
ABSTRAK
Penyelamatan genetika merupakan strategi pengelolaan konservasi yang mengurangi dampak negatif dari pergeseran genetika dan perkawinan sedarah pada populasi kecil dan terisolasi. Namun, populasi tersebut mungkin sudah rentan terhadap fluktuasi acak dalam laju pertumbuhan (stokastik demografi). Oleh karena itu, keberhasilan penyelamatan genetika tidak hanya bergantung pada komposisi genetika populasi sumber dan target, tetapi juga pada hasil yang muncul dari interaksi proses demografis dan peristiwa stokastik lainnya. Oleh karena itu, pengembangan model prediktif yang memperhitungkan umpan balik antara proses demografis dan genetika (‘umpan balik demo-genetik’) diperlukan untuk memandu penerapan penyelamatan genetika guna meminimalkan risiko kepunahan populasi yang terancam. Di sini, kami menjelaskan bagaimana penguatan timbal balik dari pergeseran genetika, perkawinan sedarah, dan stokastik demografis meningkatkan risiko kepunahan pada populasi kecil. Kami kemudian menjelaskan bagaimana proses ini dapat dimodelkan dengan membuat parameterisasi mekanisme yang mendasarinya, termasuk mutasi yang merusak dengan dominasi parsial dan laju demografis dengan varians yang meningkat seiring dengan penurunan kelimpahan. Kami menggabungkan saran kami tentang parameterisasi model dengan perbandingan kemampuan dan fleksibilitas yang relevan dari lima program sumber terbuka yang dirancang untuk membangun simulasi berbasis individu yang eksplisit secara genetika. Dengan menggunakan salah satu program, kami menyediakan model heuristik untuk menunjukkan bahwa penyelamatan genetik yang disimulasikan dapat menunda kepunahan populasi virtual kecil yang jika tidak demikian akan terpapar pada risiko kepunahan yang lebih besar karena umpan balik demo-genetik. Kami kemudian menggunakan studi kasus marsupial Australia yang terancam untuk menunjukkan bahwa data genetik yang dipublikasikan dapat digunakan dalam satu atau semua tahap pengembangan dan penerapan model, termasuk parameterisasi, kalibrasi, dan validasi. Kami menyoroti bahwa penyelamatan genetik dapat disimulasikan dengan variasi urutan virtual atau empiris (atau pendekatan hibrida) dan menyarankan bahwa pengambilan keputusan berbasis model harus diinformasikan dengan memberi peringkat sensitivitas probabilitas/waktu kepunahan yang diprediksi terhadap variasi parameter model (misalnya, ukuran translokasi, frekuensi, populasi sumber) di antara berbagai skenario penyelamatan genetik.
1 Pendahuluan
Banyak spesies yang terancam hanya bertahan hidup sebagai populasi kecil dan terisolasi di mana efek gabungan dari perkawinan sedarah dan pergeseran genetik dapat mengakumulasi mutasi yang merugikan dan mengurangi keragaman genetik, bersama-sama meningkatkan risiko kepunahan (Frankham 2005 ; Kardos et al. 2021 ). Selain perlindungan dan pemulihan habitat, konservasi yang berhasil dari banyak spesies yang terancam mungkin memerlukan pergerakan yang disengaja dari individu-individu yang berdiferensiasi secara genetik dari satu populasi ke populasi lain untuk meningkatkan keragaman genetik dan kebugaran populasi target—proses yang dikenal sebagai penyelamatan genetik (Glosarium—Tabel 1 ; Bell et al. 2019 ; Fitzpatrick et al. 2023 ; Ralls et al. 2020 ; Whiteley et al. 2015 ). Namun, penyelamatan genetik biasanya diterapkan sebagai intervensi darurat untuk mencegah kepunahan populasi kecil atau yang menurun yang mungkin sudah rentan terhadap fluktuasi acak dalam tingkat pertumbuhan yang dapat menyebabkan keruntuhan populasi secara tiba-tiba (Melbourne dan Hastings 2008 ; Bell et al. 2019 ; Fitzpatrick et al. 2023 ; Ralls et al. 2020 ; Whiteley et al. 2015 ). Oleh karena itu, keberhasilan penyelamatan genetik dalam konservasi bergantung tidak hanya pada komposisi genetik dan riwayat demografi populasi sumber dan target tetapi juga pada hasil yang muncul dari proses demografi yang berinteraksi (dan peristiwa lingkungan stokastik seperti kekeringan, kebakaran, dll.). Oleh karena itu, mengembangkan model prediktif yang memperhitungkan umpan balik antara proses demografi dan genetik (‘umpan balik demo-genetik’) diperlukan untuk memandu implementasi penyelamatan genetik untuk meminimalkan risiko kepunahan populasi yang terancam.
