Abstrak
- Kelebihan struktural, di mana biomassa dialokasikan secara berlebihan ke area daun pohon dibandingkan dengan area gubal, dapat mengakibatkan stres yang mematikan selama musim kemarau. Perubahan iklim dapat mengubah isyarat iklim yang mendorong produksi area daun, seperti suhu dan curah hujan, serta dinamika musiman yang mendasari curah hujan musim panas karena Monsun Amerika Utara (NAM). Jika digabungkan, hal ini dapat menyebabkan ketidaksesuaian temporal antara permintaan dan ketersediaan air yang didorong oleh area daun, dan peningkatan kejadian kematian akibat kekeringan.
- Kami menggunakan rasio luas daun terhadap luas gubal untuk menyelidiki prevalensi tunas berlebih dan apakah tunas berlebih meningkatkan kematian akibat kekeringan. Kami mengukur populasi pohon aspen yang membentang di zona transisi utara NAM selama dan setelah kekeringan parah.
- Kami mengamati peningkatan overshoot dan kematian akibat kekeringan pada populasi di garis lintang selatan yang lebih bergantung pada curah hujan musim panas. Perubahan aktivitas konvektif akibat rendahnya lapisan salju pada musim dingin sebelumnya mungkin menjadi pendorong iklim dari meningkatnya curah hujan musim panas di wilayah tersebut dan oleh karena itu juga dapat memicu peningkatan produksi luas daun selama musim panas yang lebih basah.
- Hasil penelitian kami menunjukkan bahwa kelebihan rasio luas daun terhadap luas gubal (A L :A S ) dikaitkan dengan kematian pohon akibat kekeringan dan menyoroti bahwa perubahan NAM yang disebabkan oleh perubahan iklim dapat berdampak besar pada aklimatisasi spesies pohon terhadap perubahan lingkungan.
Baca Ringkasan Bahasa Sederhana gratis untuk artikel ini di blog Jurnal.
1. PENDAHULUAN
Pergeseran dalam alokasi biomassa antara luas daun (A L ), yang mencerminkan permintaan hidrolik, dan luas gubal (A S ), yang mencerminkan kapasitas pasokan hidrolik, merupakan cara utama spesies pohon membatasi kehilangan air karena meningkatnya permintaan penguapan selama periode kekeringan. Baik variasi antar-spesies maupun dalam-spesies dalam rasio A L :A S terjadi dengan cara yang dapat diprediksi (misalnya berkurangnya A L di iklim kering), dan penyesuaian dalam A L :A S sebagai respons terhadap perubahan lingkungan biasanya terjadi pada skala waktu menengah (misalnya selama musim tanam atau selama beberapa tahun), menjadikannya sifat fungsional respons kekeringan yang utama (Anderegg et al., 2021 ; DeLucia et al., 2000 ; Gleason et al., 2013 ; Lachenbruch et al., 2021 ; Martínez-Vilalta et al., 2009 ; Mencuccini & Grace, 1995 ; Rosas et al., 2019 ; Trugman et al., 2019 ). Namun, ketidaksesuaian temporal antara permintaan dan ketersediaan air dapat menyebabkan maladaptasi terhadap kondisi lingkungan, sehingga mengakibatkan ‘kelebihan struktural’ di mana pohon membangun terlalu banyak area daun (yakni penguapan) selama periode basah relatif terhadap area gubal yang dapat memasok air ke tajuk dan kemudian akibatnya mengalami stres mematikan selama periode kekeringan (Jump et al., 2017 ; Zhang et al., 2021 ).
Perubahan iklim dapat mengubah hubungan di antara faktor-faktor iklim, seperti hubungan musiman antara suhu dan lapisan salju dengan curah hujan musim panas, yang dapat memiliki implikasi besar bagi kemampuan spesies pohon untuk beraklimatisasi terhadap perubahan lingkungan yang cepat. Di Amerika Serikat bagian barat daya (AS), Monsun Amerika Utara (NAM) menghasilkan lebih dari setengah presipitasi regional sebagai curah hujan musim panas (Adams & Comrie, 1997 ) dan menyediakan sumber air penting bagi banyak spesies pohon. Simulasi model iklim telah mendokumentasikan bahwa curah hujan NAM kemungkinan dipengaruhi oleh umpan balik daratan-atmosfer yang didorong oleh lapisan salju dan suhu, di mana lapisan salju yang rendah cenderung memicu peningkatan aktivitas konvektif dan curah hujan musim yang lebih kuat (Notaro & Zarrin, 2011 ; Zhu et al., 2005 ). Namun studi terbaru tentang pola kopling daratan-atmosfer telah mendokumentasikan melemahnya sinyal NAM di bawah skenario perubahan iklim saat ini yang dapat menyebabkan pola presipitasi musim panas yang lebih bervariasi di masa mendatang dan pelepasan umpan balik antara lapisan salju musim dingin dan presipitasi monsun musim panas, yang disebut di sini sebagai ‘pelepasan NAM’ (Pascale et al., 2017 ). Perubahan iklim juga cenderung menyebabkan lebih banyak tahun dengan lapisan salju rendah (Ashfaq et al., 2013 ), yang, dikombinasikan dengan kegagalan monsun, dapat mematikan hutan barat daya. Jika pohon tidak dapat beraklimatisasi dengan pola iklim yang berubah ini, pelepasan NAM dapat memicu overshoot struktural di hutan sebagai respons terhadap sinyal iklim yang sangat berfluktuasi, meningkatkan kematian pohon karena luas daun melebihi ketersediaan air di tegakan hutan.
