
Abstrak
Di Afrika Selatan, hutan bakau dan padang lamun sering kali hidup berdampingan dalam sistem estuari dan dalam kombinasi menjadi inang bagi keanekaragaman hayati yang kaya dan seringkali endemik. Ada kebutuhan mendesak untuk mengeksplorasi keanekaragaman komunitas dalam ekosistem vegetasi pesisir, mengingat efek degradasi dari perubahan iklim dan tekanan antropogenik. Metabarkode DNA lingkungan adalah alat biomonitoring yang menjanjikan dalam ekosistem pesisir Afrika Selatan, meskipun penelitian menyoroti perlunya mengoptimalkan dan menstandardisasi protokol pengambilan sampel. Penelitian ini bertujuan untuk berkontribusi dalam mengembangkan protokol pengambilan sampel dengan memahami penerapan metabarkode DNA lingkungan (eDNA) dalam habitat vegetasi pesisir dan dengan menyelidiki keanekaragaman menggunakan dua jenis sampel yang berbeda. Sampel DNA lingkungan akuatik dan sedimen dari tiga estuari di sepanjang pantai timur Afrika Selatan, dikombinasikan dengan metabarkode MiFish, mendeteksi 148 unit taksonomi operasional (OTU) yang mewakili 67 genera ikan dari 50 famili, meskipun hanya 16% yang terpecahkan menjadi spesies. Kami mengamati bahwa sampel eDNA akuatik mendeteksi 97% dari total keanekaragaman ikan, yang menunjukkan bahwa ini adalah alat yang efisien untuk mendeteksi keanekaragaman iktiofauna secara komprehensif. Kami tidak mendeteksi komunitas ikan yang berbeda antara hutan bakau dan lamun dan temuan kami menggarisbawahi pentingnya mempertimbangkan konektivitas hidrologis untuk desain pengambilan sampel. Secara keseluruhan, pekerjaan kami memperkuat pertimbangan utama untuk studi metabarkode eDNA di masa mendatang yang difokuskan pada fauna ikan di muara, dan oleh karena itu berkontribusi untuk mengoptimalkan protokol pengambilan sampel guna mendukung pemantauan hayati keanekaragaman hayati pesisir.
1. PENDAHULUAN
Muara adalah ekosistem yang beragam hayati, dinamis, dan produktif yang melakukan berbagai fungsi, termasuk penyediaan tempat berkembang biak, pembibitan, dan mencari makan yang penting bagi ichthyofauna. Hal ini difasilitasi oleh keberadaan ekosistem vegetasi pesisir heterogen (CVE) termasuk bakau dan lamun (Barbier, 2017 ; Beck et al., 2009 ; Whitfield, 2017 ). Di Afrika Selatan, bakau terbatas pada pantai timur, di mana mereka dapat hidup berdampingan dengan lamun di muara (Adams et al., 2016 ). Sebaliknya, Zostera capensis yang terancam punah , lamun paling melimpah di negara tersebut, dapat ditemukan di muara di sepanjang garis pantai (Adams & van der Colff, 2018 ; von der Heyden et al., 2024 ).
Tekanan antropogenik lokal dan perubahan global mendorong hilangnya habitat, degradasi, dan redistribusi CVE (Adams & Rajkaran, 2021 ; Peer et al., 2018 ; von der Heyden et al., 2024 ). Dampak tekanan ini diperburuk oleh distribusinya yang terbatas ke muara (Adams & Rajkaran, 2021 ; von der Heyden et al., 2024 ). Dengan demikian, populasi mangrove dan lamun terfragmentasi, dengan cakupan yang relatif kecil (masing-masing ~2087 dan 1755 ha; Raw et al., 2023 ) dibandingkan dengan populasi global seperti di Australia (~11.142 ha mangrove di seluruh Australia dan ~35.000 ha lamun di Kawasan Warisan Dunia Great Barrier Reef; Coles et al., 2015 ; Lymburner et al., 2020 ).
Meskipun cakupannya relatif kecil, hutan bakau dan padang lamun mendukung keanekaragaman iktiofauna yang kaya dengan penggunaan dan distribusi regionalnya di CVE yang terdokumentasi dengan baik (misalnya Beckley, 1983 ; Edworthy & Strydom, 2016 ; James & Harrison, 2020 , 2022 ; McGregor & Strydom, 2018 ; Wasserman et al., 2020 ; Whitfield et al., 1989 ). Muara Afrika Selatan mendukung lebih dari 155 spesies terkait muara yang tercatat (Whitfield & Cowley, 2010 ) di mana 50 di antaranya endemik Afrika Selatan dan termasuk spesies seperti Rhabdosargus holubi (Heemstra et al., 2022 ). Komunitas iktiofauna yang beragam berkontribusi pada perikanan regional, mendukung beberapa kegiatan penting secara ekonomi seperti penangkapan ikan rekreasi, subsisten, dan komersial (Lamberth & Turpie, 2003 ), dengan ~50% ikan yang berasosiasi dengan muara dieksploitasi atau digunakan (Lamberth & Turpie, 2003 ). Bersama dengan dampak tekanan terus-menerus yang terjadi di muara secara nasional (van Niekerk et al., 2019 ) yang mendorong penurunan populasi spesies (Mann et al., 2014 ) dan kepunahan (Everett et al., 2015 ), ada kebutuhan untuk pemantauan berkelanjutan terhadap kelompok fauna penting ini untuk pengelolaan dan perlindungan.
Namun, teknik pengambilan sampel standar kemungkinan meremehkan keanekaragaman hayati lokal sementara pemantauan keanekaragaman hayati laut yang berkelanjutan, cepat dan efektif di Afrika Selatan tetap menjadi tantangan. Hal ini sebagian besar disebabkan oleh terbatasnya ketersediaan data, kurangnya sumber daya keuangan dan infrastruktur yang buruk yang menghambat pengumpulan dan penilaian data pada skala temporal dan spasial yang relevan (Griffiths et al., 2010 ; Smit et al., 2022 ). Metode yang ada seperti pengambilan sampel tangkapan dan survei visual telah berperan penting untuk inventarisasi keanekaragaman hayati, tetapi sering dianggap kurang efisien, lebih memakan waktu, padat karya dan berpotensi merusak (Stat et al., 2017 ; Valentini et al., 2016 ). Estimasi keanekaragaman hayati yang diperoleh dari survei visual sering dibatasi oleh kesulitan dalam membedakan spesies dengan morfologi riwayat hidup yang bervariasi (Lira et al., 2023 ). Lebih jauh lagi, kualitas data dari survei video bawah air jarak jauh yang menggunakan umpan dapat terpengaruh secara negatif oleh air yang keruh, yang mengurangi visibilitas dan sangat bergantung pada keahlian taksonomi pengamat (Cole et al., 2022 ; Gladstone et al., 2012 ; Wilms et al., 2022 ).