| Ketentuan | Definisi | Referensi |
|---|---|---|
| Alel | Masing-masing dari dua atau lebih bentuk alternatif gen yang muncul melalui mutasi dan ditemukan di tempat yang sama pada kromosom. | |
| Efek Allee | Pergeseran dari kompensasi (hubungan negatif antara laju pertumbuhan per kapita dan kelimpahan) ke depensasi (hubungan positif antara laju pertumbuhan per kapita dan kelimpahan) pada ukuran populasi kecil disebabkan oleh antara lain depresi perkawinan sedarah, erosi jaringan sosial, kesempatan kawin atau pemeliharaan yang tidak memadai. | Courchamp dkk. 1999 ; Herrando-Pérez dkk. 2012 |
| Jumlah penduduk sensus ( N c ) | Jumlah individu dalam suatu populasi, terlepas dari status reproduksi atau kontribusi genetik mereka terhadap generasi mendatang (dibandingkan dengan ukuran populasi efektif ). | Frankham dkk. 2019 |
| Alel yang merugikan | Versi gen yang, rata-rata, menurunkan kebugaran dalam lingkungan individu saat ini. | |
| Umpan balik demo-genetik | Efek timbal balik dari proses demografi (misalnya, umpan balik kepadatan, stokastisitas demografi) pada proses genetik populasi (misalnya, pergeseran genetik, seleksi, aliran gen) yang bersama-sama menentukan pertumbuhan populasi, keragaman genetik, dan beban genetik. | |
| Stokastisitas demografi | Varian dalam pertumbuhan populasi karena sifat peluang pada kelahiran, kematian, dan migrasi individu. | Melbourne dan Hastings 2008 |
| Umpan balik kepadatan (~ ketergantungan kepadatan) | Ketika interaksi sosial dan trofik mengubah laju demografi dan perubahan laju demografi yang dihasilkan mengubah kepadatan populasi, ‘memberikan umpan balik’ untuk mengubah intensitas interaksi tersebut. | Bradshaw dan Herrando-Pérez 2023 |
| Dominasi | Penyimpangan fenotipe individu yang heterozigot pada lokus tertentu dari fenotipe rata-rata individu homozigot. Dominasi lengkap terjadi ketika fenotipe heterozigot tidak dapat dibedakan dari fenotipe induk homozigot. Dominasi parsial (atau tidak lengkap) terjadi ketika fenotipe heterozigot berada di antara fenotipe kedua induk homozigot (salah satunya adalah homozigot dominan dan yang lainnya homozigot resesif). | Frankham dkk. 2019 |
| Beban melayang | Pengurangan kebugaran rata-rata disebabkan oleh peningkatan stokastik dalam frekuensi mutasi (yang biasanya lemah atau cukup merugikan) pada populasi kecil. | Lynch dkk. 1995 ; Whitlock 2000 |
| Epistasis | Ketergantungan efek substitusi gen pada latar belakang genetik. Secara umum, interaksi antar gen pada lokus yang berbeda ( lih . alel pada lokus yang sama, lihat dominasi ).