Kematian yang disebabkan oleh kekeringan yang luas di banyak hutan Amerika Utara telah terjadi dalam beberapa dekade terakhir, khususnya di Amerika Serikat bagian barat daya (Anderegg et al., 2012 ; Bell et al., 2014 ; Williams et al., 2020 ). Mengingat potensi melemahnya NAM, yang menyediakan >50% air untuk sebagian besar hutan di AS bagian barat daya, memahami respons sifat fungsional dan ketidaksesuaian yang memengaruhi risiko kematian akibat kekeringan tetap menjadi area penelitian penting untuk memperkirakan pola kematian hutan di masa depan (Trugman et al., 2021 ). Hubungan yang sebelumnya didokumentasikan antara luas daun spesifik (SLA, luas daun dibagi massa) dan kematian meningkatkan kemungkinan bahwa ketidaksesuaian sifat dengan pasokan hidrolik dapat mendorong kematian (Greenwood et al., 2017 ). A L :A S juga dapat memengaruhi kematian pohon setelah kekeringan, karena A L :A S yang lebih tinggi meningkatkan permintaan evaporatif dan mengurangi kapasitas pasokan selama periode ketersediaan air yang lebih rendah. Ketidaksesuaian antara penawaran dan permintaan dapat terjadi pada skala jaringan atau skala tegakan, namun penskalaan tingkat cabang ke tingkat tegakan A L :A S tetap menjadi hal yang tidak diketahui yang penting untuk memeriksa kerentanan kekeringan tingkat populasi. Persamaan allometrik menyediakan alat untuk upaya dan model penskalaan hutan (Eagleson, 1982 ; Martin et al., 1998 ; Sperry et al., 2019 ), dan pemahaman yang lebih relevan secara ekologis tentang struktur tegakan hutan, persaingan sumber daya, dan respons pohon terhadap stres. Oleh karena itu, hipotesis penting tetapi belum teruji adalah bahwa overshoot struktural dan variasi maladaptif dalam tingkat cabang dan tegakan A L :A S (luas daun lebih banyak atau permintaan evaporatif daripada pasokan air yang tersedia) mempercepat kematian hutan selama peristiwa kekeringan yang parah.
Kami menguji hipotesis bahwa variasi maladaptif dalam A L :A S dapat mempercepat mortalitas hutan di dalam dan di antara populasi aspen ( Populus tremuloides ), spesies pohon yang tersebar luas dan penting secara ekologis di Amerika Utara yang telah mengalami stres kekeringan parah dan mortalitas akibat kekeringan regional selama dua dekade terakhir (Anderegg et al., 2013 ; Worrall et al., 2008 ). Populasi ini menjangkau zona transisi utara NAM di Utah dan Colorado. Kami bertanya: (1) Bagaimana A L :A S (baik tingkat cabang dan tegakan) bervariasi di seluruh gradien temporal dan spasial? (2) Apakah korelasi iklim antara lapisan salju musim dingin dan curah hujan musim panas (yaitu NAM) menyediakan mekanisme untuk pola produksi daun pada populasi aspen yang bergantung pada musim hujan, dan apakah NAM gagal selama tahun-tahun kekeringan parah yang memicu mortalitas aspen yang meluas akibat kekeringan? dan (3) apakah overshoot struktural luas daun terkait dengan peningkatan risiko mortalitas akibat kekeringan?
2 METODE
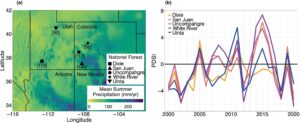
Kami mengukur dinamika luas daun aspen dan mortalitas selama dan setelah kekeringan parah pada tahun 2018 dan 2020 di lima populasi aspen alami di Hutan Nasional (NF) yang mencakup zona transisi utara NAM di Utah dan Colorado, Amerika Serikat. Ini termasuk Dixie NF, San Juan NF, Uncompahgre NF, White River NF dan Uinta NF (Gambar 1a ). Populasi paling selatan (misalnya Dixie dan San Juan) biasanya mengalami presipitasi NAM yang substansial, sedangkan sinyal NAM biasanya tidak terjadi di hutan lintang utara (misalnya White River dan Uinta). Kami juga secara intensif mempelajari dinamika luas daun di San Juan NF di seluruh ketinggian dan melalui tahun-tahun yang berbeda secara meteorologis (2014, 2018, 2019, 2020). Dalam beberapa dekade terakhir, semua populasi telah mengalami beberapa tahun stres kekeringan sebagaimana tercermin oleh nilai Indeks Keparahan Kekeringan Palmer (PDSI) (Gambar 1b ). Selama tahun-tahun kekeringan parah tahun 2018 dan 2020, nilai PDSI di seluruh populasi yang dijadikan sampel berkisar antara -3 hingga -5 dan dari -2,5 hingga -6, masing-masing (Gambar 1b ). Di dalam NF San Juan, nilai PDSI positif selama tahun 2014 dan 2019, yang menunjukkan tahun-tahun yang lebih basah daripada rata-rata (Gambar 1b , Tabel S1 ). Lihat Kerr et al. ( 2022 ) untuk informasi lebih lanjut mengenai kondisi iklim untuk plot di dalam NF San Juan.
2.1 Lokasi Lapangan
Pada tahun 2014, 15 plot studi didirikan di tegakan aspen alami di San Juan NF di Colorado Barat Daya di batas rentang elevasi bawah (rendah, 2669 m), pusat rentang (tengah, 2875 m), dan batas rentang elevasi atas (tinggi, 3080 m) dari spesies ini. Plot-plot ini dipilih untuk mempertahankan lereng dan aspek yang relatif konsisten untuk menghindari variasi mikrotopografi (Tabel S2 ). Pada tahun 2018, dua dan satu plot tambahan didirikan pada batas rentang bawah dan tengah, masing-masing, untuk menggantikan plot yang telah sepenuhnya mati sejak 2014. Pada tahun 2020, enam plot didirikan di masing-masing dari lima hutan alami di Dixie, San Juan, Uncompahgre, White River dan Uinta NF di Utah dan Colorado (Gambar 1 ; Tabel S2 ). Plot-plot ini dipilih dari daftar kandidat populasi aspen alami yang mencakup gradien iklim dari iklim dingin, basah hingga panas, kering yang telah dihasilkan dari basis data Inventarisasi dan Analisis Hutan Dinas Kehutanan Amerika Serikat. Plot studi tahun 2020 dipilih agar memiliki lereng, aspek, dan elevasi yang serupa, sehingga perbedaan iklim makro merupakan perbedaan iklim utama di antara populasi-populasi tersebut (lihat Tabel S2 untuk detail lebih lanjut tentang kondisi lokasi). Semua plot studi memiliki radius 18 m, plot melingkar, dan titik-titik pusat ditandai dengan unit GPS (Trimble Geo 7x, Trimble, Inc.). Survei plot pada tahun 2018 dan 2020 dilakukan untuk mendokumentasikan diameter pada tinggi dada (DBH) untuk setiap pohon dengan DBH > 3 cm. DBH berkisar antara 11,4 hingga 21,5 cm di seluruh plot studi kami (Tabel S2 ). Plot ditinjau ulang pada musim tanam berikutnya (2019 untuk transek elevasi San Juan dan 2021 untuk perbandingan multihutan) dan pohon-pohon dinilai secara visual untuk kesehatan dan kematian tajuk baru-baru ini (yaitu kematian). Karena 2018 dan 2020 merupakan tahun-tahun kekeringan parah bagi populasi ini, mengevaluasi kematian tajuk memberikan perkiraan yang berguna tentang bagaimana variasi sifat memediasi kematian akibat kekeringan di antara populasi alami yang dipelajari di sini.