DNA metabarkode lingkungan memungkinkan pemantauan keanekaragaman hayati yang terstandarisasi dan berkelanjutan dalam ekosistem pesisir heterogen yang mencakup wilayah geografis yang luas (Miya, 2022 ; Taberlet et al., 2012 ; von der Heyden, 2023 ). Metode berbasis genetik ini menghasilkan potret kontemporer tingkat komunitas dengan resolusi spasial tinggi, yang diperoleh dengan kecepatan yang cepat dan terjangkau (Courtaillac et al., 2024 ; Jeunen et al., 2019 , 2020 ; Port et al., 2016 ; West et al., 2020 ). Materi genetik, disebut DNA lingkungan (eDNA), dapat berupa campuran DNA organisme dan ekstra-organisme, seperti hewan uniseluler, gamet, dan jaringan, yang tersuspensi dalam substrat lingkungan seperti air atau sedimen (Pawlowski et al., 2020 ; Rodriguez-Ezpeleta et al., 2021 ; Ruppert et al., 2019 ).
Sejumlah penelitian telah menggunakan eDNA metabarcoding untuk menyelesaikan dan meningkatkan inventaris keanekaragaman hayati di lingkungan estuari secara global, mengingat bahwa hal itu non-invasif, cepat dan hemat biaya untuk survei keanekaragaman hayati ikan (Alenzi, 2024 ; Miya, 2022 ; Thomsen & Willerslev, 2015 ). Di Malaysia, Zainal Abidin et al. ( 2022 ) menemukan bahwa pengambilan sampel eDNA mendeteksi ~82% dari keluarga ikan yang tercatat selama satu dekade melalui survei tradisional, semuanya hanya dalam waktu 2 hari. Analisis antara hulu, tengah dan hilir muara juga telah mengungkapkan komunitas terstruktur, meskipun konektivitas hidrologis (Garcia-Vazquez et al., 2021 ). Sebuah penelitian oleh Cole et al. ( 2022 ) di Australia menunjukkan bahwa eDNA metabarcoding menyelesaikan perbedaan dalam kekayaan dan komposisi ikan pada skala spasial kecil (<1 km).
Meskipun eDNA metabarcoding berpotensi digunakan sebagai alat untuk memantau ikan pesisir (Miya, 2022 ), masih terdapat beberapa tantangan dalam alur kerja (misalnya Alberdi et al., 2018 ; Shirazi et al., 2021 ; Wee et al., 2023 ), yang mencakup kebutuhan untuk menstandardisasi dan mengoptimalkan protokol pengambilan sampel eDNA (von der Heyden, 2023 ), dan pilihan jenis sampel terbaik, yaitu air atau sedimen, untuk memaksimalkan deteksi keanekaragaman hayati (Ruppert et al., 2019 ).
Meskipun sebagian besar studi eDNA iktiofauna telah menggunakan sampel air (Miya, 2022 ), eDNA sedimen (eDNA yang diperoleh dari sedimen yang terendam) memainkan peran yang semakin penting dalam studi perikanan karena memiliki sifat yang unik dibandingkan dengan eDNA akuatik (eDNA yang diperoleh dari air) (Huston et al., 2023 ), seperti tingkat persistensi yang lebih lama dan konsentrasi eDNA yang lebih tinggi (Sakata et al., 2020 ; Torti et al., 2015 ; Turner et al., 2015 ). Namun, hasil yang beragam dari ekosistem air tawar menunjukkan peningkatan deteksi eDNA ikan dalam sedimen (Sakata et al., 2020 ; Turner et al., 2015 ), sementara beberapa menunjukkan bahwa sampel air meningkatkan deteksi keanekaragaman hayati (Baldigo et al., 2017 ; Shaw et al., 2016 ). Yang terakhir ini konsisten dengan studi yang dilakukan di ekosistem pesisir, yang juga melaporkan bahwa sampel air mendeteksi lebih banyak keanekaragaman ikan secara signifikan (Tagliabue et al., 2023 ; Zhu et al., 2023 ). Sebaliknya, studi yang menargetkan komunitas metazoa yang lebih luas menganjurkan penggunaan gabungan jenis sampel untuk memaksimalkan deteksi spesies (misalnya Antich et al., 2021 ; Holman et al., 2019 ; Koziol et al., 2019 ). Meskipun demikian, ada penelitian terbatas yang membandingkan keanekaragaman ikan yang dinilai menggunakan eDNA akuatik dan eDNA sedimen yang dikumpulkan dari lingkungan muara, khususnya dalam konteks Afrika (von der Heyden, 2023 ).
Dengan latar belakang ini, penelitian ini bertujuan untuk berkontribusi pada pengembangan protokol pengambilan sampel dengan memahami penerapan eDNA metabarcoding dalam habitat vegetasi pesisir dan dengan menyelidiki keanekaragaman menggunakan dua jenis sampel yang berbeda. Tujuannya adalah untuk menyelidiki apakah eDNA metabarcoding mendeteksi perubahan dalam keanekaragaman ikan (kekayaan dan komposisi komunitas) antara (1) jenis sampel dan (2) habitat muara yang berbeda. Kami berhipotesis bahwa akan ada kekayaan yang lebih tinggi yang terdeteksi dari eDNA sedimen dibandingkan dengan eDNA akuatik. Kami selanjutnya berhipotesis bahwa tidak akan ada perbedaan dalam kekayaan atau komposisi komunitas antara bakau dan lamun yang terdeteksi oleh eDNA metabarcoding.
2 METODE DAN BAHAN
2.1 Wilayah Studi
Sampel dikumpulkan dari tiga muara: Nahoon, Mngazana dan Teluk Kosi (Gambar 1 ). Muara-muara ini terletak di daerah biogeografis subtropis dan tropis, beriklim hangat (van Niekerk et al., 2020 ; Gambar 1a ). Teluk Kosi menerima curah hujan musim panas dari September hingga April sementara Nahoon dan Mngazana menerima curah hujan bimodal yang mencapai puncaknya pada musim gugur (Maret) dan musim semi (Oktober/November) (Harrison, 2004 ). Muara-muara tersebut bervariasi dalam ukuran, dengan Teluk Kosi menjadi yang terbesar (5000 ha), diikuti oleh Mngazana (768 ha) dan Nahoon (33 ha), tempat mangrove ditanam pada tahun 1969 (Hoppe-Speer et al., 2015 ). Muara Nahoon dan Mngazana diklasifikasikan sebagai muara yang sebagian besar terbuka, sedangkan sistem Teluk Kosi merupakan danau muara, yang terdiri dari serangkaian empat danau yang terhubung, berakhir di cekungan pasang surut dangkal dengan flat pasang surut yang luas (van Niekerk et al., 2020 ). Di Teluk Kosi, konektivitas laut bervariasi, di mana sistem memiliki saluran masuk yang menyempit, memfasilitasi rentang pasang surut dari 0,1 hingga 0,15 m. Sebaliknya, Nahoon dan Mngazana terbuka antara 90% dan 100% dari waktu ke waktu ke laut, dengan rentang pasang surut rata-rata 0,75–1,5 m. Nahoon dan Mngazana secara khas linier dalam geomorfologi, di mana aksi pasang surut dalam kombinasi dengan masukan fluvial mendorong gradien salinitas yang jelas. Pencampuran hidrologi di Teluk Kosi difasilitasi oleh angin dan masukan fluvial sesekali (van Niekerk et al., 2020 ).
