
ABSTRAK
Ekologi dan genetika lanskap menyediakan kerangka kerja analitis penting untuk menyelidiki dampak fitur lingkungan pada proses ekologi. Namun, beberapa studi empiris telah menguji secara bersamaan bagaimana karakteristik lanskap memengaruhi pola spasial aliran gen dan kelimpahan serangga hama di lingkungan yang heterogen. Untuk mengatasi hal ini, kami melakukan studi gabungan ekologi lanskap/genetik lanskap terhadap lalat buah tephritid, Bactrocera tryoni , di wilayah Wide Bay-Burnett di Queensland Tenggara, Australia. Wilayah ini berisi area hutan hujan, hutan Eucalyptus , padang rumput yang dibuka, area pemukiman, dan dua area produksi hortikultura intensif; satu menerapkan praktik manajemen di seluruh area. Kami mengumpulkan sampel B. tryoni dari 26 lokasi pada tahun 2021 selama bulan April, Agustus, Oktober, Desember dan tahun berikutnya selama Februari dan April. Kami menggunakan genotipe SNP DArTseq kepadatan tinggi pada sampel yang dikumpulkan selama periode pengambilan sampel April, Agustus, dan Desember 2021. Kami kemudian memodelkan karakteristik lanskap kontemporer dan faktor manajemen yang memengaruhi aliran gen dan kelimpahan spesies hama ini. Analisis SNP di seluruh genom memperkirakan ukuran populasi efektif tak terbatas di semua lokasi dan mendeteksi struktur genetik terbatas di seluruh lanskap. Namun, kelimpahan lalat bervariasi secara signifikan di antara habitat, dengan padang rumput yang dibuka berhubungan negatif dengan kelimpahan populasi dan bertindak sebagai penghalang aliran gen. Selain itu, jalan raya komposit dengan padang rumput yang dibuka menunjukkan efek penghalang yang sangat kuat. Kelimpahan tertinggi berada di daerah pemukiman dan hutan hujan, terendah di hutan Eucalyptus , dan berkurang di wilayah hortikultura dengan manajemen di seluruh wilayah yang diterapkan. Kami membahas manfaat pengumpulan data genetik dan ekologi secara simultan untuk menginformasikan dan mengevaluasi program manajemen di seluruh wilayah untuk hama serangga dan menyoroti pertimbangan dalam analisis spasial data SNP ketika ukuran populasi efektif sangat besar.
1 Pendahuluan
Bahasa Indonesia: Banyak hama pertanian terpenting di dunia bersifat mobil dan polifag, yang berarti bahwa mereka dapat berkembang biak di sumber nontanaman sebelum bermigrasi dan berkembang biak di tanaman—sehingga pengelolaan di lahan saja tidak cukup (Walter 2003 ). Untuk mengatasi masalah invasi dari area nontanaman ini, pengelolaan di seluruh area (AWM) dapat dimanfaatkan (Vreysen et al. 2007 ). Pendekatan ini menargetkan penekanan populasi hama di seluruh lanskap, daripada mencoba mengelola masalah di tingkat pertanian individu (Hendrichs et al. 2007 ). Informasi tentang habitat yang disukai hama di dalam lanskap, dan bagaimana dan kapan hama bergerak (atau tidak) melintasi lanskap dan di antara petak habitat dianggap sebagai pengetahuan inti untuk penerapan pendekatan ini (Byrne 2008 ).
Karena bersifat polifag dan bergerak, lalat buah tephritid (Diptera: Tephritidae) sering menjadi target program AWM (Hendrichs et al. 2002 ; Vargas et al. 2008 ; Aluja et al. 2012 ; Rashid et al. 2021 ). Pengetahuan tentang lokasi tephritid berada di lanskap dapat diperoleh dari penangkapan lalat dewasa dan pengumpulan buah untuk larva (Vargas et al. 1990 ; Kounatidis et al. 2008 ; Ortega et al. 2021 ) atau dari pelepasan dan penangkapan kembali individu yang ditandai (MacFarlane et al. 1987 ; Iwaizumi dan Shiga 1989 ; Peck dan McQuate 2004 ; Froerer et al. 2010 ). Studi semacam itu cenderung menunjukkan bahwa populasi terbentuk di tempat inang perkembangbiakan tersedia (Vargas et al. 1983 ; Paredes et al. 2022 ) dan bahwa sementara pergerakan individu dewasa biasanya terbatas pada beberapa ratus meter, penyebaran jarak jauh lebih dari 10 km bukanlah hal yang tidak umum (MacFarlane et al. 1987 ; Froerer et al. 2010 ). Namun, studi semacam itu biasanya berfokus pada titik akhir penyebaran, dan pentingnya atribut lanskap seperti lahan terbuka, jalur air, hutan tertutup, dll. dalam membentuk pola penyebaran tephritid sebagian besar masih disimpulkan daripada diuji secara langsung.
Lalat buah Queensland, Bactrocera tryoni (Froggatt), adalah serangga hama hortikultura terpenting di Australia (Dominiak 2019 ). Penelitian tentang penekanan populasi atau pemberantasan B. tryoni di tingkat lanskap dimulai 60 tahun yang lalu (Monro 1960 ; Bateman et al. 1966 ) dan terus berlanjut hingga saat ini (Jessup et al. 2007 ; Lloyd et al. 2010 ; Tam et al. 2020 ; Dominiak et al. 2022 ). Penelitian lain telah menggunakan penggalian pendapat ahli untuk menentukan peringkat kesesuaian habitat (van Klinken et al. 2019 ) atau menggunakan pemodelan lanskap untuk membuat prediksi teoritis tentang pergerakan B. tryoni yang acak atau sempurna di lanskap virtual (Schwarzmueller et al. 2019 ). Namun, meskipun ada upaya penelitian yang ekstensif ini, studi empiris tentang ekologi lanskap B. tryoni masih kurang (Clarke et al. 2011 ). Lebih jauh lagi, tidak ada studi yang ada yang memperhitungkan keterkaitan genetik, yang mencerminkan kepadatan dan penyebaran individu di seluruh lanskap (Wright 1943 ). Memanfaatkan informasi genetik dan teori genetika populasi dapat mengatasi keterbatasan pemantauan hama tradisional (Beaurepaire et al. 2024 ) dengan secara langsung melacak pola penyebaran di antara populasi yang diduga dan menilai pengaruh variabel lanskap pada pola penyebaran yang diamati tanpa informasi apriori (Baguette et al. 2013 ).