Definisi ini mengacu pada ‘epistasis fungsional’ ( sensu Hansen 2013 ) lih . epistasis statistik ( sensu Walsh dan Lynch 2018 ). |
Hansen tahun 2013 |
| Beban genetik | Akumulasi mutasi yang merugikan dalam suatu populasi. Dalam hal kebugaran, beban genetik adalah fraksi perbedaan rata-rata populasi dari genotipe referensi (yaitu, genotipe dengan kebugaran maksimum). Secara matematis, beban genetik = beban yang terwujud + beban yang tersamar (= jumlah koefisien seleksi dari semua mutasi yang merugikan). | Dussex dkk. 2023 |
| Ukuran populasi efektif ( N e ) | Jumlah individu yang akan mengakibatkan hilangnya keragaman genetik, perkawinan sedarah, pergeseran genetik, atau penggabungan jika mereka berperilaku seperti populasi ideal. Perlu dicatat, N e sering kali lebih kecil daripada ukuran populasi sensus karena faktor-faktor seperti rasio jenis kelamin yang tidak sama, variasi dalam keberhasilan reproduksi, dan fluktuasi populasi. | Frankham dkk. 2019 |
| Penyelamatan genetik | Peningkatan kebugaran populasi (diukur paling baik berdasarkan laju pertumbuhan populasi) karena aliran gen. Penyelamatan genetik dapat terjadi karena aliran gen alami atau terbantu. Dalam konteks konservasi spesies yang terancam, penyelamatan genetik dapat didefinisikan secara lebih sempit sebagai pengenalan genetik yang disengaja yang bertujuan untuk menutupi alel yang merusak yang bertanggung jawab atas beban genetik dalam populasi kecil yang menyebabkan peningkatan laju pertumbuhan populasi. | Whiteley dkk. 2015 ; Hoffmann dkk. 2021 |
| Depresi akibat perkawinan sedarah | Penurunan kebugaran individu dengan orang tua terkait. Berbeda dengan perkawinan sedarah, yaitu perkawinan antara individu yang memiliki hubungan lebih dekat daripada pasangan individu yang dipilih secara acak dalam suatu populasi. | Kardos dkk. 2021 |
| Lokus, lokus (jamak) | Posisi gen atau penanda genetik pada kromosom. | |
| Rekombinasi | Proses di mana pasangan kromosom bertukar DNA satu sama lain selama pembentukan gamet. Rekombinasi menyatukan kombinasi gen baru—sumber variasi yang menjadi dasar seleksi alam. | |
| Model Wright-Fisher | Model rantai Markov waktu diskrit dari frekuensi alel dalam populasi terbatas dengan ukuran konstan, dengan asumsi perkawinan acak, generasi yang tidak tumpang tindih, tidak ada mutasi, tidak ada migrasi, dan tidak ada seleksi. |
Bidang ekologi dan genetika populasi masing-masing memiliki rangkaian model deterministik (respons rata-rata) dan stokastik yang kaya yang telah memajukan prediksi dinamika populasi alami dan keragaman genetiknya, termasuk elemen umpan balik demo-genetik (Govaert et al. 2019 ; Nordstrom et al. 2023 ). Tetapi menggabungkan umpan balik demo-genetik ke dalam model terapan penyelamatan genetik merupakan tantangan karena biasanya memerlukan pendekatan yang mahal secara komputasi dan tentu saja rumit (yaitu, kaya parameter dan haus data). Untungnya, kemajuan cepat (dan komplementer) dalam daya komputasi, teknologi sekuensing, dan perangkat lunak proyeksi maju yang eksplisit secara genetik memfasilitasi pengembangan model simulasi canggih yang diparameterisasi dan divalidasi dengan lebih banyak data atau berdasarkan asumsi yang lebih realistis daripada yang sebelumnya memungkinkan (Haller dan Messer 2019 ; Carey et al. 2019 ; Carturan et al. 2020 ; Pilowsky et al. 2022 ). Kemajuan pelengkap ini memungkinkan pengembangan dan penerapan model proses-eksplisit yang diperlukan untuk membuat prediksi yang lebih akurat tentang dinamika populasi satwa liar di bawah intervensi pengelolaan yang diusulkan seperti penyelamatan genetik.