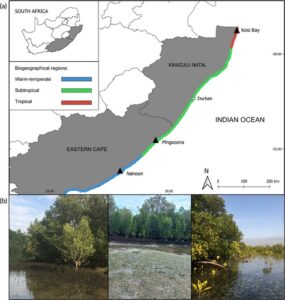
Mangrove seperti Avicennia marina, Rhizophora mucronata dan Bruguiera gymnorhiza tumbuh di ketiga muara, meskipun A. marina mendominasi pinggiran pasang surut (Adams et al., 2016 ) . Zostera capensis juga menonjol sebagai padang rumput berkelanjutan di zona pasang surut dan subpasang surut Nahoon dan Mngazana, sementara padang rumput yang tidak merata dari Halodule uninervis dan Z. capensis tersebar di seluruh dataran pasang surut Teluk Kosi (Adams et al., 2016 ; von der Heyden et al., 2024 ). Di ketiga sistem, dua tipe ekosistem vegetasi terhubung dengan baik secara hidrologis selama pasang surut karena mereka tumbuh sebagai konfigurasi mosaik di sepanjang tepian pasang surut muara (Gambar 1b ).
Muara Nahoon dilindungi dan terletak di kawasan perkotaan sehingga rentan terhadap tekanan antropogenik seperti polusi (Hoppe-Speer et al., 2015 ), sementara muara Mngazana berada di komunitas pesisir yang relatif kurang berkembang, dengan beberapa rumah yang dekat dengan muara, dan Teluk Kosi terletak di kawasan lindung (Peer et al., 2022 ).
2.2 Pengambilan sampel DNA lingkungan
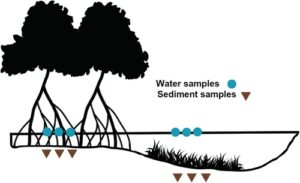
Di dalam masing-masing dari tiga muara, tiga lokasi diambil subsampelnya dengan dua titik pengambilan sampel per lokasi yang mencakup kontinum mangrove dan lamun. Sampel air dan sedimen dikumpulkan dalam rangkap tiga di mangrove dan lamun di zona pasang surut dari setiap sublokasi. Jarak antara lokasi pengambilan sampel mangrove dan lamun adalah ~10 m dan jarak antara setiap sublokasi adalah 500 m. Secara total, kami mengumpulkan 108 sampel yang terdiri dari 54 sampel air dan 54 sampel sedimen dari tiga muara ( n = 18 per muara) (Gambar 1 ) pada bulan Februari 2023. Jenis sedimen bervariasi antara muara, dengan sedimen lanau-lempung di Nahoon dan Mngazana dan sedimen berpasir di Teluk Kosi.
Sampel sedimen dikumpulkan dalam rangkap tiga saat air surut ketika sedimen terekspos (Gambar 2 ). Tabung Falcon digunakan untuk mengumpulkan 20 mL sedimen dari permukaan sedimen, setelah itu sampel diberi label, disegel dengan parafilm dan disimpan di atas es sampai dikembalikan ke laboratorium. Sampel sedimen dibekukan pada suhu -25 °C sampai ekstraksi DNA selesai. Setelah pengambilan sampel sedimen, lokasi pengambilan sampel yang sama diakses melalui perahu pada titik tertinggi air pasang untuk mengumpulkan sampel air. Sampel air dikumpulkan dalam rangkap tiga (Gambar 2 ) menggunakan spuit 50 mL, kemudian disaring terus menerus melalui unit Sterivex 0,22 μm sampai tidak ada air yang dapat melewatinya, sambil mencatat volume yang disaring untuk setiap sampel (Tabel S1 ). DNA yang ditangkap diawetkan dengan menyuntikkan 2 mL buffer ATL (Qiagen) ke dalam Sterivex, diikuti dengan penyegelan dengan Helapet Combi-caps (Helapet, Inggris) dan parafilm. Semua sampel disimpan pada suhu ruangan hingga ekstraksi. Untuk memperhitungkan kemungkinan kontaminasi di lapangan, kontrol negatif (satu kontrol negatif per lokasi, total tiga) disertakan menggunakan protokol yang sama, dengan menyaring 500 mL air suling. Di lapangan, sarung tangan diganti setiap kali pengambilan sampel dan peralatan disterilkan dengan larutan natrium hipoklorit 10%.
2.3 Protokol ekstraksi DNA
Sekitar 1 mL sedimen dari setiap sampel digunakan untuk ekstraksi eDNA, menggunakan kit Qiagen DNeasy PowerSoil, mengikuti spesifikasi pabrik. Tiga ekstraksi dilakukan dari setiap sampel sedimen. DNA yang diekstraksi disimpan pada suhu −25°C. eDNA akuatik diekstraksi menggunakan kit DNeasy Blood and Tissue (Qiagen), mengikuti protokol modifikasi dari Czachur et al. ( 2022 ). DNA diekstraksi langsung dari filter Sterivex, dengan protokol ini diterapkan pada semua kontrol negatif. Untuk mencegah kontaminasi, semua ekstraksi dilakukan di laboratorium yang hanya diperuntukkan bagi ekstraksi eDNA di Departemen Botani dan Zoologi, Universitas Stellenbosch; permukaan dan peralatan diekspos ke UV selama 30 menit sebelum ekstraksi dan dibersihkan dengan larutan pemutih 10%. Kontrol negatif ( n = 4) disertakan dalam setiap batch ekstraksi untuk memantau potensi kontaminasi. DNA yang diekstraksi disimpan pada suhu −20°C.
2.4 Persiapan dan pengurutan perpustakaan
Tiga replikasi DNA dari setiap sampel sedimen digabungkan sebelum pengurutan, menghasilkan total 54 sampel sedimen dan 54 sampel akuatik yang digunakan untuk konstruksi pustaka. Untuk penelitian ini, kami menggunakan primer MiFish E/U, yang mengamplifikasi sebagian dari mtDNA 12S rRNA dan telah berhasil digunakan dalam sejumlah penelitian yang berfokus pada komunitas iktiofauna (Miya, 2022 ), termasuk di Afrika Selatan (Courtaillac et al., 2024 ; Czachur et al., 2022 ; Rossouw et al., 2024 ). Persiapan dan pengurutan pustaka dilakukan oleh Clockmics (Jepang), menggunakan metode Ushio et al. ( 2022 ).