Untuk mengatasi kesenjangan pengetahuan ini, kami melakukan studi gabungan ekologi bentang alam/genetika bentang alam terhadap B. tryoni di wilayah Wide Bay-Burnett di Queensland Tenggara, bagian dari wilayah endemik spesies tersebut. Wilayah tersebut, yang meliputi area seluas sekitar 19.800 km 2 , berisi area hutan hujan, yang diasumsikan sebagai habitat endemik lalat buah Bactrocera (Drew 1989 ), serta hutan eukaliptus, lahan penggembalaan pertanian, dan dua area produksi hortikultura intensif (distrik Bundaberg dan Gayndah-Mundubbera). Gayndah-Mundubbera telah menjalankan pengendalian B. tryoni di seluruh area sejak awal tahun 2000-an (Lloyd et al. 2007 , 2010 ), sementara petani di Bundaberg mengelola lalat buah di lahan pertanian secara ad-hoc (Missenden 2014 ; Senior 2016 ). Bahasa Indonesia: Dalam makalah ini, kami melaporkan temuan dari studi selama setahun terhadap B. tryoni dewasa yang terperangkap setiap 2 bulan dari susunan lokasi yang terstruktur secara spasial. Kami memeriksa kelimpahan B. tryoni sehubungan dengan jenis penggunaan lahan, atau jarak dari AWM. Selain itu, kami memeriksa pengaruh jenis penggunaan lahan pada pola spasial dan temporal skala halus dari struktur genetik di B. tryoni menggunakan data polimorfisme nukleotida tunggal (SNP) di seluruh genom pada tiga waktu pengambilan sampel (April, Agustus, Desember) yang sesuai dengan akhir, awal, dan tengah siklus perkembangbiakan tahunan spesies tersebut. Populasi Bactrocera tryoni menurun drastis antara April dan Agustus selama kemungkinan periode penuaan tetapi kemudian meningkat dengan cepat lagi hingga Desember sebelum perlahan menurun hingga Mei (Clarke et al. 2022 ). Pemilihan sampel April, Agustus, dan Desember untuk analisis genetik dilakukan berdasarkan asumsi apriori bahwa 3 bulan ini akan dikaitkan dengan variasi genetik maksimum di dalam dan di antara populasi yang berbeda dalam lanskap. Metode keterkaitan lanskap dan pemodelan isolasi melalui resistensi (IBR) digunakan untuk memahami apakah jenis penggunaan lahan bertindak sebagai penghalang atau fasilitator penyebaran. Estimasi ukuran populasi efektif juga dihitung untuk setiap lokasi. Sepengetahuan kami, ini adalah studi genetik dan ekologi lanskap gabungan pertama dari spesies tephritid.
2 Metode
2.1 Pengambilan sampel
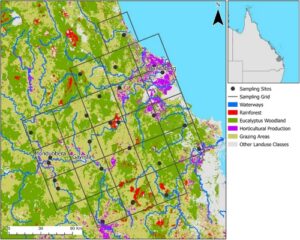
Sampel Bactrocera tryoni dikumpulkan dari 26 lokasi di wilayah Wide-Bay/Burnett di Queensland, Australia (Gambar 1 ) pada bulan April 2021, Agustus 2021, Oktober 2021, Desember 2021, Februari 2022, dan April 2022. Area ini dipilih karena memiliki berbagai jenis penggunaan lahan, yang memungkinkan pengambilan sampel berulang. Selain itu, area ini mencakup dua area produksi hortikultura yang berbeda: Mundubbera/Gayndah dan Bundaberg. Area produksi Mundubbera/Gayndah memiliki program pengelolaan lalat buah permanen di seluruh area, sementara pengelolaan di area produksi Bundaberg dilakukan berdasarkan pertanian per pertanian. Agar lokasi pengambilan sampel tersebar di seluruh area studi, kami menggunakan struktur kisi untuk pengambilan sampel, dengan setiap kotak kisi berukuran 30 km × 30 km. Desain pengambilan sampel kami terdiri dari 22 kotak kisi, dengan satu lokasi pengambilan sampel dipilih dalam setiap kotak, dengan lokasi pengambilan sampel tambahan di empat dari 22 kotak kisi untuk mengambil sampel secara terus-menerus dalam satu jenis penggunaan lahan. Sampel dikumpulkan dari lima area pertanian, tujuh area hutan Eucalyptus , enam area hutan hujan, dan delapan area pemukiman. Perangkap lalat buah umpan jantan yang bersumber secara komersial ( https://bugsforbugs.com.au/product/fruit-fly-trap/ ) digantung antara tujuh hingga 14 hari (jumlah hari yang sama dalam periode waktu pengambilan sampel, jumlah hari yang berbeda antara periode waktu pengambilan sampel) untuk menangkap B. tryoni di setiap lokasi pengambilan sampel. Sampel diawetkan dalam propilen glikol saat berada di perangkap dan kemudian dipindahkan ke etanol 100% dan disimpan pada suhu -20°C. Sampel diidentifikasi menurut fitur diagnostik yang dijelaskan dalam The Australian Handbook for the Identification of Fruit Flies v3.1 (Plant Health Australia 2018 ).
2.2 Ekstraksi DNA dan Genotyping
DNA diekstraksi dari 545 sampel B. tryoni jantan (188 sampel dari April 2021, 182 sampel dari Agustus 2021, dan 175 sampel dari Desember 2021) menggunakan kit Qiagen DNeasy Blood and Tissue sesuai dengan protokol pabrik pembuatnya. Langkah elusi akhir dimodifikasi dengan memanaskan buffer AE hingga 60°C dan menginkubasi buffer pada kolom spin selama 5 menit sebelum spin down akhir. DNA yang diekstraksi diperiksa kualitas dan konsentrasinya pada gel agarosa yang diwarnai dengan gel merah dan menggunakan spektrofotometer nanodrop. DNA dikirim ke Diversity Arrays Technology (DArT) untuk genotipe kepadatan tinggi DArTseq. Kombinasi enzim restriksi yang digunakan oleh DArT adalah PstI/SphI. SNP dipanggil berdasarkan jalur DArT milik sendiri (Tomkowiak et al. 2022 ) dan dipetakan ke genom referensi B. tryoni , CSIRO_BtryS06_freeze2 (tersedia di NCBI dengan nomor BioProject PRJNA560467).