Di sini, kami mengeksplorasi pengembangan dan penerapan model komputasional untuk menginformasikan penyelamatan genetik dalam konservasi. Pertama-tama kami merangkum literatur teoritis dan empiris tentang proses demografi dan genetik yang relevan dengan penyelamatan genetik, dan kemudian menyarankan pendekatan untuk membuat parameterisasi model yang menggabungkan mekanisme umpan balik demo-genetik untuk meningkatkan prediksi. Kami menguraikan secara singkat kemampuan relatif dari lima program komputer sumber terbuka yang ada yang dapat digunakan untuk simulasi penyelamatan genetik. Dengan menggunakan salah satu program ini, kami mengembangkan model heuristik sebagai bukti konsep untuk pengaruh umpan balik demo-genetik pada hasil simulasi penyelamatan genetik. Kami kemudian fokus pada marsupial Australia yang terancam sebagai studi kasus dan membahas bagaimana data genetik yang dipublikasikan dapat digunakan dalam pengembangan dan penerapan model (yaitu, parameterisasi, kalibrasi, dan validasi). Kami menyimpulkan dengan menguraikan arah strategis untuk penelitian masa depan dan penerapan pengambilan keputusan berbasis model dalam penyelamatan genetik.
2 Umpan Balik Demo-Genetik dalam Penyelamatan Genetik
Intervensi penyelamatan genetik bertujuan untuk mengurangi kemungkinan kepunahan populasi kecil dan menurun dengan melawan ancaman genetik terhadap kebugaran dan pertumbuhan populasi (Fitzpatrick et al. 2023 ). Namun, populasi yang menjadi target penyelamatan genetik biasanya rentan terhadap kombinasi proses genetik dan demografi stokastik, penguatan timbal balik yang menciptakan lingkaran umpan balik positif (umpan balik demo-genetik) yang meningkatkan risiko kepunahan saat populasi menurun—sebuah fenomena yang disebut sebagai ‘pusaran kepunahan’ (Caughley 1994 ; Fagan dan Holmes 2006 ; Gilpin dan Soulé 1986 ). Akibatnya, sementara penyelamatan genetik dapat segera meningkatkan kebugaran, keuntungan ini mungkin berumur pendek jika ketidakstabilan demografis berlanjut, menangkal manfaat intervensi genetik dan menarik populasi kembali ke pusaran kepunahan. Oleh karena itu, memaksimalkan efektivitas intervensi penyelamatan genetik memerlukan pemahaman yang komprehensif tentang umpan balik demo-genetik dan perannya dalam membentuk hasil penyelamatan genetik. Pada bagian ini, kami fokus pada mekanisme umpan balik demo-genetik dan menyoroti relevansinya terhadap penyelamatan genetik. Pada bagian berikut, kami menjelaskan pendekatan yang kami sarankan untuk membangun model simulasi berbasis individu dari umpan balik demo-genetik dan bagaimana pendekatan tersebut dapat diterapkan untuk mengevaluasi berbagai skenario intervensi penyelamatan genetik.