PCR pertama dilakukan pada empat replikasi teknis dengan volume reaksi 20-μL yang berisi 10,0 μL 2× Platinum SuperFi II PCR Master Mix, 6,0 μL setiap set primer PCR F/R 1,66 μM pertama, dan 4,0 μL cetakan DNA (lihat Tabel S2 untuk profil siklus termal). Set primer PCR pertama terdiri dari primer sekuensing Illumina, sekuens spesifik sampel 8-basa dengan kombinasi ganda indeks maju dan mundur (i5 dan i7), dan primer MiFish E/U.
Setelah amplifikasi, 1 μL ExoSAP-IT Express ditambahkan ke setiap produk PCR 20-μl. Campuran tersebut diinkubasi pada suhu 37°C selama 30 menit, diikuti oleh suhu 80°C selama 1 menit. Produk PCR pertama digabungkan menjadi satu sampel dan dimurnikan menggunakan manik-manik AMPure XP (produk PCR:manik-manik AMPure XP = 1:0,8). Produk yang dimurnikan berfungsi sebagai cetakan untuk PCR kedua.
Adaptor Illumina P5/P7 ditambahkan ke sampel PCR pertama yang dikumpulkan selama PCR kedua. PCR kedua dilakukan dengan volume reaksi 20 μL yang berisi 10,0 μL 2× Platinum SuperFi II PCR Master Mix, 8,0 μL setiap primer PCR F/R 1,25 μM (lihat Tabel S1 untuk profil siklus termal) dan 2,0 μL produk PCR pertama yang diencerkan.
Produk PCR kedua yang dikumpulkan dimurnikan menggunakan manik-manik AMPure XP (produk PCR:manik-manik AMPure XP = 1:0,8; Beckman Coulter). DNA berukuran target dari pustaka yang dimurnikan dan dikonsentrasikan dua kali lipat dipotong menggunakan E-Gel SizeSelect (Thermo Fisher Scientific). Konsentrasi DNA untai ganda dari pustaka diukur menggunakan Quantus Fluorometer (Promega).
Konsentrasi pustaka disesuaikan menjadi 50 pM menggunakan 10 mM Tris–HCl (pH 8,5). Pengurutan dilakukan pada platform Illumina NovaSeq dengan kit 500 siklus (2 × 150 bp paired-end sequencing; Illumina), yang menggabungkan 30% PhiX spike-in untuk meningkatkan kualitas pengurutan.
2.5 Pengolahan data sekuens dan penugasan taksonomi
Urutan mentah didemultipleks dan daerah primer dipangkas menggunakan cutadapt 2.10 (Martin, 2011 ). Urutan yang didemultipleks dan dipangkas primer diproses menggunakan DADA2 (Callahan et al., 2016 ), pendekatan varian urutan amplikon (ASV). Pertama, untuk proses penyaringan kualitas, bacaan berkualitas rendah dan pendek yang tidak terduga dihilangkan (≧Q30). Tingkat kesalahan dihitung dan urutan didereplikasi, kesalahan dikoreksi, dan digabungkan untuk menghasilkan matriks sampel ASV, diikuti dengan penghapusan urutan chimeric. ASV yang terdeteksi digabungkan dan dikelompokkan menjadi unit taksonomi operasional (OTU) pada kesamaan 97% menggunakan paket DECIPHER R (Wright, 2016 ), yang mengubah matriks sampel ASV menjadi matriks sampel OTU. Varian sekuens amplikon (ASV) diubah menjadi OTU karena ASV sering mendeteksi variabilitas intraspesifik, sedangkan fokus penelitian ini adalah komposisi komunitas ichthyofauna pada tingkat spesies dan tingkat yang lebih tinggi (misalnya Antich et al., 2023 ; Tsuji et al., 2020 ).
Identifikasi taksonomi dilakukan untuk OTU berdasarkan metode auto-k-nearest-neighbour (QCauto) yang berpusat pada kueri (Tanabe & Toju, 2013 ) yang diterapkan di Claident ( www.claident.org ). Metode QCauto adalah metode konservatif, karena semua tetangga terdekat dari urutan kueri harus memiliki informasi taksa yang sama untuk penugasan taksa. Misalnya, jika metode QCauto menugaskan nama genus ke OTU, semua tetangga terdekat di sekitar OTU memiliki nama genus yang sama. Metode ini meningkatkan jumlah OTU tanpa nama spesies tetapi mengurangi potensi penugasan yang salah dalam informasi spesies (Sato et al., 2018 ; Wang et al., 2007 ). Taksonomi ditetapkan ke OTU menggunakan penelusuran BLAST terhadap basis data GenBank (NCBI), dengan persentase batas identitas 97 (spesies), Karena metode QCauto memerlukan setidaknya dua sekuens dari satu takson, DNA standar diidentifikasi secara terpisah menggunakan BLAST (Camacho et al., 2009 ). Karena kelompok sasaran studi ini adalah ichthyofauna, kumpulan data MiFish yang dihasilkan kemudian difilter secara taksonomi untuk OTU yang ditetapkan ke Actinopterygii dan Chondrichthyes, dengan menghilangkan yang tunggal. Penetapan taksonomi OTU dan distribusi geografis taksa ditinjau menggunakan catatan taksonomi berikut: Ikan Pesisir Samudra Hindia Barat (Heemstra et al., 2022 ), basis data World Register of Marine Species (WoRMS), dan basis data FishBase.
2.6 Analisis statistik
Metadata sampel (yaitu nama sampel, muara, lokasi, jenis sampel eDNA), tabel takson, dan tabel OTU diimpor dan diproses menggunakan phyloseq (McMurdie & Holmes, 2013 ) dan vegan (Oksanen et al., 2007 ) di R studio (versi 4.3.2). Kurva rarefaksi jumlah OTU pada peningkatan jumlah pembacaan, serta kurva akumulasi OTU dengan peningkatan jumlah sampel dibuat. Plot batang dan diagram Venn dibuat menggunakan kumpulan data yang tidak dirarefikasi.
Rarefaction hingga 2108 pembacaan (jumlah minimum pembacaan per sampel) (McKnight et al., 2019 ; Weiss et al., 2017 ) dipilih sebagai metode normalisasi, menggunakan fungsi rarefy . Sampel air dan sedimen dari Mngazana dan Teluk Kosi, dengan pembacaan kurang dari 2108, dikecualikan dari analisis berikut. Semua analisis statistik berikut dilakukan pada tingkat OTU karena resolusi taksonomi hingga tingkat spesies.
Untuk menguji kemungkinan perbedaan dalam keragaman alfa (kekayaan OTU) antara tipe sampel eDNA, tipe habitat, dan di antara muara yang dijadikan sampel, analisis varians permutasi univariat tiga arah (PERMANOVA, α = 0,05; Anderson et al., 2008) dilakukan menggunakan matriks jarak Euklides.
