087 tahun | 16.3570 | < 0,0001 |
| Intercept (dipilih) | -0,0459 | 0,0115 | -3.9794 | 0,0001 |
| DW 25 hari | pendidikan | Ref.df | F | P |
|---|---|---|---|---|
| B. Istilah halus | ||||
| s(suhu, tidak dipilih) | 1.6596 | 1.8153 | 9.6515 | 0,0001 |
| s(suhu, dipilih) | 1.3257 | 1.5142 | 0.2626 | 0,5926 tahun |
| kandang | 4.6894 | 22.0000 | 0.2925 | 0.1261 |
Tidak ada dampak signifikan dari suhu atau seleksi yang diamati pada panjang cangkang antara kedua kelompok (Gambar 4b , Tabel 1 , p > 0,05). Panjang cangkang rata-rata kelompok yang dipilih dan tidak dipilih pada hari ke-25 masing-masing adalah 2,33 ± 0,52 mm dan 2,47 ± 0,67 mm, yang menunjukkan efek seleksi yang tidak signifikan ( p = 0,0663) pada pertumbuhan selanjutnya. Efek halus suhu secara keseluruhan signifikan, seperti yang ditunjukkan oleh kelompok yang tidak dipilih ( p = 0,0006), tetapi tidak berbeda secara signifikan antara kedua kelompok ( p = 0,1260), dengan juvenil yang tidak dipilih pada suhu 26°C lebih kecil (2,04 mm ± 0,45) daripada juvenil pada suhu 21°C (2,37 ± 0,49 mm).
Untuk analisis berat jaringan kering, intersep untuk kelompok terpilih, yang menunjukkan perbedaannya dari garis dasar yang tidak terpilih, secara signifikan lebih rendah ( p = 0,0001; Gambar 4c , Tabel 1 ). Hasil ini menunjukkan pengaruh seleksi awal terhadap massa jaringan kering di semua perlakuan suhu. Pengaruh suhu yang halus secara keseluruhan signifikan, seperti yang ditunjukkan oleh kelompok yang tidak terpilih ( p = 0,0001), tetapi tidak berbeda secara signifikan antara kelompok terpilih dan tidak terpilih ( p = 0,5926). Berat jaringan kering yang diamati dari spat yang tidak terpilih menurun dari 0,16 ± 0,06 mg pada 21°C menjadi 0,06 ± 0,10 mg pada 26°C, dan berat jaringan kering dari spat terpilih menurun dari 0,14 ± 0,11 mg pada 21°C menjadi 0,05 ± 0,15 mg pada 26°C.
Hubungan ini dimodelkan sebagai model campuran aditif dengan asumsi distribusi t residual berskala. Intersep (atau rata-rata) dan kemiringan atau nonlinieritas (derajat kebebasan efektif, edf) dari penghalus level referensi (naif) masing-masing dibandingkan dengan nol; dan, estimasi penghalus level perawatan (terpilih) kemudian dibandingkan dengan level referensi. Selain itu, efek intersep acak dari kurungan diuji. Dampak signifikan dipertimbangkan jika nilai p ≤ 0,05.
4 Diskusi
Dalam penelitian ini, kami menilai dampak suhu musim panas yang sangat tinggi terhadap variasi genetik dan kinerja Mytilus spp. juvenil Baltik. Kami menemukan bahwa seleksi terhadap suhu yang sangat tinggi mengakibatkan pengayaan genotipe yang didominasi M. edulis dan secara bersamaan menyebabkan berkurangnya pertumbuhan jaringan, dalam uji pertumbuhan lanjutan pada semua suhu yang diuji. Oleh karena itu, kami memberikan bukti pertama bahwa suhu dapat memainkan peran penting dalam komposisi genomik dan fenotipik kerang Mytilus di Laut Baltik. Tren ini terbukti setelah hanya satu generasi dan peristiwa seleksi, dan mungkin merupakan indikasi proses yang terjadi di alam liar selama musim panas yang hangat. Selain itu, respons cepat terhadap seleksi ini menunjukkan bahwa Mytilus spp mungkin cocok untuk seleksi buatan guna mengubah komposisi genomik dan sifat fisiologis populasi, sehingga meningkatkan ketahanan mereka terhadap perubahan global yang semakin ekstrem. Lebih banyak pekerjaan diperlukan untuk memahami bagaimana genotipe merespons seleksi di alam liar, dan untuk menunjukkan apakah pengembangan populasi yang tahan panas melalui seleksi di laboratorium selama beberapa generasi dapat dilakukan. Namun, hasil kami merupakan langkah awal yang penting untuk menunjukkan bahwa pekerjaan tambahan ini diperlukan dan menjanjikan.