2.3 Genetika Populasi
Paket dartR v2.7.2 (Gruber et al. 2018 ; Mijangos et al. 2022 ) digunakan untuk menyaring SNP, mempertahankan situs biallelik dengan rasio panggilan > 95% dan frekuensi alel minor > 0,01. Selain itu, kami menyaring SNP yang menyimpang dari ekspektasi keseimbangan Hardy–Weinberg (HWE) dalam setiap populasi secara terpisah karena hal ini telah terbukti mendeteksi struktur populasi dengan lebih andal daripada metode penyaringan HWE alternatif (Pearman et al. 2022 ). Selain itu, kami menggunakan dartR untuk memperkirakan parameter genetik populasi spesifik situs pada setiap waktu pengambilan sampel termasuk heterozigositas SNP yang diamati (H O ), heterozigositas SNP yang diharapkan (H E ), koefisien perkawinan sedarah (F IS ) dan melakukan analisis koordinat utama diferensiasi genetik individu (PCoA). Kami menggunakan analisis diskriminan komponen utama (DAPC), yang diimplementasikan dalam paket R adegenet v2.1.3 (Jombart 2008 ; Jombart et al. 2010 ) untuk memperkirakan jumlah populasi dalam data kami menggunakan nilai K dari 1 hingga 20. Nilai optimal untuk K diperkirakan menggunakan Kriteria Informasi Bayesian (BIC). Kami juga menggunakan fastStructure (Raj et al. 2014 ) untuk menilai secara independen jumlah klaster menggunakan rentang K = 1–10. Kami menggunakan program utilitas chooseK.py yang disediakan oleh fastStructure untuk menentukan nilai optimal untuk K . Barplot untuk berbagai skenario klaster dibuat menggunakan pophelper v2.3.1 (Francis 2017 ) di R (Tim Inti R 2022 ). Kami menggunakan paket R SNPRelate v1.32.2 (Zheng et al. 2012 ; Zheng et al. 2017 ) untuk menghitung nilai kekerabatan berpasangan antara individu menggunakan metode estimasi kemungkinan maksimum dan 1000 permutasi. Untuk analisis hilir, kami kemudian menghitung kekerabatan rata-rata untuk semua perbandingan berpasangan antara lokasi pengambilan sampel dan mengecualikan semua perbandingan berpasangan antara lokasi yang memiliki kurang dari lima sampel. Uji Mantel digunakan untuk memeriksa pola isolasi berdasarkan jarak untuk setiap periode waktu pengambilan sampel, menggunakan kekerabatan rata-rata (ditransformasikan menjadi 1—kekerabatan rata-rata) sebagai pengukuran ketidakmiripan genetik. AMOVA dilakukan menggunakan paket R poppr v2.9.3 (Kamvar et al. 2014 , 2015 ) untuk memperkirakan variasi genetik yang terkandung dalam dan di antara populasi menggunakan kekerabatan (ditransformasikan menjadi 1—kekerabatan) sebagai variabel jarak. Ukuran populasi efektif (N e ) diperkirakan menggunakan metode ketidakseimbangan hubungan yang diterapkan dalam NeEstimator V2 (Do et al. 2013 ) untuk setiap lokasi pengambilan sampel pada setiap waktu pengambilan sampel.
2.4 Genetika Lanskap
Sebelas permukaan resistensi (RS) dikembangkan di ArcGIS Pro v2.9.1 menggunakan prediktor lanskap yang dianggap memiliki potensi untuk memengaruhi pergerakan B. tryoni (Clarke et al. 2011 ; Clarke 2019 ; van Klinken et al. 2019 ), dan karenanya aliran gen, di seluruh area studi. Alat geoprosesing ‘Minimum Bounding Geometry (Data Management)’ digunakan untuk menentukan bentuk untuk setiap RS, dan buffer 1 km diterapkan untuk membatasi jumlah jalur yang dapat disertakan dalam pemodelan RS dalam analisis. Karena analisis IBR menggabungkan semua jalur yang mungkin antara perbandingan berpasangan (McRae 2006 ; McRae et al. 2008 ) pemotongan luas setiap RS dilakukan untuk mengecualikan jalur penyebaran yang secara biologis tidak masuk akal selama optimasi (Cameron et al. 2019 ). Kami mengembangkan delapan RS biner kategoris yang mewakili keberadaan/ketiadaan area hortikultura, jalan raya, area penggembalaan, jalan dan rel kecil, area hutan hujan, hutan Eucalyptus , jalur air, dan area pemukiman. Setiap RS dikembangkan dengan merasterisasi data vektor yang diambil dari katalog Queensland Spatial—QSpatial (lihat Lampiran S1–S14 untuk metadata lapisan spasial dan lokasi pengambilan). Setiap raster dikembangkan pada ukuran sel 500 m. Tiga RS berkelanjutan dikembangkan: indeks kebasahan topografi (TWI), elevasi, dan lereng. RS Kemiringan dikembangkan berdasarkan raster elevasi kami dengan menggunakan alat Kemiringan (Analis Spasial) di ArcGIS Pro dan menggunakan metode geodesik dengan pengukuran keluaran yang ditetapkan ke derajat. Kemudian kami mengubah resolusi spasial dari 25 m menjadi 500 m menggunakan alat resampling dan teknik resampling bilinear.
Kami menggunakan pendekatan IBR untuk menilai pengaruh struktur lanskap pada struktur populasi B. tryoni . Kami menggunakan paket R ResistanceGA v4.2–10 (Peterman 2018 ) untuk mengoptimalkan RS menggunakan CIRCUITSCAPE di Julia (Hall et al. 2021 ) menggunakan mean kinship (ditransformasikan menjadi 1-mean kinship) sebagai variabel respons, dan skema koneksi delapan tetangga. ResistanceGA menggunakan algoritma genetik untuk melakukan pengoptimalan model secara berulang hingga dukungan model tidak membaik di 25 iterasi berikutnya. Kami melakukan ini sebagai analisis dua langkah: yang pertama adalah analisis univariat dengan setiap RS dioptimalkan secara terpisah, yang kedua adalah analisis multivariat di mana model univariat dengan kinerja terbaik dioptimalkan secara bersamaan dengan RS tambahan untuk membuat RS komposit. Kinerja model diperingkat menggunakan Kriteria Informasi Akaike yang dikoreksi untuk ukuran sampel terbatas (AICc). Paket R qpcR (Ritz dan Spiess 2008 ) digunakan untuk menghitung bobot Akaike ( ω ) untuk setiap model.
2.5 Ekologi Lanskap
Kami menggunakan alat Empirical Bayesian Kriging (Geostatistical Analyst Tools) yang diimplementasikan dalam ArcGIS Pro untuk menginterpolasi kelimpahan B. tryoni di seluruh area studi. Kami melakukan Tukey HSD (Miller 1981 ; Yandell 1997 ) dalam R untuk menentukan apakah kelimpahan B. tryoni bervariasi antara jenis penggunaan lahan dan di seluruh periode pengambilan sampel. Selain itu, kami menggunakan pemodelan campuran linier (LMM) untuk menyelidiki prediktor lingkungan teratas dari kelimpahan B. tryoni . Analisis LMM menggabungkan informasi kategori penggunaan lahan yang terkandung dalam lapisan spasial yang diambil dari Qspatial, dan ini digabungkan menjadi lima kategori: area penggembalaan, area perumahan, hutan Eucalyptus , hutan hujan dan produksi pertanian, dan persentase area yang tercakup untuk setiap jenis penggunaan lahan dalam radius 1 km dari setiap situs dihitung. Variabel lain termasuk jarak dari setiap situs ke sumber air abadi terdekat, dan lintang dan bujur setiap situs. Lintang dan bujur disertakan untuk menguji potensi efek tingkat populasi AWM. Secara khusus, kami berhipotesis bahwa populasi lalat harus lebih tinggi di dalam dan di sekitar area produksi Bundaberg (di timur laut area pengambilan sampel kami) daripada area Gayndah/Mundubbera (di barat daya area pengambilan sampel kami) (Gambar 1 ) karena ada atau tidaknya AWM. Kelimpahan lalat buah Queensland diubah menjadi tangkapan per unit usaha (CPUE) di mana unit usaha dianggap sebagai 1 hari penangkapan. Situs Tansey 1 dihilangkan dari analisis LMM karena jarak antara Tansey 1 dan Tansey 2 hanya 1 km dan akan melanggar asumsi independensi spasial antara lokasi pengambilan sampel.