Keanekaragaman beta dihitung menggunakan metrik Jaccard dalam vegan . Jumlah baca semua sampel diubah menjadi tabel ada-tidaknya OTU untuk menghitung matriks jarak Jaccard. Pola komposisi komunitas divisualisasikan dengan ordinasi multidimensi non-metrik (NMDS) menggunakan fungsi vegan metaMDS. PERMANOVA multivariat dilakukan dengan 999 permutasi bebas dengan fungsi adonis dalam vegan untuk menilai komunitas menggunakan faktor jenis sampel eDNA, jenis habitat, dan muara. Karena desain pengambilan sampel yang tidak seimbang (yaitu jumlah sampel yang tidak sama untuk faktor tetap: jenis sampel eDNA, jenis habitat, dan muara), yang dapat sangat membiaskan hasil analisis PERMANOVA (Anderson & Walsh, 2013 ), homogenitas kelompok menggunakan betadisper (dengan 1000 permutasi) diuji. Pengujian ini diimplementasikan menggunakan rutin PERMDISP2 berdasarkan centroid (Anderson, 2006 ). Analisis SIMPER dilakukan untuk mendapatkan keluarga yang bertanggung jawab atas perbedaan antara faktor-faktor yang menunjukkan perbedaan signifikan dalam PERMANOVA. Keluarga utama diidentifikasi sebagai keluarga yang menyumbang lebih dari 70% perbedaan antara kelompok.
3 HASIL
3.1 Urutan hasil
Metabarcoding dari uji MiFish ( n = 108 sampel DNA lingkungan) menghasilkan 15.822.752 pembacaan sekuensing. Setelah penyaringan kualitas, demultiplexing, dereplika dan eliminasi chimera, total 15.160.369 pembacaan dipertahankan, sementara tujuh sampel tidak berhasil selama amplifikasi PCR dan selanjutnya dikeluarkan dari analisis. Pembacaan ini bervariasi antara sampel sebagai berikut: 0 dan 923.684 pembacaan (147.087 ± 19.071 per sampel; rata-rata ± SE; Gambar S1 ). Dataset terdiri dari 214 OTU metazoa dalam sampel yang 162 OTU milik ichthyofauna, sedangkan dataset akhir terdiri dari 148 OTU setelah penjernihan. Teluk Kosi memiliki jumlah pembacaan tertinggi (8.367.055; 35 sampel), diikuti oleh Nahoon (4.529.556 pembacaan; 35 sampel) dan Mngazana (596.543 pembacaan; 31 sampel) (Gambar S1 ). Dari kurva rarefaction (Gambar S2 ), kedalaman sekuensing bervariasi di seluruh sampel, sementara kurva akumulasi OTU (Gambar S3 ) tidak jenuh, meskipun kurva hampir mencapai plateau, yang menunjukkan bahwa peningkatan upaya pengambilan sampel kemungkinan akan meningkatkan jumlah total OTU yang diambil.
3.2 Keanekaragaman taksonomi secara keseluruhan
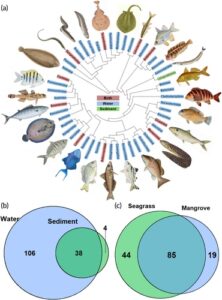
Secara total, metabarcoding sampel air dan sedimen yang dikumpulkan di habitat mangrove dan lamun mendeteksi 67 genera di 50 famili dan 33 ordo dalam Actinopterygii dan Chondrichthyes, yang diwakili oleh total 148 OTU (Tabel S3 ). Chondrichthyes diwakili oleh sembilan OTU dan terdiri dari lima famili berikut: Dasyatidae, Rajidae, Torpedinidae, Gymnuridae dan Myliobatidae. Secara keseluruhan, 85% dari 148 OTU dipecahkan ke ordo, 82% ke famili dan 59% ke tingkat genus dan hanya 16% dari OTU yang dapat ditetapkan ke spesies (Tabel 1 ).
| Hutan bakau | Rumput laut | ||
|---|---|---|---|
| Air | Memesan | 16 | 20 |
| Keluarga | 45 | 55 | |
| Marga | 35 | 41 | |
| Jenis | 16 | 20 | |
| Endapan | Memesan | 2 | 14 |
| Keluarga | 11 | 16 | |
| Marga | 10 | 17 | |
| Jenis | 8 | 4 | |
3.3 Pola keanekaragaman pada sampel eDNA dan tipe habitat yang berbeda
Meskipun kami mendeteksi tumpang tindih dalam OTU dari sampel akuatik dan sedimen, sampel air mengandung ~2,8 kali lebih banyak OTU daripada sedimen dan mendeteksi 97% dari semua OTU. Dari 144 OTU yang terdeteksi dalam sampel air, 38 juga terdapat dalam sedimen, sementara hanya empat OTU (OTU30 Leiognathidae spp.; OTU57 Dasyatis spp.; OTU59 Gymnothorax undulatus ; OTU171 Gobiidae spp.) yang hanya terdeteksi dari sedimen (Gambar 3 ). Sebaliknya, ~57% dari OTU yang terdeteksi dibagi antara habitat, sementara 44 OTU secara eksklusif terdeteksi di lamun dan 19 OTU di bakau (Gambar 3 ). Famili Actinopterygii yang secara konsisten terdeteksi dalam kelimpahan yang tinggi, tetapi bervariasi dalam sampel akuatik dan sedimen, termasuk Gobiidae, Mugilidae, Clupidae dan Sparidae (Gambar 4 ).

PERMANOVA univariat menunjukkan bahwa jenis sampel eDNA, muara, dan interaksi keduanya memiliki efek signifikan terhadap kekayaan OTU yang diamati (Tabel 2 ). Keragaman yang terdeteksi dalam sampel eDNA akuatik secara signifikan lebih tinggi daripada sampel sedimen (Gambar 3 ).
| Bahasa Indonesia : df | Jumlah kuadrat | R 2 | Pseudo- F | Nilai P | |
|---|---|---|---|---|---|
| Substrat | 1 | 4347.6 | 0.43497 | 114.7055 | 0,001** |
| Habitat | 1 | 0.2 | 0,00002 | 0,0044 tahun | 0,958 |
| Muara | 2 | 1315.6 | 0.13163 | 17.3555 | 0,001** |
| Jenis sampel × habitat | 1 | 0.6 | 0,00006 | 0,0171 tahun | 0.883 |
| Jenis sampel × muara | 2 | 1309.9 | 0.13106 | 17.2803 | 0,001** |
| Habitat × muara | 2 | 94.8 | 0,00949 | 1.2509 | 0.292 |
| Sisa | 76 | 2880.6 | 0.28820 | ||
| Total | 87 | 9995.1 | 1.00000 |
Catatan : ** p = 0,001.