Pusat plot studi geografis digunakan untuk mengekstrapolasi variabel iklim bulanan yang diminati pada resolusi spasial ~4 km dari tahun 1958 hingga 2020 (TerraClimate, Abatzoglou et al., 2018 ): suhu maksimum ( T maks, °C), akumulasi presipitasi (PPT, mm) dan ekuivalen air salju (SWE, cm). PPT dari tahun 1961 hingga 1990 digunakan untuk menentukan presipitasi yang terjadi sebagai curah hujan musim panas (Juli, Agustus) di wilayah plot studi kami dan Monsun Amerika Utara (NAM). Periode waktu ini ditargetkan untuk memberikan estimasi historis curah hujan musim panas sebelum kekeringan yang intens dan sering terjadi di wilayah ini dalam beberapa dekade terakhir. Tidak ada izin atau izin yang diperlukan untuk kerja lapangan dalam studi ini.
2.2 Pengukuran
Bahasa Indonesia: Di dalam setiap plot, pohon-pohon individu yang sehat dan dewasa (tiga pohon pada tahun 2014, lima di semua tahun lainnya) dipilih untuk pengukuran tingkat cabang A L :A S . Plot dikunjungi dari akhir Juni hingga awal Juli 2014, 2018, 2019 dan 2020, dan satu hingga tiga sampel cabang per pohon fokus dikumpulkan dari bagian atas tajuk yang terkena sinar matahari menggunakan senapan laras ganda kaliber 20 di dalam setiap plot. Sementara fenologi daun tidak dilacak dalam penelitian ini, aspen di wilayah tersebut telah terbukti memulai musim tanamnya pada pertengahan April (Meier et al., 2015 ), dengan perkembangan daun penuh dan pertumbuhan cabang baru terjadi selama jangka waktu pengukuran kami. Ujung cabang yang patah segera dibungkus dengan tisu basah, ditempatkan dalam kantong plastik lembab dan ke dalam pendingin untuk diangkut kembali ke laboratorium.
Segmen cabang dipotong di dalam air menggunakan pisau cukur tajam untuk menghasilkan sampel cabang dengan diameter gubal ~5 mm dan panjang ~10 cm untuk mengakomodasi panjang pembuluh (Sperry et al., 1991 ). Konduktansi hidrolik pada sampel diukur dengan metode aliran-tekanan (Sperry et al., 1988 ) dengan larutan 2% kalium klorida 0,2 μm yang disaring. Konduktansi asli ( k nat) diukur, kemudian sampel menjalani infiltrasi vakum semalam untuk menghilangkan emboli dan menentukan konduktansi maksimum ( k maks). Persentase kehilangan konduktansi (PLC), yang menunjukkan derajat emboli asli yang ada dalam sampel, dikuantifikasi oleh Persamaan ( 1 ):
![]()
Total luas daun satu sisi diukur menggunakan gambar dedaunan distal ke sampel cabang yang dipotong (menggunakan kamera digital jarak tetap pada tahun 2014, 2018, dan meteran area LI-3100C (Li-Cor Biosciences) pada tahun 2019, 2020), dan ImageJ (Schneider et al., 2012 ). Luas gubal dihitung dari diameter gubal di ujung basal segmen cabang, dan rasio A L :A S dihitung dengan membagi A L dengan A S. Pekerjaan sebelumnya di San Juan NF menunjukkan cabang aspen sebesar ini mempertahankan pertumbuhan rata-rata ~8 tahun, jadi kami tidak memperhitungkan usia bagian cabang dalam penelitian ini (Anderegg et al., 2013 ). Untuk San Juan NF, dasar untuk A L :A S diasumsikan tercermin dalam hasil tahun 2014 karena ini adalah tahun iklim rata-rata untuk plot ini. Untuk semua hutan pada tahun 2020, NF Uinta menyediakan garis dasar untuk A L :A S . Populasi ini paling tidak bergantung pada curah hujan musim panas dan memiliki nilai PDSI paling negatif pada tahun 2020 (Gambar 1b ). Oleh karena itu, nilai A L :A S pada pohon-pohon NF Uinta kemungkinan besar mewakili kombinasi permintaan daun versus pasokan yang paling tidak menimbulkan stres.
Kami juga menyelidiki apakah pola tingkat cabang ditingkatkan ke tingkat tegakan dengan menganalisis variasi pada rasio indeks luas daun (LAI) tingkat tegakan terhadap rasio luas gubal (SA) tingkat tegakan dan LAI terhadap indeks luas pangkal (BAI) tingkat tegakan. Untuk BAI, kami menggunakan BAI untuk pertumbuhan 8 tahun terakhir (BAI 8 ) mengingat cabang biasanya mempertahankan pertumbuhan ~8 tahun (lihat di atas). Pada bulan Juli 2020, satu inti riap dikumpulkan pada tinggi dada dari 15 pohon fokus dari masing-masing plot studi yang ditetapkan pada tahun 2020 ( n = 90 inti per hutan). Selama pengumpulan, kedalaman gubal (cm) dicatat dengan menandai titik di mana gubal beralih ke kayu teras. Titik ini ditentukan dengan memegang inti ke matahari, karena kayu gubal mentransmisikan lebih banyak cahaya daripada kayu teras terutama karena perbedaan kadar air (Kaufmann & Troendle, 1981 ). Di laboratorium, inti dipasang dan diampelas dengan amplas yang semakin halus hingga batas lebar cincin terlihat di bawah mikroskop. Penanggalan silang diverifikasi pada tingkat signifikansi p < 0,01 dan menggunakan jendela 50 tahun, yang tumpang tindih selama 25 tahun dalam program COFECHA (Holmes, 1983 ). Paket R ‘dplr’ (v.1.7.2) digunakan untuk mengonversi lebar cincin mentah menjadi peningkatan luas basal (BAI; Bunn, 2008 ), dan total BAI dari 8 tahun pertumbuhan terbaru (2013–2020 untuk plot studi 2020, dan 2011–2018 untuk plot San Juan NF) ditentukan. SA pertama kali dihitung menggunakan inti untuk setiap hutan dengan Persamaan ( 2 ):
![]()
di mana sd i adalah kedalaman gubal pohon ke – i dan DBH i adalah diameter pohon ke-i pada tinggi dada. Persamaan ini mengasumsikan pohon memiliki bentuk silinder. Meskipun pohon mungkin tidak berbentuk lingkaran sempurna, ini adalah asumsi standar dan dianggap cukup akurat. SA kemudian diestimasi untuk semua pohon di setiap plot studi menggunakan model regresi daya yang sesuai antara DBH pohon dan SA (Gambar S1 ; Meinzer et al., 2001 ). BAI 8 juga diestimasi untuk semua pohon di setiap plot studi menggunakan model regresi linier yang sesuai antara DBH pohon dan BAI 8 (Gambar S2 ).