Kami menggunakan paket R lme4 versi 1.1–30 (Bates et al. 2015 ) dan paket R MuMIn versi 1.47.1 (Barton 2022 ) untuk menilai semua kombinasi variabel prediktor, menggunakan waktu pengambilan sampel sebagai variabel acak. Fungsi ‘dredge()’ digunakan untuk memberi peringkat semua LMM menurut kriteria informasi Akaike yang dikoreksi untuk ukuran sampel kecil (AICc). Setelah pengerukan, LMM dengan kinerja terbaik menurut dukungan AICc digunakan untuk melakukan perataan model, yang diimplementasikan dalam fungsi ‘model.avg.()’. Ini menilai setiap variabel dari model dengan kinerja terbaik untuk koefisien linier, kesalahan standar, signifikansi statistik, dan interval kepercayaan 95% atas dan bawah.
3 Hasil
3.1 Struktur Populasi
DArTseq menghasilkan total 53.368 SNP, dengan 3759 SNP dipertahankan setelah penyaringan untuk tingkat panggilan (> 0,95), MAF (> 0,01) dan penyimpangan dari HWE. Kumpulan data SNP yang dihasilkan memiliki kedalaman sekuensing rata-rata 113× (kesalahan standar ± 1,8), kedalaman sekuensing minimum 5×, dan maksimum 537×. H O di setiap situs di tiga periode pengambilan sampel berkisar antara 0,092 hingga 0,119, H E berkisar antara 0,094 hingga 0,123, dan nilai F IS berkisar antara 0,005 hingga 0,136 (Lampiran S2 ). Analisis diskriminan komponen utama (DAPC) menunjukkan nilai optimal K = 1 (Lampiran S3 ). FastStructure mengidentifikasi nilai optimal untuk K berada dalam rentang K = 1–6 (kompleksitas model yang memaksimalkan kemungkinan marginal = 1 dan komponen model yang digunakan untuk menjelaskan struktur dalam data = 6); namun, plot batang untuk skenario K = 2–6 tidak memberikan interpretasi yang bermakna secara biologis dari struktur genetik B. tryoni (Lampiran S4 ), yang menunjukkan sampel berasal dari satu populasi tanpa substruktur yang terdeteksi. Analisis koordinat utama (PCoA) mendukung K = 1, karena tidak ada pengelompokan yang terkait dengan lokasi (Gambar 2A ) atau waktu pengumpulan (Gambar 2B ) yang diamati. Dua sumbu pertama PCoA, PC1 dan PC2, hanya mencakup 1,4% dari total variasi. AMOVA juga mendukung K = 1, dengan 99,96% variasi ditemukan dalam populasi dan hanya 0,04% variasi di antara populasi (Lampiran S5 ). Estimasi N e tidak terbatas untuk semua lokasi pengambilan sampel pada setiap periode pengambilan sampel. Hanya Childers pada bulan Desember yang memiliki interval kepercayaan bawah tak terhingga (LCI = 2,2; Lampiran S6 ). Estimasi N e tetap tinggi jika mempertimbangkan semua periode waktu secara bersamaan; hanya Bundaberg yang memiliki N e tak terhingga (N e = 706,8), dan hanya Howard yang memiliki interval kepercayaan bawah tak terhingga (LCI = 11.411,6). Semua interval kepercayaan atas tak terhingga.

Nilai kekerabatan berpasangan rata-rata antara lokasi berkisar dari 0,002 hingga 0,008 Lampiran S7 dan dalam lokasi berkisar dari 0,000 hingga 0,007 (Lampiran S7 ) yang menunjukkan tidak ada struktur genetik yang signifikan di seluruh area studi atau waktu pengambilan sampel. Nilai kekerabatan berpasangan antara individu berkisar dari 0,000 hingga 0,146 (Lampiran S7 ) yang menunjukkan keberadaan saudara tiri yang diduga dalam set data. Ada enam set saudara tiri yang diduga, yang masing-masing berkaitan dengan perbandingan individu dari lokasi pengambilan sampel yang berbeda, tetapi dalam periode pengambilan sampel yang sama. Uji Mantel menemukan korelasi yang signifikan secara statistik, tetapi lemah antara jarak geografis dan kekerabatan selama bulan Agustus (Lampiran S8 dan S9 ).
3.2 Genetika Lanskap
Analisis lanskap-genetik secara lemah mendukung beberapa model berbeda sebagai prediktor aliran gen B. tryoni di seluruh area studi. Prediktor dengan kinerja terbaik untuk bulan April adalah area penggembalaan—keberadaan area penggembalaan diberi nilai resistansi optimal sebesar 21,97 (Lampiran S10 ), yang menunjukkan bahwa itu merupakan penghalang aliran gen. Model dengan kinerja terbaik pada bulan Agustus adalah area pemukiman, meskipun model jarak saja didukung dengan cara yang sama (Tabel 1 ) yang sesuai dengan hasil dari uji Mantel untuk periode pengambilan sampel ini. Beberapa model lain, termasuk model nol tanpa struktur geografis, memiliki ∆AICc < 2 pada bulan Agustus, yang menunjukkan beberapa dukungan untuk model alternatif juga. Namun, nilai resistansi optimal untuk semua RS kategoris pada bulan Agustus sangat rendah, yang menunjukkan bahwa resistansi yang dikenakan oleh ada/tidaknya setiap fitur yang dimodelkan memiliki sedikit dampak pada aliran gen. Model univariat dengan kinerja terbaik untuk semua periode waktu yang digabungkan adalah model jarak saja, meskipun model nol menunjukkan dukungan yang serupa meskipun lebih rendah (Lampiran S10 ). Ini juga didukung oleh uji mantel untuk semua periode waktu ( p = 0,07; Lampiran S8 dan S9 ). Hanya dua model dengan ∆AICc < 2 dalam analisis univariat di bulan April, Agustus, dan semua periode pengambilan sampel digabungkan yaitu model nol tanpa struktur geografis dan model area penggembalaan; selain itu, model area penggembalaan yang dioptimalkan selalu menjadi penghalang aliran gen, yang menunjukkan aliran gen B. tryoni lebih tinggi tanpa adanya area penggembalaan. Peta terkini yang dibuat berdasarkan RS area penggembalaan yang dioptimalkan untuk setiap periode waktu menunjukkan potensi laju aliran gen yang lebih tinggi di sisi timur area studi (Gambar 3 ). RS area penggembalaan adalah satu-satunya model univariat yang dimasukkan dalam semua RS komposit multivariat karena dukungan yang tidak konsisten untuk RS univariat lainnya. Hanya RS univariat TWI yang dikalahkan oleh semua model multivariat pada bulan April; tidak ada model multivariat yang mengungguli model univariat lainnya pada bulan Agustus, dan model dengan kinerja terbaik dalam analisis sepanjang periode adalah permukaan multivariat area penggembalaan dan jalan raya (Tabel 1) .). Nilai resistansi yang dioptimalkan untuk area penggembalaan dan jalan raya mengidentifikasi area penggembalaan (resistansi = 56,49) dan jalan raya (resistansi = 500,48) sebagai penghalang kuat terhadap aliran gen. Dalam RS komposit ini, kontribusi relatif dari model univariat area penggembalaan (kontribusi relatif = 87%) jauh lebih tinggi daripada model univariat jalan raya (kontribusi relatif = 13%). Tidak ada RS multivariat lain dalam analisis semua periode waktu yang mengungguli model univariat mana pun. Tidak ada model lanskap yang dapat ditentukan untuk Desember karena korelasi negatif antara variabel respons genetik dan jarak geografis. Karena dukungan yang umumnya lemah yang ditunjukkan untuk model berbasis jarak dalam pengujian Mantel dan analisis IBR kami, kami menduga korelasi negatif antara kekerabatan dan jarak pada bulan Desember adalah artefak statistik.