Populasi kecil sangat dipengaruhi oleh proses stokastik yang muncul pada tingkat populasi dari mekanisme genetik dan individu yang mendasarinya. Sifat kelahiran dan kematian yang terpisah dan biner memperkenalkan keacakan ke dalam tingkat demografi (misalnya, kelangsungan hidup dan kesuburan) dan fenomena yang muncul (misalnya, rasio jenis kelamin, usia kematangan reproduksi), yang menyebabkan fluktuasi stokastik dalam tingkat pertumbuhan populasi dan kelimpahan (stochasticity demografi). Fluktuasi dalam kelimpahan menimbulkan sedikit ancaman bagi populasi yang lebih besar karena penurunan kepadatan cenderung meningkatkan kebugaran rata-rata (misalnya, melalui berkurangnya persaingan), yang mengarah pada peningkatan kompensasi dalam tingkat pertumbuhan populasi per kapita (kompensasi, Herrando-Pérez et al. 2012 ) . Namun, pada populasi yang lebih kecil, stokastisitas demografi dapat menyebabkan penurunan kelimpahan di bawah ambang batas di mana mereka menjadi depensatory — ketika penurunan kepadatan berikutnya menurunkan kebugaran rata-rata dan menimbulkan penurunan lebih lanjut (Gambar 1 ). Penurunan populasi yang bergantung dapat disebabkan oleh mekanisme ekologi dan perilaku (misalnya, keterbatasan pasangan, gangguan struktur sosial), fenomena yang diketahui menimbulkan apa yang juga disebut sebagai efek Allee (Courchamp et al. 1999 ). Depensasi juga dapat disebabkan oleh mekanisme genetik (misalnya, pergeseran genetik dan perkawinan sedarah) yang dapat dianggap mirip dengan mekanisme genetik yang mendasari efek Allee (Luque et al. 2016 ). Mekanisme genetik dari depensasi populasi inilah yang secara khusus ingin diimbangi oleh penyelamatan genetik.
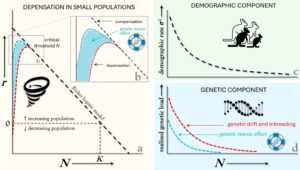
Seperti halnya fluktuasi stokastik dalam tingkat demografi, sifat diskret dan biner dari kelahiran dan kematian individu mendukung pergeseran genetik acak frekuensi alel dalam populasi. Karena pengaruh kuat pergeseran genetik dalam populasi kecil, mutasi yang lemah atau cukup merusak dapat terakumulasi dan menjadi tetap, sehingga mengurangi kebugaran populasi rata-rata (beban pergeseran; Angst et al. 2022 ; Whitlock 2000 ). Perkawinan sedarah juga menjadi lebih umum ketika populasi menurun, dan meningkatkan kemungkinan bahwa mutasi resesif yang merusak akan diekspresikan pada individu homozigot untuk mengurangi kebugaran mereka (yaitu, depresi perkawinan sedarah) (Charlesworth dan Willis 2009 ; Frankham 2005 ; Frankham et al. 2019 ; O’Grady et al. 2006 ). Dengan meningkatkan frekuensi alel yang merusak dan mengurangi keragaman genetik, pergeseran genetik dan perkawinan sedarah mengubah beban genetik terselubung yang sebelumnya tersembunyi dalam keadaan heterozigot menjadi beban genetik yang nyata, yang pada akhirnya mengurangi pertumbuhan dan ketahanan populasi (Dussex et al. 2023 ; Mathur dan DeWoody 2021 ). Mutasi yang sangat merusak cenderung tidak terakumulasi melalui pergeseran genetik karena kontribusinya terhadap depresi perkawinan sedarah cenderung mendorong penghapusannya melalui seleksi alam (pembersihan genetik), bahkan dalam populasi kecil (Dussex et al. 2023 ; Grossen et al. 2020 ; Hedrick dan Garcia-Dorado 2016 ). Peran mutasi yang sangat merusak dan pembersihan genetik dalam keberhasilan penyelamatan genetik merupakan topik perdebatan terkini dan kontroversial (Kyriazis et al. 2021 ; Pérez-Pereira et al. 2022 ; Ralls et al. 2020 ; Robinson et al. 2018 ; Robinson et al. 2021 ), dan merupakan area penelitian di mana model simulasi berpotensi memberikan kontribusi yang berharga.