Kami mengkaji luas daun tegakan secara keseluruhan menggunakan pengamatan indeks luas daun (LAI) yang lebih langsung. Pada bulan Agustus 2018, penganalisa tajuk tanaman Licor LAI-2000 digunakan untuk mengukur LAI dalam plot transek elevasi San Juan menurut metode dalam manual dengan penutup pandangan 90° yang ditempatkan di atas lensa (Licor, 1992 ). Sensor ditempatkan di bawah tajuk (~1 m di atas tanah) dan dipindahkan ke 12 titik berbeda dalam setiap plot (pusat plot dan pada 5, 10 dan 15 m di setiap arah mata angin) untuk menangkap variabilitas dalam tajuk plot. Pada bulan Juni/Juli 2020, foto hemisferis digunakan untuk mengukur LAI di semua plot hutan menggunakan perangkat lunak Gap Light Analyzer (GLA v2; Frazer et al., 1999 ). Foto diambil di 5 titik berbeda dalam setiap plot (pusat plot dan pada 10 m di setiap arah mata angin) untuk menangkap variabilitas dalam tajuk plot. Kami menggunakan variabel keluaran LAI 4-cincin yang mewakili estimasi LAI terintegrasi pada sudut zenith 0–60°. Untuk kedua metode, pengukuran dan foto LAI-2000 diambil saat fajar atau senja untuk memastikan sinar matahari langsung tidak memengaruhi estimasi LAI.
2.3 Statistik
Model efek campuran linear dibangun untuk menghubungkan nilai pengukuran absolut (A L :A S , LAI:SA, LAI, LAI:BAI 8 ) di antara populasi pada tahun 2020 (Gambar 2A,C,E ), di antara aspen San Juan NF pada tahun 2018 (Gambar 2D,F ), dan di seluruh ketinggian dan tahun di San Juan NF dari tahun 2014 hingga 2020 (Gambar 2B ). Model efek campuran linear juga dibangun untuk menghubungkan nilai pengukuran absolut PLC di antara populasi tahun 2020. Untuk analisis populasi tahun 2020, kami menggunakan efek tetap populasi dan efek acak plot. Untuk analisis San Juan NF tahun 2018, kami menggunakan efek tetap elevasi dan efek acak plot. Untuk analisis San Juan NF antartahunan, kami menggunakan efek tetap tahun, elevasi, dan interaksinya, dan efek acak pohon untuk memperhitungkan pengukuran berulang. Asumsi normalitas model diperiksa dengan plot diagnostik residual. Efek populasi, elevasi, tahun, dan interaksi elevasi-tahun ditentukan dengan uji rasio kemungkinan (likelihood ratio test/LRT) dengan membandingkan model lengkap dengan model yang diperkecil. Ketika efeknya signifikan, perbandingan berpasangan dilakukan antara kelompok menggunakan estimasi rata-rata marginal dan metode Tukey untuk penyesuaian nilai- p .
Regresi linear digunakan untuk menguji hubungan signifikan antara SWE tahun sebelumnya (November–Maret) dan curah hujan Juli tahun berjalan untuk semua tahun dari 1959 hingga 2020. Bulan Juli biasanya merupakan saat curah hujan monsun musim panas terjadi di wilayah ini. Untuk menentukan apakah mortalitas pada populasi alami dimediasi oleh variasi dalam A L :A S dan LAI:SA, regresi linear juga digunakan untuk menguji hubungan signifikan antara luas daun: luas gubal (A L :A S , LAI:SA), curah hujan Juli selama tahun kekeringan, dan % kematian tajuk yang dinilai pada tahun 2019 atau 2021 (tahun-tahun setelah setiap kekeringan). Nilai tingkat plot untuk A L :A S dirata-ratakan, menghasilkan satu titik data per hutan. Koefisien determinasi ( R 2 ) dan nilai- p ditentukan melalui regresi linear.
Analisis komputasional dan statistik dilakukan dalam R versi 4.2.1 (Tim Inti R, 2021 ). Paket ‘lme4’ (versi 1.1.30), ‘lmerTest’ (versi 3.1.3), dan ‘emmeans’ (versi 1.8.3) digunakan untuk membangun dan menganalisis model efek campuran linier (Bates et al., 2015 ; Kuznetsova et al., 2017 ; Lenth, 2022 ). Signifikansi efek tetap ditentukan dengan LRT menggunakan ‘lmerTest’ dan metode perkiraan Satterthwaite. Perbandingan berpasangan dilakukan menggunakan ‘emmeans’. Paket ‘car’ (versi 3.1.0) digunakan untuk memplot garis normal QQ dengan interval kepercayaan 95% untuk validasi asumsi model linier (Fox et al., 2018 ). Untuk semua analisis, tingkat signifikansi <0,05 dan =0,05–0,1 masing-masing dianggap signifikan secara statistik dan signifikan secara marginal.