| Model | ∆AICc | ω | Bahasa Inggris: K | |
|---|---|---|---|---|
| April | Merumput | 0.00 | 0.249 | 3 |
| Batal | -0,20 | 0.226 | 1 | |
| Lereng | -1,31 | 0.130 | 4 | |
| Jarak | -2,83 | 0,061 tahun | 2 | |
| Daerah pemukiman | -3,07 | 0,054 tahun | 3 | |
| Jalan raya dan rel | -4,02 | 0,033 | 3 | |
| Ketinggian | -4,08 | 0,032 | 4 | |
| Hortikultura | -4,11 | 0,032 | 3 | |
| Saluran air | -4,19 | 0,031 | 3 | |
| Jalan Raya | -4,35 | 0,028 | 3 | |
| Kayu putih | -4,37 | 0,028 | 3 | |
| Hutan hujan | -4,47 | 0,027 | 3 | |
| Penggembalaan + hutan hujan | -6,43 | 0,010 | 5 | |
| Penggembalaan + jalan raya | -6,45 | 0,010 | 5 | |
| Penggembalaan + hortikultura | -6,81 | 0,008 | 5 | |
| Penggembalaan + jalur air | -6,81 | 0,008 | 5 | |
| Merumput + Eucalyptus | -6,95 | 0,008 | 5 | |
| Penggembalaan + jalan raya dan rel | -6,96 | 0,008 | 5 | |
| Penggembalaan + perumahan | -7,00 | 0,008 | 5 | |
| Dua Belas | -8,21 | 0,004 tahun | 4 | |
| Penggembalaan + lereng | -8,29 | 0,004 tahun | 6 | |
| Penggembalaan + elevasi | -11,09 | 0,001 | 6 | |
| Merumput + TWI | -11,73 | 0,001 | 6 | |
| Agustus | Daerah pemukiman | 0.00 | 0,176 tahun | 3 |
| Jarak | -0,16 | 0.162 | 2 | |
| Batal | -1,06 | 0.103 | 1 | |
| Jalan Raya | -1,08 | 0.102 | 3 | |
| Hortikultura | -1,32 | 0,091 tahun | 3 | |
| Merumput | -1,43 | 0,086 tahun | 3 | |
| Jalan raya dan rel | -1,73 | 0,074 tahun | 3 | |
| Kayu putih | -2,21 | 0,058 | 3 | |
| Saluran air | -2,76 | 0,044 tahun | 3 | |
| Hutan hujan | -2,77 | 0,044 tahun | 3 | |
| Ketinggian | -4,92 | 0,015 | 4 | |
| Dua Belas | -5,52 | 0,011 | 4 | |
| Lereng | -5,55 | 0,011 | 4 | |
| Penggembalaan + hortikultura | -7,33 | 0,004 tahun | 5 | |
| Penggembalaan + jalur air | -7,33 | 0,004 tahun | 5 | |
| Penggembalaan + perumahan | -7,50 | 0,004 tahun | 5 | |
| Penggembalaan + jalan raya | -8,34 | 0,003 | 5 | |
| Merumput + Eucalyptus | -8,74 | 0,002 | 5 | |
| Penggembalaan + hutan hujan | -8,89 | 0,002 | 5 | |
| Penggembalaan + jalan raya dan rel | -8,91 | 0,002 | 5 | |
| Merumput + TWI | -12,06 | 0.000 | 6 | |
| Penggembalaan + lereng | -12,56 | 0.000 | 6 | |
| Penggembalaan + elevasi | -12,67 | 0.000 | 6 | |
| Semua periode waktu | Penggembalaan + jalan raya | 0.00 | 0.751 | 5 |
| Jarak | -5,63 | 0,045 pukul 0,045 | 2 | |
| Batal | -6,17 | 0,034 tahun | 1 | |
| Hortikultura | -6,19 | 0,034 tahun | 3 | |
| Jalan Raya | -6,48 | 0,029 | 3 | |
| Merumput | -7,60 | 0,017 tahun | 3 | |
| Jalan raya dan rel | -7,91 | 0,014 tahun | 3 | |
| Hutan hujan | -8,15 | 0,013 | 3 | |
| Saluran air | -8,17 | 0,013 | 3 | |
| Kayu putih | -8,20 | 0,012 | 3 | |
| Perumahan | -8,20 | 0,012 | 3 | |
| Ketinggian | -9,51 | 0,006 | 4 | |
| Lereng | -10,47 | 0,004 tahun | 4 | |
| Dua Belas | -11,03 | 0,003 | 4 | |
| Penggembalaan + perumahan | -11,17 | 0,003 | 5 | |
| Penggembalaan + jalan raya dan rel | -11,41 | 0,003 | 5 | |
| Penggembalaan + jalur air | -12.10 | 0,002 | 5 | |
| Penggembalaan + hortikultura | -12,14 | 0,002 | 5 | |
| Penggembalaan + hutan hujan | -13,44 | 0,001 | 5 | |
| Merumput + Eucalyptus | -13,88 | 0,001 | 5 | |
| Penggembalaan + elevasi | -16,33 | 0.000 | 6 | |
| Penggembalaan + lereng | -16,53 | 0.000 | 6 | |
| Merumput + TWI | -16,92 | 0.000 | 6 |

3.3 Ekologi Lanskap
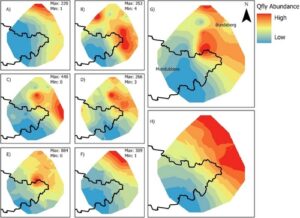
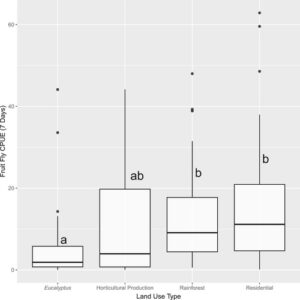
Sebanyak 13.583 B. tryoni tertangkap selama penelitian ini. Kriging Bayesian empiris terhadap kelimpahan B. tryoni untuk semua periode waktu menunjukkan kecenderungan kelimpahan tinggi di timur laut hingga kelimpahan rendah di barat daya area penelitian (Gambar 4 dan Lampiran S11 ). Pola ini terlihat jelas di seluruh enam periode pengambilan sampel, meskipun dengan sedikit fluktuasi di lokasi dengan kelimpahan tertinggi. Lokasi Good Night 2 pada bulan Februari 2022 merupakan outlier dengan tangkapan per unit usaha sebesar 126,3. Tangkapan tertinggi kedua per unit usaha kurang dari setengah nilai tersebut, sebesar 62,9 untuk Gin Gin pada Oktober 2021. Uji Tukey HSD mendeteksi perbedaan yang signifikan ( p < 0,05) antara jumlah lalat yang ditangkap di hutan Eucalyptus dan hutan hujan serta area pemukiman (Lampiran S12 dan S13 ): Lebih jauh lagi, ketika tangkapan outlier pada Good Night 2 dihilangkan, perbedaan yang signifikan secara statistik antara kelimpahan lalat buah dan habitat tangkapan tidak berubah (Gambar 5 dan Lampiran S14 ). Tidak ada perbandingan berpasangan lainnya yang berbeda secara signifikan ( p > 0,05). Analisis selanjutnya dalam hasil tersebut menghilangkan lokasi outlier Good Night 2, Februari 2022.