Bahasa Indonesia: Untuk mengurangi ancaman yang ditimbulkan oleh beban genetik yang terwujud terhadap populasi kecil dan menurun, intervensi penyelamatan genetik sengaja memperkenalkan individu heterozigot dan berdiferensiasi secara genetik ke dalam populasi sasaran dalam upaya untuk menutupi kembali alel-alel yang merusak yang berkontribusi terhadap beban genetik, dengan demikian menimbulkan peningkatan laju pertumbuhan populasi (lihat Hoffmann et al. 2021 untuk tinjauan berbagai bentuk pencampuran genetik dalam konservasi). Keberhasilan penyelamatan genetik bergantung pada apakah menutupi beban genetik meningkatkan kebugaran rata-rata yang cukup untuk meningkatkan kelimpahan populasi di atas ambang batas di mana pertumbuhan populasi beralih dari depensasi menjadi kompensasi (Gambar 1 ). Model berbasis individu menyediakan alat yang ampuh dan praktis untuk membandingkan dan memberi peringkat skenario alternatif intervensi penyelamatan genetik berdasarkan hasil penyelamatan genetik virtual dalam populasi simulasi. Model yang menggabungkan mekanisme umpan balik demo-genetik yang mendorong dinamika depensatori dapat digunakan oleh praktisi konservasi untuk membuat proyeksi ke depan dari dampak memperkenalkan individu yang berdiferensiasi secara genetik (virtual) pada beban genetik yang terwujud dan laju pertumbuhan populasi (simulasi). Pada bagian berikut, kami menyarankan pendekatan untuk mengembangkan model demo-genetik yang dapat digunakan untuk mensimulasikan penyelamatan genetik, dan memberikan gambaran singkat tentang perangkat lunak sumber terbuka dan kemampuannya yang relevan.
3 Membangun Simulasi Demo-Genetik Penyelamatan Genetik
3.1 Menghubungkan Mekanisme Umpan Balik ke Parameter Model
Alat komputasi telah maju ke tempat yang sekarang memungkinkan untuk mengembangkan simulasi praktis untuk merencanakan dan mengimplementasikan skenario penyelamatan genetik. Model berbasis individu yang eksplisit secara genetik adalah yang paling menjanjikan untuk menggabungkan mekanisme umpan balik demo-genetik untuk mengevaluasi probabilitas keberhasilan penyelamatan genetik populasi kecil dan terisolasi. Program perangkat lunak sumber terbuka yang mampu mensimulasikan pengaruh umpan balik demo-genetik pada hasil penyelamatan genetik termasuk SLiM (Haller dan Messer 2023 ), quantiNemo (Neuenschwander et al. 2019 ), perluasan dari perangkat lunak Nemo asli (Guillaume dan Rougemont 2006 ), CDMetaPOP (Day et al. 2023 ), RangeShifter (Bocedi et al. 2021 ), dan HexSim (Schumaker dan Brookes 2018 ). Meskipun kemungkinan ada model khusus lainnya, kelima platform ini berisi sebagian besar proses yang diperlukan untuk mengintegrasikan proses demo-genetik dalam skenario penyelamatan genetik. Di sini, kami mulai dengan menjelaskan jenis parameter yang dapat ditentukan dalam simulasi penyelamatan genetik untuk memungkinkan munculnya umpan balik demo-genetik dari mekanisme yang mendasarinya (Tabel 2 ). Kami kemudian memberikan deskripsi singkat dari masing-masing dari lima platform ini dan merangkum kemampuan mereka untuk membuat parameterisasi mekanisme umpan balik demo-genetik (Tabel 3 ).