3 HASIL
3.1 Variasi dalam A L :A S Melintasi gradien temporal dan spasial
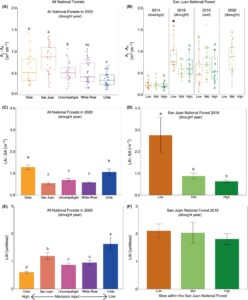
Untuk pola tingkat cabang, populasi aspen di hutan lintang selatan dan pada elevasi rendah di dalam NF San Juan umumnya mengalokasikan area daun yang jauh lebih banyak per unit area gubal (yaitu tingkat cabang yang lebih tinggi A L :A S ) selama tahun-tahun kekeringan parah 2018 dan 2020 (Gambar 2A,B ) dibandingkan dengan asumsi dasar (nilai 2014 untuk NF San Juan dan nilai NF Uinta untuk 2020; lihat metode untuk detail lebih lanjut tentang penentuan dasar).
Untuk pola tingkat tegakan, populasi aspen di Dixie NF (Gambar 2C ) dan pada elevasi rendah dengan San Juan NF (Gambar 2D ) juga mengalokasikan lebih banyak area daun per unit area gubal (yaitu LAI:SA tingkat tegakan yang lebih tinggi) selama tahun-tahun kekeringan parah 2018 dan 2020. Selain itu, populasi aspen di Dixie NF memiliki rasio LAI:BAI 8 yang lebih tinggi secara signifikan (Gambar S3 ), yang menunjukkan bahwa pertumbuhan gubal kemungkinan lebih lambat daripada produksi area daun dalam populasi ini. Nilai area gubal jauh lebih kecil untuk pohon-pohon dalam populasi Dixie NF (Gambar S1 ), yang tampaknya mendorong perbedaan untuk populasi ini antara pola rasio area daun tingkat cabang dan tegakan (misalnya Gambar 2A vs. C). Pola indeks area daun (LAI) tingkat tegakan menguatkan area daun yang lebih tinggi dalam San Juan NF selama kekeringan 2018 dan 2020 (Gambar 2E,F ). Sementara LAI tidak lebih tinggi secara signifikan di aspen dataran rendah di San Juan NF selama tahun 2018, perlu dicatat bahwa LAI aspen dataran rendah hampir setara dengan aspen dataran menengah dan tinggi yang ada di lingkungan yang lebih mesik (Gambar 2F ). Bertentangan dengan pola LAI:SA, LAI umumnya berkurang di lintang rendah, populasi aspen yang lebih monsunal, selain dari San Juan NF (Gambar 2E ). Namun, plot di Dixie NF memiliki banyak pohon aspen yang baru saja mati, yang menunjukkan bahwa LAI sebelum mati mungkin lebih dekat ke hutan yang lebih utara.
3.2 NAM dan pola produksi daun pada populasi pohon aspen yang bergantung pada musim hujan
Dengan menggunakan kumpulan data 62 tahun dari variabel iklim petak untuk setiap pita hutan/elevasi, kami mengukur hubungan antara ekuivalen air salju (SWE) dan curah hujan Juli. Kami menemukan bahwa tahun-tahun dengan lapisan salju rendah dikaitkan dengan curah hujan monsun (Juli) yang lebih kuat di beberapa lokasi tempat kami mendokumentasikan overshoot struktural. Kami mengamati hubungan negatif antara SWE dan peningkatan curah hujan musim panas di hampir semua populasi, dan hubungan ini secara statistik signifikan secara marginal untuk San Juan NF ( R 2 = 0,06, p = 0,052; Gambar 3a ). Sebaliknya, kami mengamati hubungan positif yang signifikan secara statistik antara SWE dan curah hujan musim panas di Uinta NF ( R 2 = 0,11, p = 0,01; Gambar 3a ), populasi lintang paling utara yang paling tidak bergantung pada curah hujan monsun. Di San Juan NF, ada hubungan negatif yang konsisten di mana tahun-tahun dengan SWE yang berkurang pada musim dingin sebelumnya memiliki curah hujan musim panas yang lebih tinggi (Gambar 3b ). Hubungan ini secara statistik sedikit signifikan untuk populasi aspen di dataran rendah ( R 2 = 0,05, p = 0,071) (Gambar 3b ).
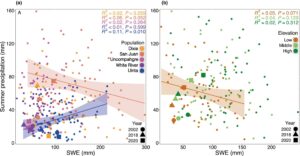
Di wilayah yang menjadi perhatian dalam studi ini, telah terjadi beberapa kekeringan parah dalam beberapa dekade terakhir, termasuk selama tahun 2002, 2018, dan 2020 (Williams et al., 2020 ). Secara umum, NAM gagal terwujud dan membawa curah hujan yang cukup ke hutan selama tahun-tahun kekeringan ini (selain tahun 2018 di Dixie NF dan tahun 2020 di San Juan NF; Gambar 3 , Tabel S1 ), yang mungkin berkontribusi terhadap kematian pohon aspen akibat kekeringan di wilayah investigasi kami.
3.3 Kelebihan luas daun maladaptif dan risiko kematian akibat kekeringan
Selama tahun 2020, pohon-pohon dari populasi lintang lebih selatan (Dixie, San Juan dan Uncompahgre) mengalami persentase kehilangan konduktansi hidrolik cabang yang lebih tinggi, yang menunjukkan potensi stres kekeringan yang meningkat, meskipun hasil ini tidak signifikan secara statistik (Gambar S4 ). Overshoot struktural sebagian dikaitkan dengan peningkatan risiko kematian setelah kekeringan parah di seluruh populasi dan lokasi. Pada tingkat cabang, populasi dengan produksi daun lebih banyak atau produksi gubal lebih rendah (yaitu tingkat cabang yang lebih tinggi A L :A S ) selama tahun-tahun kekeringan 2018 dan 2020 memiliki lebih banyak kematian tajuk pada musim tanam berikutnya ( R 2 = 0,53, p = 0,042; Gambar 4a ), hubungan yang khususnya penting untuk populasi dari lintang selatan (Dixie, San Juan) dan ketinggian rendah di dalam San Juan NF. Demikian pula, populasi dengan LAI:SA tingkat tegakan yang lebih tinggi selama tahun-tahun kekeringan 2018 dan 2020 juga memiliki lebih banyak kematian tajuk pada musim tanam berikutnya, meskipun hubungan ini tidak signifikan secara statistik ( R 2 = 0,32, p = 0,243), dan tampaknya terutama didorong oleh zona elevasi rendah dari San Juan NF (Gambar 4b ). Baik LAI:SA selama tahun kekeringan, curah hujan Juli selama tahun kekeringan (PPT), maupun istilah interaksi mereka tidak memiliki efek signifikan pada kematian tajuk tahun setelah kekeringan (Gambar 4b ).