Analisis LMM mencakup total 512 kombinasi prediktor lanskap. Perataan model dilakukan menggunakan LMM dengan ∆AICc < 2 dari model teratas (Tabel 2 ). Model terbaik mempertahankan lima prediktor (Tabel 3 ) dari sembilan yang disertakan dalam analisis. Model dengan kinerja terbaik adalah kombinasi area penggembalaan dan jarak dari sumber air terdekat dengan keduanya berkorelasi negatif dengan kelimpahan B. tryoni . Model peringkat kedua adalah model univariat yang hanya mempertimbangkan area penggembalaan, yang memiliki korelasi negatif dengan kelimpahan B. tryoni . Model peringkat ketiga, keempat dan kelima mencakup berbagai kombinasi area penggembalaan dan variabel tambahan (lintang, bujur dan elevasi). Setelah perataan model, area penggembalaan adalah satu-satunya prediktor lanskap yang signifikan secara statistik dari kelimpahan B. tryoni (Tabel 3 ) dan berkorelasi negatif. Temuan ini konsisten dengan hasil analisis genetik lanskap di mana area penggembalaan adalah RS yang berasal dari lanskap dengan kinerja terbaik.
| Model | ∆AICc | ω | df |
|---|---|---|---|
| Penggembalaan + sumber air | 0.00 | 0.22 | 5 |
| Merumput | 0,01 | 0.22 | 4 |
| Merumput + garis lintang | 0,45 | 0.18 | 5 |
| Merumput + garis bujur | 1.40 | 0.11 | 5 |
| Penggembalaan + sumber air + garis lintang | 1.56 | 0.10 | 6 |
| Merumput + Lintang + Bujur | 1.82 | 0,09 | 6 |
| Penggembalaan + sumber air + ketinggian | 1.91 | 0,08 | 6 |
Catatan: Model diperingkat berdasarkan AIC yang dikoreksi untuk ukuran sampel terbatas (∆AICc) dan bobot Akaike ( ω ). Derajat kebebasan (df) untuk setiap model juga diberikan.
| Variabel | Koefisien | Kesalahan standar | Bahasa Indonesia: Z | P | 2,5% CI | 97,5% CI |
|---|---|---|---|---|---|---|
| (Mencegat) | -31,32 | 247.3 | 0,125 | 0.900 | -520,9 | 458.3 |
| Merumput | -0,132 | 0,388 | 3.352 | 8 × 10−4 | -0,209 | -0,055 |
| Sumber Air | -0,006 | 0,010 | 0.608 | 0.543 | -0,035 | 0,006 |
| Lintang | 0.126 | 2.384 | 0,525 | 0.600 | -2,181 | 9.082 |
| Garis bujur | 0.523 | 1.604 | 0.323 | 0.747 | -2.775 | 8.063 |
| Ketinggian | 8 × 10−4 | 0,005 | 0.163 | 0.870 | -0,019 | 0,038 |
Catatan: Koefisien linier, kesalahan standar (kesalahan standar), skor Z, nilai p , interval kepercayaan (CI) 2,5% dan 97,5% dilaporkan. Teks tebal menunjukkan signifikansi statistik dalam model dan CI yang tidak tumpang tindih dengan 0.
4 Diskusi
Analisis gabungan kami tentang ekologi lanskap dan genetika lanskap B. tryoni menemukan hubungan baru antara variabel lanskap dan kelimpahan atau aliran gen sambil juga mengonfirmasi aspek biologi yang diketahui. Area lahan penggembalaan berasosiasi negatif dengan kelimpahan B. tryoni , yang sesuai dengan prediksi model kesesuaian lanskap untuk B. tryoni yang dibangun menggunakan pendapat ahli (van Klinken et al. 2019 ). Namun, lahan penggembalaan dapat bertindak sebagai penghalang penyebaran sebelumnya tidak diketahui dan memiliki implikasi signifikan terhadap pengelolaan. Bahwa tidak ada struktur populasi geografis yang dapat dideteksi dalam kisaran endemik B. tryoni telah ditetapkan sebelumnya (Popa-Báez et al. 2020 ). Namun, metode keterkaitan lanskap berbasis kekerabatan yang digunakan di sini memberikan data baru tentang penyebaran pada spesies yang sangat melimpah dengan kapasitas penyebaran yang tinggi. Akhirnya, kami membahas dampak potensial AWM dalam mengurangi kelimpahan populasi B. tryoni , yang menunjukkan nilai pendekatan ini untuk pengelolaan serangga hama. Poin-poin ini dikembangkan lebih lanjut di bawah bagian genetika lanskap, ekologi lanskap, dan implikasinya terhadap pengelolaan hama.