| Mekanisme komponen | Umpan balik dengan berlimpah | Efek pada kebugaran | Parameter model yang relevan |
|---|---|---|---|
| Stokastisitas demografi (varian acak dalam tingkat pertumbuhan populasi) | Meningkatnya varians dalam tingkat demografi komponen (kelangsungan hidup, kesuburan) seiring dengan menurunnya kelimpahan | Tidak tergantung pada kebugaran | Tingkat demografi: khususnya, parameter yang digunakan untuk menyebabkan peningkatan varians dalam tingkat demografi seiring dengan menurunnya kelimpahan (lihat Informasi Pendukung ) |
| Pergeseran genetik | Meningkatnya pengaruh pergeseran genetik pada populasi kecil | Menyebabkan akumulasi dan fiksasi alel yang merugikan | Frekuensi alel populasi: dihasilkan melalui mutasi de novo virtual selama simulasi burn-in, atau frekuensi alel atau data sekuens yang ditentukan pengguna pada inisialisasi model
Tingkat rekombinasi Koefisien dominasi alel untuk mutasi/alel yang merugikan Efek kebugaran dari mutasi/alel yang merugikan |
| Kawin sedarah | Meningkatnya kejadian perkawinan sedarah seiring dengan menurunnya kelimpahan populasi | Mengurangi kebugaran individu karena homozigositas pada lokus yang merugikan (depresi perkawinan sedarah);
Mengurangi kebugaran rata-rata karena homozigositas pada lokus yang merugikan (beban genetik yang terwujud) |
Kejadian perkawinan sedarah pada populasi virtual merupakan proses yang muncul dalam model berbasis individu yang mencakup reproduksi seksual.
Pengaruh perkawinan sedarah terhadap prevalensi depresi perkawinan sedarah dan beban genetik yang direalisasikan dimediasi oleh frekuensi alel, tingkat rekombinasi, dominasi, dan efek kebugaran |
| Pembersihan genetik | Kemanjuran seleksi dalam pembersihan alel yang merugikan biasanya lebih lemah pada populasi yang lebih kecil (karena meningkatnya pengaruh pergeseran); perkawinan sedarah dalam populasi kecil juga dapat meningkatkan kemanjuran pembersihan | Meningkatkan kebugaran rata-rata karena penghapusan alel yang merugikan dari populasi | Frekuensi alel
Tingkat rekombinasi Dominasi alelik Efek kebugaran |
| Kemampuan | Fleksibilitas | Langsing | quantiNemo sebuah | CDMetaPop | Penggeser Jangkauan | HexSim |
|---|---|---|---|---|---|---|
| Genetika | ||||||
| Mutasi de novo | Tarif tunggal | Ya | Ya | Ya | Ya | Ya |
| Beberapa tarif | Ya | Ya | Ya | TIDAK | Ya | |
| Alel virtual | Abstrak b | Ya | Ya | Ya | Ya | Ya |
| Urutan yang dipetakan c | Ya d | Ya benar | TIDAK | TIDAK | Ya f | |
| Alel empiris | Berdasarkan genotipe | — | — | Ya | Ya | Ya |
| Input file genotipe | Ya g | Ya , h | Ya | TIDAK | TIDAK | |
| Urutan yang dipetakan | Ya aku | Ya j | TIDAK | TIDAK | TIDAK | |
| Efek kebugaran dari mutasi | Relatif k | Ya | Ya | Ya aku | Ya saya | |
| Absolut n | Ya | Ya | TIDAK | TIDAK | ||
| Nilai titik | — | — | Ya | — | ||
| Distribusi (tetap) | — | — | TIDAK | Ya | ||
| Distribusi (Ditentukan pengguna) | Ya | Ya | TIDAK | TIDAK | ||
| Rekombinasi | Lintasan | Ya | Ya | TIDAK | Ya | Ya |
| Tidak berpindah jalur | Ya | Ya | TIDAK | TIDAK | TIDAK | |
| Tarif tunggal | — | — | TIDAK | Ya | ||
| Beberapa tarif | Ya | Ya | TIDAK | TIDAK | ||
| Dominasi (alel) | Ya | Ya | TIDAK | TIDAK | TIDAK | |
| Demografi | ||||||