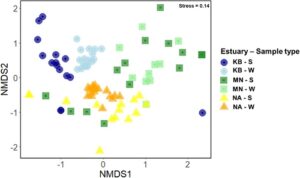
Komposisi komunitas ikan berdasarkan keberadaan dan ketiadaan OTU menunjukkan bahwa komunitas tersebut terstruktur di antara berbagai faktor yang dianalisis. Analisis PERMANOVA multivariat tiga arah mengungkapkan bahwa komposisi komunitas dipengaruhi secara signifikan oleh jenis sampel eDNA, muara, dan interaksinya (Tabel 3 ). Efek signifikan dari interaksi tersebut menunjukkan bahwa efek jenis sampel eDNA pada komunitas ikan bervariasi tergantung pada muara yang diambil sampelnya, dan sebaliknya. Muara memberikan kontribusi terbesar terhadap variasi yang diamati antara komunitas (12%) diikuti oleh interaksi antara jenis sampel eDNA dan muara (6%). Uji homogenitas multivariat dari dispersi kelompok signifikan untuk jenis sampel eDNA ( nilai F 1 = 6,5379, p = 0,01) dan perbandingan muara ( nilai F 1 = 5,5704, p = 0,005), yang menunjukkan bahwa dispersi data tidak homogen, sementara tidak ada perbedaan signifikan ( p > 0,05) dalam homogenitas dispersi kelompok untuk perbandingan jenis habitat. Hasil PERMANOVA didukung oleh ordinasi NMDS, yang menunjukkan bahwa komunitas dari berbagai jenis habitat tumpang tindih dengan beberapa derajat diferensiasi komunitas, meskipun komunitas mengelompok lebih jelas menurut jenis sampel eDNA (Gambar 5 ). Dari analisis SIMPER, famili yang berkontribusi terhadap perbedaan komunitas antara jenis sampel eDNA meliputi Mugilidae diikuti oleh Sparidae, Gobiidae, Eleotridae, Atherinidae dan Clupeidae. Famili yang sama berkontribusi terhadap perbedaan antara mangrove dan lamun, kecuali Ambassidae, bukan Atherinidae, melainkan famili yang penting. Mugilidae dan Gobiidae secara konsisten berkontribusi terhadap perbedaan antara muara.
| df | Jumlah kuadrat | R 2 | Pseudo- F | nilai p | |
|---|---|---|---|---|---|
| Jenis sampel | 1 | 1.605 | 0,04110 | 4.6056 | 0,001** |
| Habitat | 1 | 0,549 tahun | 0,01406 | 1.5753 | 0.153 |
| Muara | 2 | 4.642 | 0.11890 | 6.6623 | 0,001** |
| Jenis sampel × habitat | 1 | 0.513 | 0,01313 | 1.4712 | 0.251 |
| Jenis sampel × muara | 2 | 2.304 | 0,05901 tahun | 3.3067 | 0,001** |
| Habitat × muara | 2 | 0,999 | 0,02560 | 1.4343 | 0,077 tahun |
| Jenis sampel × habitat × muara | 2 | 0.910 | 0,02330 | 1.3054 | 0,029* |
| Sisa | 76 | 27.524 | 0.70492 | ||
| Total | 87 | 39.046 | 1.00000 |
Catatan : * p < 0,05; ** p = 0,001.
4 DISKUSI
Muara dan ekosistem vegetasi pesisir terkaitnya mendukung beragam komunitas, termasuk ikan, dan mewakili tempat berkembang biak, pembibitan, dan mencari makan yang penting bagi taksa yang penting secara komersial dan ekologis (Beck et al., 2009 ). Namun, sistem ini semakin terancam oleh perubahan global (James et al., 2013 ) dan tekanan antropogenik (Whitfield et al., 2018 ). Pemantauan berkelanjutan terhadap komunitas fauna terkait sangat penting untuk menginformasikan pengelolaan dan konservasi berbasis bukti, tetapi terbatas karena ketersediaan sumber daya logistik dan teknis yang diperlukan (Griffiths et al., 2010 ; Smit et al., 2022 ). Mengingat bahwa eDNA metabarcoding menghindari beberapa tantangan yang terkait dengan alat konvensional (Deiner et al., 2017 ; von der Heyden, 2023 ), ada ruang lingkup yang besar untuk menyelidiki iktiofauna muara menggunakan pendekatan molekuler ini. Namun, pengaruh jenis sampel eDNA pada deteksi komunitas iktiofauna lokal belum diteliti dengan baik dalam sistem muara, termasuk di Afrika Selatan.
4.1 eDNA akuatik saja sudah cukup untuk menangkap komunitas ichthyofauna
Sementara penelitian sebelumnya menunjukkan estimasi keanekaragaman yang kuat diperoleh dari pengambilan sampel kombinasi berbagai jenis sampel eDNA (misalnya Antich et al., 2021 ; Holman et al., 2019 ; Koziol et al., 2019 ) atau bahwa DNA ikan lebih terkonsentrasi di sedimen (Sakata et al., 2020 ; Turner et al., 2015 ), hasil kami kurang kongruen. Jumlah total spesies yang terdeteksi dari eDNA secara signifikan lebih tinggi dalam sampel air dibandingkan dengan sampel sedimen dan banyak OTU ikan hanya terdeteksi oleh eDNA akuatik, termasuk taksa yang melimpah di wilayah Samudra Hindia Barat seperti kakap merah bakau ( Lutjanus argentimaculatus ), belut moray bergelombang ( Gymnothorax undulatus ) dan ikan penggerutu bulan sabit ( Terapon jarbua ) serta spesies yang hampir terancam dengan distribusi terbatas ( Anguilla mossambica ; Pike et al., 2020 ).
Deteksi spesies laut pelagis lepas pantai (Houndfish Tylosurus crocodilus , bonito, dan sarden) melalui eDNA akuatik menggarisbawahi peningkatan kemampuan untuk mendeteksi kelompok taksonomi dengan distribusi spasial yang luas. Seperti yang dicatat dalam penelitian sebelumnya (Koziol et al., 2019 ; Tagliabue et al., 2023 ; Zhu et al., 2023 ), sifat bergerak ichthyofauna dan tahap riwayat hidup planktoniknya dikombinasikan dengan transportasi dan difusi eDNA di kolom air memungkinkan metabarcoding eDNA akuatik untuk menangkap gambaran yang lebih luas tentang keanekaragaman spesies dari wilayah geografis yang lebih luas.