4.1 Dampak Peningkatan Suhu terhadap Variasi Genetik Kerang Spat
Kami menyelidiki apakah paparan terhadap stres termal dapat memodifikasi struktur genetik populasi kerang biru muda dengan membandingkan komposisi genetik dua kelompok spat, sebelum dan sesudah seleksi, menggunakan multi-spesies 60 K SNP-array (Nascimento-Schulze et al. 2023 ). Latar belakang genetik populasi secara umum sesuai dengan data yang diterbitkan sebelumnya tentang kerang yang berasal dari Laut Baltik, di mana tiga spesies berkontribusi pada populasi: M. edulis , M. trossulus dan M. galloprovincialis . Meskipun demikian, kontribusi nenek moyang M. galloprovincialis dalam spat yang dianalisis dalam penelitian ini jauh lebih tinggi daripada yang telah dijelaskan sebelumnya di wilayah ini. Sementara Vendrami et al. ( 2020 ) membuat genotipe kerang dewasa, juvenil (berusia sekitar 2 bulan) dibuat genotipe dalam penelitian kami. Laut Baltik merupakan lingkungan yang unik karena gradien salinitasnya dan terbatasnya pencampuran air dengan Atlantik Utara melalui Selat Denmark. Individu yang memiliki alel M. galloprovincialis dapat secara sporadis diangkut ke wilayah ini oleh arus air, dibawa oleh air pemberat, atau diikatkan pada kapal rekreasi lainnya. Ada kemungkinan bahwa kelangsungan hidup genotipe yang berbeda melalui perkembangan dapat menjelaskan perbedaan leluhur. Namun, diperlukan lebih banyak penelitian untuk memverifikasi hipotesis ini.
Kami menemukan bukti bahwa seleksi suhu menggeser komposisi genetik kerang biru, dengan frekuensi genotipe M. edulis yang lebih tinggi dalam kelompok yang dipilih dibandingkan dengan populasi yang tidak dipilih. Mortalitas yang bergantung pada genotipe sebelumnya telah diamati pada kerang (Han et al. 2020 ; Koehn et al. 1980 ), mendukung respons cepat terhadap seleksi dalam penelitian kami. Misalnya, seleksi pasca-penyelesaian yang bekerja pada alel Lap telah diamati pada M. edulis yang menghuni gradien salinitas, dengan keberadaan alel menurun melalui perkembangan dalam salinitas rendah (Koehn et al. 1980 ). SST yang ditemukan di seluruh distribusi alami M. edulis dan M. galloprovincialis lebih tinggi daripada yang ditemukan dalam kisaran M. trossulus (Tabel S6 ; Popovic dan Riginos 2020 ). Pengamatan ini mendukung penurunan proporsi alel M. trossulus dalam populasi terpilih dalam penelitian kami yang disebabkan oleh seleksi terhadap genotipe yang toleran terhadap suhu. Adaptasi lokal terhadap salinitas rendah mungkin lebih mendukung peningkatan genotipe M. edulis daripada M. galloprovincialis dalam populasi terpilih. Meskipun demikian, meskipun menunjukkan pergeseran signifikan dalam genotipe populasi antara populasi yang tidak terpilih dan terpilih, ketidakpastian dari pemodelan Bayesian menunjukkan sedikit kehati-hatian harus diambil ketika menafsirkan hasil ini, karena heterogenitas genomik dalam leluhur pada tingkat individu, dan dengan demikian ketidakpastian dari keseluruhan besarnya pergeseran ini. Selain itu, penelitian lebih lanjut diperlukan untuk memahami bagaimana perubahan dalam komposisi genomik populasi spat terwujud dalam populasi dewasa.
Pada tingkat genomik, adaptasi cepat setelah peristiwa seleksi satu generasi telah ditunjukkan sebelumnya pada beberapa spesies invertebrata, termasuk salinitas pada larva spesies tiram yang berkerabat dekat (Griffiths et al. 2021 ), serta pengasaman laut pada remis (Bitter et al. 2019 ) dan larva bulu babi (Brennan et al. 2019 ). Meskipun demikian, ini adalah studi pertama yang menyelidiki adaptasi cepat genotipe populasi setelah peristiwa seleksi satu generasi dalam sistem studi yang terdiri dari individu-individu dengan keturunan campuran. Dengan demikian, hasil kami memberikan kemajuan penting dalam pemahaman kami tentang interaksi spesies dan introgresi di zona hibrida dalam kondisi iklim yang berubah. Namun, untuk sepenuhnya memahami dampak seleksi ini pada komposisi populasi, penting bagi penelitian di masa depan untuk menentukan apakah efek ini bertahan sepanjang waktu dan generasi, atau apakah seleksi yang seimbang secara temporal dapat mengurangi dampak peristiwa ini pada tingkat populasi (misalnya, Durland et al. 2021 ). Selain itu, penting untuk menilai dampak stres lingkungan di semua tahap kehidupan: sementara penelitian kami difokuskan pada juvenil, tekanan seleksi dapat sangat bervariasi di antara tahap kehidupan. Menyelidiki perbedaan ini akan memberikan pemahaman yang lebih komprehensif tentang bagaimana seleksi cepat membentuk lintasan evolusi organisme ini.