4.1 Genetika Lanskap
Beberapa penelitian telah mengevaluasi struktur genetik B. tryoni pada skala spasial yang besar (multinegara bagian hingga benua), tetapi tidak ada yang meneliti keterkaitan genetik dalam mosaik hortikultura pada skala lanskap. Sementara penelitian sebelumnya telah mendeteksi penataan genetik populasi di Australia selatan, tempat populasi B. tryoni baru-baru ini invasif (Gilchrist et al. 2006 ; Gilchrist dan Meats 2010 ; Blacket et al. 2017 ), penataan populasi dalam rentang endemik tropis dan subtropis B. tryoni hampir seluruhnya tidak ada (Gilchrist et al. 2006 ; Cameron et al. 2010 ; Popa-Báez et al. 2020 ; Parvizi et al. 2024 ). Secara keseluruhan, data genetik skala halus kami dalam wilayah kecil rentang endemik B. tryoni mendukung temuan penelitian sebelumnya ini, dengan struktur genetik terbatas yang terbukti di antara area yang diambil sampelnya dengan padat. Namun, studi kami bersifat baru karena dengan menggunakan desain pengambilan sampel sistematis di seluruh lanskap, kami telah mampu mengidentifikasi lahan penggembalaan sebagai penghalang aliran gen pada spesies hama ini. Studi kami menyoroti nilai pemodelan resistensi lanskap untuk mendeteksi penghalang atau fasilitator aliran gen bahkan pada spesies yang menunjukkan struktur genetik terbatas (misalnya, Pavlacky Jr et al. 2009 ; Verba et al. 2023 ). Sebelumnya, satu-satunya penghalang aliran gen telah ditemukan di antara populasi invasif B. tryoni yang terpisah di Alice Springs, Victoria, dan di Kepulauan Pasifik (Popa-Báez et al. 2020 ). Lebih jauh lagi, studi kami juga menyoroti potensi pendekatan genomik lanskap untuk menginformasikan pengelolaan spesies serangga hama dengan lebih baik melalui penerapan AWM untuk membatasi aliran gen dan untuk mengeksploitasi penghalang alami dalam penerapan pelepasan teknik serangga mandul strategis (SIT) (Schmidt et al. 2018 , 2023 ).
Bahasa Indonesia: Merancang strategi manajemen untuk spesies hama sering kali bergantung pada kemampuan kita untuk melacak pergerakan tingkat individu dan populasi di seluruh lanskap. Saat ini, nilai data genetik untuk melacak secara langsung pergerakan B. tryoni dan tephritid lainnya di tingkat lanskap masih belum jelas. Mikrosatelit (Gilchrist dan Meats 2010 ) dan data sekuens mitokondria (Blacket et al. 2017 ) cukup untuk mengidentifikasi dan melacak populasi B. tryoni yang berdekatan secara geografis di garis depan invasi selatan di mana ukuran populasi efektifnya kecil. Namun, untuk B. tryoni dan lalat melon, Zeugodacus cucurbitae (Coquillet), analisis SNP genomik telah mendeteksi penataan genetik minimal dalam rentang habitat endemik mereka di Australia timur (Popa-Báez et al. 2020 ) dan Asia Tenggara (Dupuis et al. 2018 ), meskipun populasi invasif dapat diidentifikasi. Dalam studi ini, kami menemukan bahwa ukuran populasi efektif B. tryoni sangat besar di wilayah asli, dan ini mengaburkan deteksi struktur populasi skala halus. Namun, ukuran keterkaitan lanskap (Norman et al. 2017 ), menggunakan nilai kekerabatan, telah terbukti mengungguli metode lain untuk mendeteksi struktur populasi (misalnya, F ST dan algoritma pengelompokan) ketika ukuran populasi efektif mendekati tak terhingga, atau ketika struktur populasi lemah (Iacchei et al. 2013 ; Palsbøll et al. 2010 ). Oleh karena itu, kami berhipotesis bahwa pendekatan keterkaitan lanskap dalam wilayah invasif mungkin berguna untuk melacak pergerakan B. tryoni secara langsung di tingkat lanskap dan membantu merancang manajemen untuk serangga hama. Waktu pengambilan sampel juga dapat memengaruhi hasil investigasi genetik karena berbagai RS menunjukkan berbagai tingkat dukungan tergantung pada waktu pengambilan sampel. Ini mungkin artifaktual mengingat struktur genetik yang sangat lemah yang ditunjukkan di semua periode waktu. Sebagai alternatif, kapasitas untuk mendeteksi struktur genetik (jika ada) dapat ditingkatkan sebelum perpindahan antarpopulasi ketika individu yang lebih dekat hubungannya berada dalam jarak geografis yang lebih dekat, yaitu, jika pengambilan sampel dilakukan pasca-musim dingin, ketika jumlah populasi B. tryoni mulai meningkat (Clarke et al. 2022 ). Hasil kami menunjukkan beberapa dukungan untuk hal ini, dengan Agustus (bulan paling awal dalam setahun ketika jumlah populasi mulai meningkat) menunjukkan dukungan yang lebih kuat untuk IBD daripada April.
Jalan raya muncul sebagai kemungkinan penghalang aliran gen dalam analisis multivariat kami. Mungkin penghalang ini hanya muncul dalam konteks area penggembalaan karena jalan raya adalah bentuk ekstrem dari lahan terbuka. Namun, studi simulasi telah menunjukkan bahwa ketika tingkat ‘kebisingan’ yang tinggi diperkenalkan ke dalam variabel respons genetik, analisis multivariat dapat memiliki tingkat kesalahan tipe I yang tinggi (Winiarski et al. 2020 ). Karena struktur genetik lemah yang kami amati, data kami mungkin memiliki ‘kebisingan’ yang cukup bagi MLPE untuk menyesuaikan secara berlebihan model komposit area penggembalaan dan jalan raya sepanjang periode waktu. Jika pola ini bukan kesalahan tipe I, kepentingan relatif yang tinggi dari area penggembalaan dalam model komposit semakin mendukung perannya sebagai penghalang aliran gen dalam studi kami.