Meskipun metabarkode eDNA tidak dapat membedakan antara tahap riwayat hidup (Ruppert et al., 2019 ), ia berfungsi sebagai metode yang ampuh untuk mendeteksi dan mengidentifikasi taksa yang mewakili komunitas planktonik (Leduc et al., 2019 ) atau kekurangan fitur morfologi utama karena morfologi riwayat hidup yang bervariasi (Lira et al., 2023 ). Misalnya, metabarkode mendeteksi beragam famili yang telah dideskripsikan sebagai yang umum terkait dengan habitat muara, termasuk mullet dan sparid (Whitfield, 2005 ). Spesies laut ini merekrut ke dalam muara dalam tahap perkembangan postfleksi atau juvenil awal, baik melalui migrasi aktif (Pattrick & Strydom, 2014 ) atau pasif (Harris et al., 1995 ). Sejalan dengan studi empiris regional (James et al., 2016 ; Whitfield & Mann, 2023 ), famili yang paling melimpah yang terdeteksi dalam studi ini adalah Mugilidae, yang diwakili oleh genera Chelon , Liza dan Mugil .
Larva sering terlewatkan oleh metode berbasis morfologi tradisional namun mewakili sebagian besar komunitas ikan di lingkungan muara (Beck et al., 2009 ; Strydom, 2015 ; Whitfield, 2005 ). Survei keanekaragaman ikan yang menargetkan tahap larva di muara Afrika Selatan umumnya terbatas (Strydom, 2015 ; Strydom et al., 2003 ; Strydom & Kisten, 2020 ), sebagian karena tantangan dalam memperoleh estimasi keanekaragaman dari proporsi komunitas ini. Misalnya, teknik penarikan plankton umumnya digunakan, yang memerlukan penyaringan air dalam volume besar untuk memusatkan komunitas target (khususnya larva) dan mengidentifikasi individu secara taksonomis secara visual di bawah mikroskop (Steinke et al., 2016 ). Bersama dengan penelitian di tempat lain (misalnya Garcia-Vazquez et al., 2021 ), penelitian kami menggarisbawahi bahwa metabarkode eDNA akuatik dapat memberikan inventaris keanekaragaman ikan yang berpotensi lebih komprehensif melampaui tahap dewasa dan remaja yang lebih mudah dikenali.
Di samping ikan belanak dan ikan sparid, studi ini juga mendeteksi taksa yang jumlahnya sedikit, yang sulit dideteksi dan diidentifikasi menggunakan metode survei tradisional seperti saat menggunakan jaring dan sensus visual (Deiner et al., 2017 ; Naputo et al., 2024 ) dan mencakup taksa yang tersamar dengan baik, sulit dipahami, dan tersembunyi (ikan pipa, ikan gobi, dan belut kepingan salju Echidna nebulosa ).
Dibandingkan dengan sampel eDNA akuatik, sedimen menangkap proporsi yang lebih kecil dari keragaman spesies yang ada. Masih banyak ketidakpastian mengenai berbagai faktor yang memengaruhi transportasi DNA lingkungan, pengendapan, dan pengawetan dalam sedimen laut (Huston et al., 2023 ), tetapi hasil kami menunjukkan pengawetan, pengendapan, dan sedimentasi eDNA iktiofauna yang buruk di lapisan permukaan sedimen di zona pasang surut muara, mungkin mencerminkan sifat abiotik dinamis zona litoral muara. Namun, efek dari proses abiotik ini mungkin berbeda, misalnya, di badan air tempat air memiliki waktu tinggal lebih lama atau kurang dinamis. Hal ini dapat memfasilitasi peningkatan laju transportasi, pengendapan, dan pengawetan untuk mendeteksi sinyal eDNA dalam sedimen (Nevers et al., 2020 ; Sakata et al., 2020 ; Turner et al., 2015 ).
Sifat fisik dan geokimia memainkan peran kunci dalam mengendalikan deteksi DNA dalam air (Kumar et al., 2022 ) dan sedimen (Buxton et al., 2018 ; Sakata et al., 2020 ). Faktanya, kami menemukan efek signifikan dari interaksi jenis muara-sampel untuk PERMANOVA univariat dan multivariat, yang menunjukkan bahwa perbedaan dalam komunitas iktiofauna yang diamati antara kedua jenis sampel eDNA bergantung pada muara. Perbedaan karakteristik air dan sedimen, seperti air jernih dan sedimen berpasir di Teluk Kosi dibandingkan dengan air keruh dan sedimen lanau-lempung di Nahoon dan Mngazana, kemungkinan merupakan faktor yang berkontribusi terhadap variasi yang diamati. Namun, penelitian di masa depan diperlukan untuk memverifikasi sejauh mana faktor-faktor ini menjelaskan keragaman komunitas muara.
Lebih jauh lagi, deteksi terbatas eDNA ikan dalam sedimen, bersama dengan sinyal eDNA homogen yang diamati antara sampel akuatik dan sedimen, menunjukkan bahwa pengambilan sampel pada tahap pasang surut yang berbeda memiliki dampak minimal pada variasi yang diamati dalam kekayaan eDNA dan komposisi komunitas. Ini berbeda dengan studi yang melaporkan komunitas yang berbeda pada pasang surut yang berbeda (misalnya Ahn et al., 2020 ; Sevellec et al., 2024 ) dan juga akan sangat bergantung pada waktu antara peristiwa pengambilan sampel. Misalnya, Courtaillac et al. ( 2024 ) dalam sistem pesisir Afrika Selatan menunjukkan bahwa komunitas ikan dapat bervariasi sepanjang 24 jam. Kami berpendapat bahwa perbedaan dalam desain pengambilan sampel studi sehubungan dengan kedalaman kolom air (dangkal vs. dalam) dan zona yang diambil sampelnya (subtidal vs. intertidal) menyoroti mengapa hal ini seharusnya tidak memainkan peran penting dalam hasil yang diperoleh dalam studi ini. Sebaliknya, karena kedalaman zona pasang surut yang dangkal di area pengambilan sampel kami (~1 m), terjadi pencampuran dan homogenisasi sinyal eDNA yang signifikan yang difasilitasi oleh pasang surut diurnal dan pergerakan air (Courtaillac et al., 2024 ; Rossouw et al., 2024 ).
4.2 Metabarkode DNA lingkungan tidak cocok untuk mendeteksi variasi komunitas pada skala spasial kecil di habitat pasang surut.
Efek homogenisasi potensial dari transpor eDNA melalui konektivitas hidrologi menimbulkan tantangan signifikan untuk menginterpretasikan data metabarkode dalam ekosistem pesisir dinamis (Jeunen et al., 2020 ; Rossouw et al., 2024 ; Sevellec et al., 2024 ). Temuan kami menunjukkan bahwa sinyal eDNA dari keanekaragaman iktiofauna tidak menunjukkan struktur yang jelas di seluruh habitat yang hidup berdampingan dalam skala spasial kecil (10 m). Kurangnya struktur komunitas dapat mengindikasikan luapan eDNA antar habitat, di mana bakau membentuk sabuk sempit di sepanjang tepian muara dan lamun berbatasan dengan bakau ini dan sangat mungkin bahwa air yang diambil sampelnya di bakau mengalir di atas lamun selama pasang surut. Transpor hidrologi eDNA yang disebabkan oleh pasang surut dan arus ini selanjutnya diilustrasikan oleh deteksi spesies yang terkait dengan terumbu tropis termasuk Odonus niger , Stethojulis spp. dan Rhinecanthus spp. dalam sampel dari Teluk Kosi, tempat terumbu karang berada di muara danau-muara (Harris et al., 1995 ). Secara keseluruhan, hasil kami menunjukkan bahwa metabarkode eDNA tidak cocok untuk mendeteksi perbedaan skala kecil dalam keanekaragaman dan komposisi komunitas ikan di ekosistem pesisir yang ditumbuhi vegetasi.