4.2 Dampak Peningkatan Suhu terhadap Berat Jaringan Kering, Panjang Cangkang, dan Massa Cangkang
Studi kami menemukan bahwa setelah 25 hari terpapar suhu tinggi, cangkang yang dipilih memiliki berat jaringan kering yang lebih rendah dibandingkan dengan cangkang yang tidak dipilih. Selain itu, panjang cangkang cangkang pada cangkang yang tidak dipilih menurun seiring dengan peningkatan suhu.
Dampak suhu yang meluas, dari tingkat molekuler hingga seluruh organisme, memengaruhi kinerja dan menentukan jendela termal suatu populasi. Studi pada Mytilus galloprovincialis menunjukkan bahwa pemanasan antara 24°C–26°C, menggeser piruvat kinase ke bentuk enzim yang kurang aktif, yang menyebabkan hipometabolisme dan selanjutnya ke penekanan metabolik (Anestis et al. 2007 ). Penekanan metabolik ini memungkinkan individu untuk menurunkan kebutuhan energi mereka dan mengalokasikan energi surplus untuk pemeliharaan fungsi dasar di lingkungan yang hangat (Schulte et al. 2011 ), dan telah diamati pada remis Baltik pada suhu di luar ambang batas populasi (Vajedsamiei, Melzner, et al. 2021 ; Vajedsamiei, Wahl, et al. 2021 ). Oleh karena itu, penekanan metabolik berpotensi memungkinkan energi dialokasikan dari pertumbuhan ke proses fisiologis yang diperlukan untuk melawan suhu ekstrem. Pertukaran energi ini mungkin telah berkontribusi terhadap kelangsungan hidup kelompok terpilih selama peristiwa seleksi termal, tetapi kemungkinan besar dengan mengorbankan berkurangnya pertumbuhan jaringan pada individu terpilih.
Suhu eksperimen yang lebih tinggi, 23°C–26°C, merangkum kondisi yang akan dihadapi populasi kerang selama periode musim panas terhangat yang paling ekstrem, dari saat ini hingga akhir abad ini (Meier et al. 2022 ; Pansch et al. 2018 ; Vajedsamiei, Melzner, et al. 2021 ; Vajedsamiei, Wahl, et al. 2021 ). Kerang muda mau tidak mau akan mengalami peningkatan SST dan peristiwa cuaca ekstrem selama perkembangannya di Laut Baltik. Hasil kami menunjukkan bahwa suhu ini dapat melebihi optima termal untuk kerang muda ini. Meskipun demikian, untuk memastikan apakah seleksi telah mengutamakan individu dengan laju metabolisme yang lebih rendah, konsumsi oksigen, laju makan dan ekskresi harus dinilai dalam studi lanjutan. Lebih jauh lagi, untuk menilai apakah berat jaringan kering yang berkurang menunjukkan gangguan fungsi organisme, persistensi fenomena ini hingga organisme dewasa juga harus diselidiki.
5 Kesimpulan dan Perspektif Masa Depan
Perubahan iklim diproyeksikan akan mengubah parameter fisikokimia kelautan secara global, yang telah relatif stabil selama ribuan tahun, dengan implikasi bagi ekosistem laut. Kerang biru merupakan kelompok spesies yang memiliki nilai ekonomi dan ekologi yang tinggi; dengan demikian, menjamin bahwa stok alami dan komersial dapat berkembang dalam kondisi lingkungan yang baru merupakan hal yang sangat penting.
Ini adalah studi pertama yang mengkarakterisasi secara mendalam komposisi genetik spat dari pantai Selatan Laut Baltik, menggunakan pendekatan genomik. Dengan menggunakan pendekatan seleksi satu generasi terhadap suhu tinggi, kami mengamati peningkatan yang jelas dalam dominasi leluhur M. edulis dalam populasi yang dipilih. Studi kami menunjukkan bahwa seleksi memengaruhi kinerja spat, dengan individu yang dipilih mampu mempertahankan pertumbuhan cangkang pada tingkat yang sebanding dengan individu NS, tetapi dengan mengorbankan pemeliharaan pertumbuhan jaringan somatik. Lebih jauh, ketahanan yang lebih tinggi dan respons cepat terhadap seleksi genotipe M. edulis terhadap suhu tinggi menunjukkan bahwa alel dari spesies ini dapat memberikan ketahanan terhadap peningkatan suhu yang disebabkan oleh perubahan global, bahkan dalam populasi yang sangat bercampur. Perbedaan dalam kinerja dan pergeseran cepat dalam komposisi genetik antara kelompok yang dipilih dan tidak dipilih dalam studi kami menunjukkan peran penting suhu dalam kompleks spesies ini yang membenarkan penyelidikan lebih lanjut. Informasi tersebut akan memajukan pemahaman dan kemampuan kita untuk mengukur respons kerang biru terhadap stres termal, serta kemampuan mereka untuk menyesuaikan diri dan/atau beradaptasi dengan stresor tersebut, di lautan masa depan. Menggabungkan penggunaan perangkat genom kerang biru dengan pengetahuan tentang mekanisme fisiologis khusus spesies pada akhirnya dapat berkontribusi pada pengembangan kelompok yang tahan terhadap suhu dan merupakan jalan utama untuk penelitian lebih lanjut.