4.2 Ekologi Lanskap
Pendapat ahli yang signifikan ada tentang ekologi lanskap B. tryoni , dan ini ditangkap dalam model risiko lanskap oleh van Klinken et al. Pendapat ahli itu percaya lalat akan berlokasi di lokasi yang terlindung dan lembab dengan akses yang baik ke buah untuk berkembang biak. Namun, selain studi saat ini, hanya Fletcher ( 1974a , 1974b ) di Australia beriklim sedang dan Raghu et al. ( 2000 ) di Australia subtropis yang secara bersamaan mengambil sampel beberapa tipe habitat sehingga dapat langsung membandingkan penggunaan habitat. Fletcher ( 1974b ) dan Raghu et al. ( 2000 ), dan seperti yang dimodelkan oleh van Klinken et al. ( 2019 ) dan Schwarzmueller et al. 2019 , menemukan kelimpahan B. tryoni tinggi di lokasi perkotaan. Pasokan buah hampir sepanjang tahun di halaman belakang perkotaan dan penyiraman yang sering mengakibatkan kelembaban lokal yang tinggi dianggap menjelaskan pola ini (Dominiak et al. 2006 ; Schwarzmueller et al. 2019 ). Namun, sementara kami menemukan B. tryoni secara numerik paling melimpah di perangkap perkotaan, itu tidak secara signifikan lebih banyak daripada hutan hujan dan lokasi produksi hortikultura. Sementara pentingnya area hortikultura untuk perkembangbiakan B. tryoni terbukti dengan sendirinya dari status hama, pentingnya hutan hujan sebagai tempat perkembangbiakan dan perlindungan bagi B. tryoni kurang jelas, meskipun hutan hujan dianggap sebagai tipe habitat utama untuk spesies Bactrocera Australia (Starkie et al. 2024 ). Raghu et al. ( 2000 ) menemukan B. tryoni langka di hutan hujan, sementara Drew et al. ( 1984 ) dan penelitian kami mengumpulkan sejumlah besar lalat dari hutan hujan. Ada banyak alasan mengapa hasil penelitian dapat bervariasi, tetapi dengan dua penelitian yang sekarang menangkap sejumlah besar B. tryoni di hutan hujan, harus diasumsikan bahwa ini adalah tipe habitat yang cocok untuk spesies tersebut sebagaimana halnya untuk spesies lain dalam genus tersebut. Bahwa B. tryoni dapat terperangkap di hutan eukaliptus, bahkan jika dalam jumlah rendah, diketahui dari literatur (Raghu et al. 2000 ; Fletcher 1974a , 1974b ), dengan Fletcher menyajikan bukti langsung ( 1974a ) dan tidak langsung ( 1974b ) bahwa lalat yang ditangkap di hutan eukaliptus menyebar melalui hutan ke dan dari daerah perkembangbiakan. Data penangkapan sebelumnya ini, dikombinasikan dengan data genetik dan data penangkapan kami, sangat menyimpulkan bahwa meskipun hutan eukaliptus tidak boleh dianggap sebagai habitat permanen bagiB. tryoni , bukan merupakan penghalang pergerakan. Bahwa lahan penggembalaan terbuka merupakan penghalang pergerakan B. tryoni dan berasosiasi negatif dengan kelimpahan belum teridentifikasi sebelumnya.
4.3 Implikasi terhadap Pengelolaan Hama
Bahwa AWM dapat menekan populasi hama lokal telah terdokumentasikan dengan baik (Hendrichs et al. 2007 ), seperti halnya kasus AWM B. tryoni yang berhasil di wilayah Gayndah–Mundubbera saat pertama kali diterapkan (Lloyd et al. 2007 , 2010 ). Meskipun demikian, data yang disajikan di sini adalah yang pertama untuk hortikultura Australia yang menunjukkan potensi efek lanskap AWM antara dua wilayah produksi yang berdekatan (Gambar 4 ), satu dengan AWM jangka panjang yang sekarang berlaku dan yang lainnya tanpa. Di wilayah produksi pesisir besar Bundaberg dan wilayah yang lebih kecil di selatannya, populasi lalat sangat tinggi, sementara di barat daya wilayah studi di sekitar Gayndah dan Mundubbera tempat AWM diterapkan, populasi lalat jauh lebih rendah. Program AWM untuk Gayndah–Mundubbera relatif sederhana, dengan lalat buah dikelola di lahan pertanian dan di kota-kota pedesaan (mengelola buah di pohon-pohon jalan, pohon-pohon taman, dll.), tetapi tidak di habitat alami yang menguntungkan lainnya seperti hutan hujan (Lloyd et al. 2010 ). Kebijaksanaan yang ada menunjukkan bahwa AWM kemungkinan besar akan berhasil di tempat yang populasinya relatif rendah, ada beberapa tempat berkembang biak yang tidak terkendali (yaitu, tidak ada hutan hujan), dan ada suhu rendah musiman untuk memutus siklus perkembangbiakan (Jessup et al. 2007 ). Sebaliknya, data kami menunjukkan bahwa manajemen di lahan pertanian dan kota-pedesaan saja sudah cukup untuk menekan populasi B. tryoni secara regional di seluruh lanskap subtropis yang kompleks di mana populasi besar dan suhu yang cocok terjadi sepanjang tahun, menawarkan peluang baru untuk B. tryoni AWM di daerah asalnya.
Aspek kedua dan penting dari pekerjaan kami untuk manajemen hama adalah fakta bahwa lahan penggembalaan bertindak sebagai penghalang penyebaran lalat buah dan berkorelasi negatif dengan kelimpahan lalat buah. Standar Internasional untuk Tindakan Fitosanitari (ISPM) menetapkan pedoman yang diterima secara internasional untuk mengelola risiko fitosanitari yang ditimbulkan oleh pergerakan atau perdagangan tanaman dan produk turunan tanaman. ISPM 26 ( 2006 ) dan ISPM 35 ( 2012 ) membahas proses pengurangan risiko untuk produk yang terkena dampak lalat buah melalui pembentukan area bebas hama atau penerapan pendekatan sistem (serangkaian langkah pengurangan risiko independen yang berurutan), masing-masing. Untuk kedua pendekatan, mengetahui bahwa lahan penggembalaan terbuka merupakan penghalang bagi pergerakan B. tryoni adalah penting. Area domba dan negara penggembalaan yang sangat luas umum terjadi di Australia dan area produksi, atau tempat produksi hortikultura individual (misalnya, kompleks rumah kaca besar), umumnya terletak di dalam negara penggembalaan di sekitarnya. Bertindak sebagai pencegah pergerakan dan kelimpahan lalat buah alami, lahan penggembalaan sesuai dengan ISPM 26 Bagian 2.2.1 sebagai kontribusi terhadap ‘pembentukan zona penyangga’ dan dalam ISPM 35 Bagian 2 ‘pemilihan lokasi dengan prevalensi hama rendah’.
5. Kesimpulan
Secara keseluruhan, penelitian ini menunjukkan bahwa baik AWM maupun lahan penggembalaan dapat mengurangi kelimpahan, dan dalam kasus lahan penggembalaan (dan dalam gabungan dengan jalan raya), aliran gen pada serangga hama. Temuan ini memperkuat temuan penelitian sebelumnya tetapi juga menetapkan pentingnya rezim pengambilan sampel yang direncanakan dengan cermat untuk menguji hipotesis genetik dan ekologi lanskap tertentu pada serangga hama di seluruh mosaik lanskap hortikultura. Pekerjaan di masa mendatang harus terus meneliti potensi pendekatan terkait lanskap untuk melacak wabah dan serangan di batas jangkauan pada serangga hama bergerak.