4 DISKUSI
Dalam studi ini, kami mengukur dinamika luas daun dan hubungannya dengan kematian yang disebabkan kekeringan di seluruh populasi alami aspen di Hutan Nasional (NF) yang membentang di zona transisi utara Monsun Amerika Utara (NAM). Kami menemukan bahwa variasi dalam A L :A S berkorelasi dengan matinya tajuk pohon setelah kekeringan dan menyoroti peran potensial decoupling NAM sebagai mekanisme yang mendasari produksi daun aspen di wilayah ini. Hasil ini mendukung hipotesis bahwa terlalu banyak biomassa dialokasikan ke luas daun pohon per unit area gubal, yang mengakibatkan overshoot struktural yang tidak didukung oleh kelembaban tanah yang cukup selama periode kekeringan yang parah. Peningkatan yang diamati dalam matinya tajuk setelah tahun-tahun kekeringan yang parah menunjukkan bahwa plastisitas sifat maladaptif ini menyebabkan peningkatan stres hidrolik dan kematian aspen.
4.1 Variasi dalam A L :A S Melintasi gradien temporal dan spasial
Secara umum, rasio luas daun terhadap luas gubal diamati lebih tinggi di antara populasi aspen di daerah yang lebih hangat dan kering yang lebih bergantung pada presipitasi musim panas NAM, dan kami menemukan bukti adanya kelebihan struktural baik pada tingkat cabang maupun tegakan. Mengingat bahwa luas gubal di aspen ditentukan oleh pertumbuhan selama beberapa tahun (8 tahun atau lebih dalam ukuran cabang yang kami ukur yang mungkin lebih banyak daripada di batang pohon), kelebihan ini kemungkinan didorong oleh produksi daun yang berlebihan mengingat arsitektur hidrolik yang ada. Hal ini dapat diperburuk lebih lanjut oleh pertumbuhan radial yang dibatasi setelah luas daun terbentuk. Namun, hasil antara tingkat cabang A L :A S dan tingkat tegakan LAI:SA tidak identik, yang menunjukkan ada pemutusan hubungan antara pola alometri daun tingkat cabang dan tegakan yang masih harus diselesaikan. A L :A S biasanya menurun dengan tinggi pohon (McDowell et al., 2002 ), dan dua populasi dengan rasio LAI:SA tertinggi (Dixie dan Hutan Nasional San Juan dengan elevasi rendah) secara signifikan lebih pendek daripada populasi lainnya. Selain itu, kemungkinan ada variasi dalam alometri A L :A S dengan ukuran pohon di seluruh populasi ini, yang bergabung dengan variasi dalam distribusi ukuran tegakan untuk mempersulit penskalaan cabang-ke-tegakan. Akibatnya, area gubal jauh lebih kecil untuk pohon-pohon di Dixie NF dibandingkan dengan populasi lainnya, yang mengakibatkan perbedaan besar antara rasio cabang- dan tingkat tegakan dari produksi area daun. Selain itu, struktur tajuk (misalnya orientasi cabang, jumlah cabang, tinggi tajuk, dll.) di tegakan aspen ini mungkin berbeda karena perbedaan geografi lokasi (Zhang et al., 2016 ) atau karena variabilitas musiman (Wirth et al., 2001 ), yang mewakili bentuk plastisitas lain yang mungkin memediasi mortalitas hutan. Yang penting, data kami hanya mencerminkan rasio luas daun terhadap luas gubal pada satu titik waktu, dan dinamika yang melibatkan variasi musiman dalam rasio ini dapat mengubah temuan yang dilaporkan di sini.
Khususnya, hasil LAI dan LAI:SA menunjukkan bahwa perubahan tingkat tegakan seperti matinya pohon dalam skala besar menangkal alokasi berlebih A L :A S skala jaringan dari masing-masing pohon. A L :A S tingkat cabang memberikan metrik yang solid untuk ‘aklimatisasi fisiologis’ (yaitu aklimatisasi pada tingkat jaringan) tetapi bukan metrik yang kuat untuk ‘aklimatisasi ekosistem’ (yaitu aklimatisasi pada tingkat tegakan). Metrik terakhir ini penting untuk memahami respons fungsi ekosistem tingkat tegakan dan untuk memodelkan respons hutan aspen terhadap perubahan iklim di masa mendatang (Sperry et al., 2019 ), yang menjadikan pemutusan hubungan antara alometri tingkat cabang dan tegakan sebagai masalah kritis untuk dipecahkan. Hasil ini mungkin juga menunjukkan mekanisme kompensasi tingkat tegakan, di mana pohon aspen dengan lebih banyak mati tajuk memiliki lebih sedikit cabang tetapi luas daun per cabang yang lebih tinggi untuk mengimbanginya, yang mungkin sangat lazim di daerah rawan kekeringan yang tidak punya waktu untuk pulih di antara peristiwa kekeringan berikutnya (Wu et al., 2023 ). Di sini, kami mengamati peningkatan persentase kehilangan konduktansi hidrolik pada pohon selama kekeringan 2020, terutama di antara populasi yang lebih bergantung pada curah hujan musim panas NAM (Gambar S4 ), tetapi ini tidak dapat sepenuhnya memastikan bahwa mati pucuk kanopi terjadi akibat kegagalan hidrolik karena meningkatnya permintaan penguapan dari produksi daun yang lebih banyak. Estimasi skala transportasi air dari tingkat jaringan ke tingkat seluruh pohon atau petak juga tetap menjadi tantangan untuk pemodelan respons hutan aspen terhadap perubahan iklim (Meinzer et al., 2005 ; Mencuccini, 2002 ).