Hasil kami berkontribusi pada terbatasnya jumlah studi global yang menyelidiki keanekaragaman ikan di sistem muara pasang surut. Selain itu, desain pengambilan sampel skala besar kami berkontribusi pada studi regional yang mencakup rentang geografis yang luas (Czachur et al., 2022 ). Selain itu, dengan mencakup sebagian besar distribusi mangrove di Afrika Selatan, studi ini berkontribusi pada upaya berkelanjutan untuk mendokumentasikan keanekaragaman iktiofauna mangrove Afrika Timur, yang menandai batas paling selatan dari distribusinya (Adams & Rajkaran, 2021 ).
4.3 Tantangan eDNA metabarcoding di muara sungai Afrika bagian selatan
Seperti banyak survei keanekaragaman hayati eDNA yang dilakukan secara internasional (Wee et al., 2023 ) dan regional (Courtaillac et al., 2024 ; Czachur et al., 2022 ; Rossouw et al., 2024 ), kurangnya kode batang referensi dalam basis data membatasi identifikasi taksonomi dan oleh karena itu penerapan metakode batang eDNA sebagai alat biomonitoring. Dari komunitas yang dipulihkan, kurang dari 16% OTU ditetapkan sebagai spesies, yang menyoroti perlunya upaya berkelanjutan untuk membuat kode batang keanekaragaman hayati laut regional (Singh et al., 2021 ; von der Heyden, 2023 ). Di Afrika Selatan, di mana ~36% spesies laut bersifat endemik, upaya pemberian kode batang khusus wilayah sangatlah penting (Griffiths et al., 2010 ; Griffiths & Robinson, 2016 ), termasuk untuk ikan yang umumnya memiliki basis data 12S rRNA yang tidak lengkap (Czachur et al., 2022 ). Selain itu, penugasan yang terbatas kemungkinan mencerminkan keragaman yang kaya yang didukung oleh muara dan menunjukkan ruang lingkup pengambilan sampel dan upaya taksonomi ikan secara berkelanjutan di sepanjang pantai timur Afrika Selatan (von der Heyden, 2011).
Tantangan utama selama pengumpulan eDNA akuatik, terutama di lingkungan estuari, adalah penyumbatan filter yang cepat karena partikel tersuspensi (Kumar et al., 2022 ; Naputo et al., 2024 ), yang dapat mengakibatkan volume yang dikumpulkan rendah dan bervariasi. Lebih jauh lagi, keberadaan inhibitor PCR, seperti bahan organik, dalam air estuari yang keruh dan sedimen telah berkontribusi terhadap amplifikasi PCR yang suboptimal atau tidak berhasil (Ruan et al., 2022 ). Inhibitor ini dapat mengganggu proses PCR dan selanjutnya kedalaman sekuensing untuk banyak sampel. Mengingat volume air yang diambil sampelnya tidak mencukupi (Bessey et al., 2020 ) dan amplifikasi PCR DNA yang suboptimal untuk banyak sampel (Alberdi et al., 2018 ; Shirazi et al., 2021 ) akan menghasilkan banyak deteksi negatif palsu, keragaman yang ditemukan dalam survei ini kemungkinan meremehkan keragaman iktiofauna lokal.
Studi masa depan dapat lebih diuntungkan dari peningkatan replikasi teknis (lihat misalnya Courtaillac et al., 2024 ) pada area spasial yang lebih besar di dalam muara, seperti yang terlihat dalam studi yang mengambil sampel transek di sepanjang gradien lingkungan muara untuk menangkap keanekaragaman lokal secara lebih efektif (misalnya Ahn et al., 2020 ; Garcia-Vazquez et al., 2021 ; Gibson et al., 2023 ). Strategi ini juga diharapkan dapat meningkatkan deteksi spesies yang tidak terdeteksi dalam survei ini, termasuk spesies yang terdapat di bagian tengah dan hulu muara, misalnya Gilchristella aestuaria , ikan paling melimpah di muara Afrika bagian selatan (Whitfield, 2024 ).
Demikian pula, kami mendeteksi berbagai jenis pari dan ikan pari tetapi tidak ada hiu, beberapa di antaranya tersebar luas di muara yang diambil sampelnya (Heemstra et al., 2022 ). Studi di tempat lain juga melaporkan cakupan yang buruk dari spesies yang relatif langka seperti elasmobranchs, kemungkinan karena kombinasi faktor termasuk ketersediaan kode batang DNA voucher, pengujian metabarkode yang tepat (Asbury et al., 2021 ; Burian et al., 2023 ; Zhu et al., 2023 ) dan dampak dari tingkat pelepasan eDNA yang lebih rendah dibandingkan dengan ikan bertulang (Merson et al., 2025 ).
5 KESIMPULAN
Mengingat terbatasnya jumlah studi eDNA metabarcoding yang telah dilakukan di ekosistem estuari (Czachur et al., 2022 ), bersama dengan kebutuhan untuk standardisasi dan optimalisasi alat molekuler ini (von der Heyden, 2023 ), kami bertujuan untuk berkontribusi pada badan penelitian eDNA yang terus berkembang di Afrika Selatan. Studi ini menunjukkan bahwa eDNA metabarcoding dapat secara efektif menangkap keanekaragaman iktiofauna yang tinggi, termasuk 50 famili yang terkait dengan estuari dan wilayah pesisir di dekatnya. Temuan kami menunjukkan bahwa eDNA metabarcoding akuatik lebih efektif untuk mendeteksi spesies ikan dibandingkan dengan pengambilan sampel sedimen, yang menyoroti pentingnya mempertimbangkan jenis sampel eDNA dalam desain protokol pengambilan sampel. Selain itu, eDNA metabarcoding saat ini tidak cocok untuk membedakan antara keanekaragaman ikan di ekosistem mangrove dan lamun yang berdekatan. Optimalisasi dan standarisasi metabarkode eDNA yang berkelanjutan, bersamaan dengan upaya barkode untuk wilayah gen 12S rRNA, diharapkan dapat meningkatkan pemantauan resolusi tinggi terhadap keanekaragaman hayati muara dan meningkatkan deteksi pergeseran distribusi fauna skala besar (>10 km) yang didorong oleh tekanan perubahan global (James et al., 2013 , 2016 ; van Niekerk et al., 2022 ).