4.2 NAM dan pola produksi daun pada populasi pohon aspen yang bergantung pada musim hujan
Tingkat cabang A L :A S yang lebih tinggi dan tingkat tegakan LAI:SA selama tahun-tahun kekeringan parah yang diamati dalam studi ini, dan pada ketinggian yang lebih rendah dan lebih kering, dapat menempatkan pohon aspen pada peningkatan risiko stres fisiologis dan kerusakan, mengingat meningkatnya permintaan penguapan dan karenanya tampaknya menunjukkan kelebihan struktural (Jump et al., 2017 ). Produksi luas daun mungkin telah dipengaruhi oleh sinyal lingkungan lain sebelum perkembangan kekeringan musim tanam, seperti kondisi iklim musim gugur selama pembentukan tunas vegetatif pada musim tanam sebelumnya atau suhu musim semi yang lebih hangat yang mengakibatkan fenologi lanjutan selama 2018 dan 2020 (Gordo & Sanz, 2010 ; Uemura et al., 2000 ). Mengingat tahun-tahun dengan lapisan salju rendah secara historis berkorelasi dengan curah hujan monsun yang lebih kuat melalui umpan balik potensial daratan-atmosfer (Notaro & Zarrin, 2011 ; Zhu et al., 2005 ), peningkatan luas daun pada populasi aspen di lintang selatan mungkin juga merupakan hasil dari beberapa kombinasi perbedaan genetik dan/atau fenologi tingkat lanjut sebagai respons terhadap pencairan salju lebih awal dan suhu musim semi yang lebih hangat sebagai antisipasi curah hujan monsun musim panas yang lebih lebat.
Koefisien variasi rendah secara keseluruhan dalam analisis iklim kami (Gambar 3 ) mengindikasikan bahwa variabilitas monsun dipengaruhi oleh banyak faktor selain dari lapisan salju musim dingin sebelumnya. Namun, hubungan negatif yang terlihat di sini antara lapisan salju dan presipitasi musim panas di hutan yang bergantung pada monsun konsisten dengan mekanisme umpan balik daratan-atmosfer yang terdokumentasi dalam kumpulan data observasional dan model iklim (Gutzler & Preston, 1997 ; Notaro & Zarrin, 2011 ; Zhu et al., 2005 ). Namun, lapisan salju yang rendah hanyalah satu mekanisme yang mungkin memengaruhi NAM, dan investigasi lebih lanjut ke dalam pendorong iklim lainnya akan menjadi penting untuk lebih memahami respons hutan aspen terhadap perubahan iklim masa depan dalam ekosistem yang bergantung pada NAM. Memprediksi kapan ketidaksesuaian mungkin terjadi antara produksi daun dan decoupling NAM, seperti yang terlihat selama tahun-tahun kekeringan parah dalam studi ini, dapat memberikan wawasan yang lebih baik tentang tegakan hutan mana yang paling berisiko terhadap kematian akibat kekeringan.
Penting untuk dicatat bahwa penyerapan air oleh aspen mungkin lebih rumit karena ini adalah spesies klonal di mana pohon-pohon individual (ramet) terhubung ke ramet tetangga melalui sistem akar bersama (Barnes, 1966 ; Mitton & Grant, 2006 ). Pekerjaan sebelumnya menunjukkan bahwa sumber daya kemungkinan dibagi antara ramet melalui sistem akar (Baret & DesRochers, 2011 ; De Byle, 1964 ), namun studi yang difokuskan pada keterbatasan air dan pola kematian pada aspen dan balsam poplar ( Populus balsamifera ) menunjukkan ramet dapat bertindak lebih seperti pohon individu (yaitu sedikit bukti fasilitasi dan pembagian sumber daya) di bawah kekeringan (Adonsou et al., 2016 ; Anderegg et al., 2012 ). Teori model pipa yang menyatakan bahwa kapasitas pengangkutan air dalam batang menentukan luas daun tajuk (Shinozaki et al., 1964 ), mungkin tidak berlaku hanya untuk spesies klonal seperti aspen meskipun penelitian telah menunjukkan bahwa ramet aspen yang terjadi secara alami di Sierra Nevada, California sesuai dengan teori ini (Caldwell & O’Hara, 2017 ). Masih belum diketahui seberapa besar dinamika perakaran klonal memengaruhi pola alokasi biomassa dalam keseluruhan klon atau di seluruh distribusi hutan aspen.
4.3 Kelebihan luas daun dan risiko kematian akibat kekeringan
Sementara kematian pohon dapat terjadi karena beberapa faktor abiotik dan biotik, kematian aspen setelah kekeringan telah sangat terkait dengan kegagalan hidrolik dan hilangnya jaringan penghantar air dalam xilem pohon selama beberapa tahun pasca-kekeringan (Anderegg et al., 2012 ), terutama didorong oleh pengeringan tanah dangkal selama pertengahan/akhir musim tanam (Anderegg et al., 2013 ; Bell et al., 2014 ; Refsland & Cushman, 2021 ). Oleh karena itu, kurangnya presipitasi NAM selama tahun-tahun kekeringan kritis dalam penelitian kami (2018 dan 2020), dikombinasikan dengan suhu musim semi dan musim panas yang tinggi, mungkin telah mengakibatkan kurangnya pasokan air untuk memenuhi permintaan penguapan akan lebih banyak area daun dan kemungkinan penting dalam mendorong pola spasial kematian aspen seperti yang terlihat dalam hubungan positif yang signifikan antara tingkat cabang A L :A S dan kematian kanopi. Sementara hubungan LAI:SA tingkat tegakan dengan mortalitas tajuk menunjukkan tren yang sama dengan A L :A S tingkat cabang , kami berhipotesis bahwa hubungan yang tidak signifikan dengan LAI:SA mungkin terkait dengan variasi dan ketidakpastian yang lebih tinggi dalam penskalaan hubungan dari cabang ke tegakan. Skenario perubahan iklim saat ini memprediksi kejadian kekeringan yang lebih intens dan sering terjadi di wilayah ini, yang dapat menjadi sangat mematikan jika hubungan musiman antara lapisan salju dan curah hujan musim panas menjadi lebih terpisah (Pascale et al., 2017 ; Seager et al., 2007 ).
Hasil mati tajuk yang dilaporkan di sini mungkin tidak menunjukkan kematian pohon secara menyeluruh, namun aspen di wilayah ini telah terbukti menunjukkan kematian yang tertunda dan kematian yang disebabkan oleh kekeringan dari overshoot struktural selama kekeringan 2018 dan 2020 kemungkinan akan terus berlangsung selama dekade berikutnya. Memang, studi dari kekeringan sebelumnya telah menemukan bahwa ramet aspen yang mulai mengalami mati tajuk yang substansial cenderung berlanjut ke kematian penuh dalam 6–10 tahun (Anderegg et al., 2019 ; Worrall et al., 2010 ). Overshoot struktural mungkin merupakan mekanisme penting untuk menjelaskan bahwa kematian aspen sering kali tertunda beberapa tahun setelah kekeringan awal (Anderegg et al., 2013 ; Trugman et al., 2018 ).
Yang penting, kami tidak dapat menguji dampak agen mortalitas biotik pada mati pucuk tajuk dalam studi ini. Sementara penyebab utama mortalitas mungkin patogenik atau biotik, stres kekeringan dapat menurunkan kapasitas pohon aspen untuk melawan hama dan patogen, yang dapat menjadi kontributor signifikan terhadap mortalitas, terutama di wilayah rawan kekeringan di Amerika Serikat bagian barat daya (Marchetti et al., 2011 ). Pohon aspen dalam studi ini, terutama di wilayah yang lebih bergantung pada presipitasi NAM, memang mengalami peningkatan kehilangan konduktansi hidrolik selama kekeringan 2020 (Gambar S4 ), yang menunjukkan beberapa stres terkait kekeringan.
Hasil kami menunjukkan bahwa variasi dalam A L :A S dikaitkan dengan respons kekeringan pohon aspen, yang menyoroti pentingnya memahami mekanisme yang mendasari fenologi daun dan produksi daun serta implikasinya dalam konteks penentuan alokasi biomassa dan peramalan iklim. Sinyal lingkungan yang terlibat dalam fenologi daun dan produksi daun sering kali rumit dan mungkin disebabkan oleh kombinasi iklim tahun berjalan dan tahun sebelumnya (Gordo & Sanz, 2010 ; Uemura et al., 2000 ). Penyertaan parameter iklim, seperti ekuivalen air salju atau kelembapan tanah musim semi, sebagai tambahan parameter tradisional yang digunakan dalam sebagian besar model fenologi (yaitu hari derajat tumbuh, kebutuhan dingin, dll.) dapat memungkinkan prediksi model yang lebih baik (Richardson et al., 2013 ). Menentukan sinyal iklim langsung yang terlibat dalam produksi luas daun dan waktunya juga dapat meningkatkan kemampuan kita untuk memodelkan biomassa dan alokasi sumber daya di hutan aspen dalam iklim yang berubah (Rosas et al., 2019). Di antara populasi pohon aspen kami, kami mengamati variasi intraspesifik yang substansial dalam A L :A S , yang terjadi pada skala spasial dan temporal. Menggabungkan variasi sifat intraspesifik ke dalam model berbasis sifat yang memperhitungkan sifat fungsional penting seperti A L :A S akan sangat penting untuk meningkatkan kemampuan kami dalam memperkirakan respons hutan terhadap stres kekeringan di masa mendatang (Martínez-Vilalta et al., 2002 ; Sabot et al., 2022 ; Xu et al., 2021 ).
Temuan yang dilaporkan di sini memiliki implikasi yang lebih luas untuk spesies pohon lain dan ekosistem hutan. Di antara tipe spesies lain, ada bukti substansial bahwa rasio luas daun terhadap luas gubal sesuai dengan teori evolusi, yang menyoroti dampak potensial sifat ini pada kebugaran. Misalnya, spesies menunjukkan penurunan A L :A S dengan meningkatnya kekeringan (yaitu produksi luas daun lebih sedikit dalam kondisi lebih hangat/kering; Zhu & Zhao, 2023 ), penurunan A L :A S dengan konduktansi hidrolik batang yang lebih rendah (yaitu luas daun lebih rendah untuk menyamai kemampuan konduktansi air; Macinnis-Ng et al., 2004 ), dan penurunan A L :A S dengan meningkatnya tinggi pohon (yaitu pemeliharaan efisiensi hidrolik spesifik daun dengan konduktansi hidrolik seluruh tanaman yang dibatasi; McDowell et al., 2002 ). Namun, ada juga penyimpangan dari temuan ini yang menunjukkan rasio luas daun terhadap luas gubal mungkin tidak terkait erat dengan iklim dan sinyal lingkungan dalam semua kasus. Zhu dan Zhao ( 2023 ) juga menemukan efek interaksi signifikan antara suhu dan presipitasi yang menghasilkan tren berlawanan dalam A L :A S , yang menunjukkan sinyal lingkungan yang kompleks pada rasio luas daun terhadap luas gubal (Mencuccini et al., 2019 ). McDowell et al. ( 2002 ) melaporkan peningkatan A L :A S dengan tinggi pohon untuk dua spesies ( Picea abies dan Abies balsamea ), yang keuntungannya tampaknya tidak diketahui. McBranch et al. ( 2018 ) tidak menemukan pergeseran dalam A L :A S di antara pohon Pinus edulis dan Juniperus monoperma setelah eksperimen pemanasan dan kekeringan selama 5 tahun, yang menunjukkan bahwa proses aklimatisasi yang melibatkan rasio luas daun terhadap luas gubal dapat memakan waktu lebih lama dari satu musim tanam yang penuh tekanan. Dengan demikian, menjelaskan dan mengurai lebih lanjut pendorong rasio luas daun terhadap luas gubal di masa mendatang akan memberikan wawasan berharga untuk memahami bagaimana sifat utama ini menginformasikan pemodelan dan proyeksi dinamika ekosistem hutan di masa mendatang.
5 KESIMPULAN
Studi kami menghubungkan pola kelebihan luas daun dengan peningkatan mortalitas pada spesies pohon yang tersebar luas. Hasil ini memberikan contoh kemungkinan plastisitas sifat maladaptif yang menyebabkan peningkatan stres dan mortalitas selama kekeringan parah. Mereka menyoroti bahwa hubungan sifat-lingkungan sering kali rumit dan mungkin karena respons sifat terhadap isyarat iklim yang kompleks atau berbeda dan bahwa respons alokasi tanaman selama stres merupakan kebutuhan penelitian utama (De Kauwe et al., 2014 ; Falster et al., 2015 ). Kapan dan di mana plastisitas sifat bersifat adaptif, yang mengarah pada peningkatan ketahanan iklim versus maladaptif, yang mengarah pada peningkatan mortalitas pohon, serta pertimbangan tentang bagaimana aklimatisasi tingkat tegakan melalui perubahan kepadatan memengaruhi mortalitas dalam skala waktu yang panjang, tetap menjadi pertanyaan utama untuk menerangi masa depan hutan di abad ke-21.