| Sumber varians dalam probabilitas kelangsungan hidup individu | Lingkungan | Ya | Ya | Ya | Ya | Ya |
| Genotip | Ya | Ya | Ya | TIDAK | ||
| Acak | Ya | TIDAK | Ya | Ya | ||
| Tahapan khusus | Ya | TIDAK | Ya | Ya | Ya | |
| Sumber varians dalam tingkat kesuburan individu | Lingkungan | Ya | Ya | Ya | Ya | Ya |
| Efek genotipe | Ya | Ya | Ya | Tidak o | ||
| Acak | Ya | Ya | Ya | Ya | ||
| F (kitaD2, N ) | R (kitaD2Bahasa Indonesia:akuakuD) ≠ 0 | — | TIDAK | TIDAK | Ya | TIDAK |
| R (kitaD2Bahasa Indonesia:akuakuD) = 0 | Ya | TIDAK | TIDAK | TIDAK | TIDAK | |
| Tingkat demografi spesifik tahap | Kelangsungan hidup | Ya | Ya | Ya | Ya | Ya |
| Kesuburan | Ya | TIDAK | Ya | Ya | Ya | |
| Rasio jenis kelamin | Tetap | — | — | — | — | |
| Ditentukan pengguna | Ya | Ya | Ya | Ya | ||
| Generasi | Tidak tumpang tindih | — | Ya | Ya | Ya | — |
| Tumpang tindih | Ya | TIDAK | Ya | Ya | Ya | |
| Spasialitas | Kesesuaian habitat | Ya | Ya | Ya | Ya | Ya |
| Permeabilitas lanskap | Ya | Ya | Ya | Ya | Ya | |
| Bubaran | Berbasis kernel | Ya | Ya | Ya | Ya | Ya |
| Berbasis jalur | TIDAK | TIDAK | Ya | Ya | Ya | |
Perpanjangan dari Nemo dan terkait dengan Nemo-age .
b Alel virtual digambarkan sebagai abstrak jika ia mempunyai sifat-sifat seperti koefisien seleksi atau posisi dalam genom, tetapi tidak secara eksplisit mewakili urutan nukleotida (virtual).
c Alel virtual ditentukan dengan beberapa bentuk lokasi fisik.
d Genom virtual.
e Peta rekombinasi.
f Jenis peta hubungan virtual tanpa unit.
berkas VCF.
h berkas FSTAT.
berkas FASTA.
j berkas Arlequin.
k Kebugaran bervariasi relatif terhadap konteks lingkungan dan/atau individu lain.
l Efek kebugaran relatif diimplementasikan melalui lanskap kebugaran untuk model diallelik (lokus tunggal atau dua lokus).
m Efek kebugaran diimplementasikan melalui probabilitas kelangsungan hidup selama tiga tahap penyebaran (emigrasi, transfer, pemukiman) dan efek kepadatan dan habitat spesifik pada tingkat kelangsungan hidup dan kesuburan.
n Berguna untuk pemodelan beban genetik.
o Meskipun variasi dalam sifat penyebaran, yang dapat diwariskan, secara tidak langsung memengaruhi variasi dalam tingkat kesuburan melalui kesuburan yang bergantung pada kepadatan, yang bervariasi secara spasial.
Model berbasis individu untuk simulasi penyelamatan genetik memiliki keuntungan dalam mereplikasi sifat diskrit dan biner dari kelahiran, kematian, imigrasi, dan emigrasi. Hal ini memperkenalkan keacakan (yaitu, varians) ke dalam probabilitas kelangsungan hidup dan fertilitas, dan sifat demografis yang muncul lainnya seperti rasio jenis kelamin dan struktur usia, yang mendukung fluktuasi stokastik dalam tingkat pertumbuhan per kapita ( r ) dan kelimpahan ( N ) (Gambar 2 ). Demikian pula, model berbasis individu yang eksplisit secara genetik dapat mensimulasikan akumulasi stokastik alel yang merusak melalui pergeseran genetik, dan kontribusinya terhadap depresi perkawinan sedarah dan beban genetik yang terwujud (meskipun platform pemodelan berbeda dalam detail spesifik tentang bagaimana proses ini direpresentasikan, yang dijelaskan di bawah ini pada bagian tentang kemampuan perangkat lunak). Misalnya, proses genetik stokastik dapat dimodelkan dengan menetapkan parameter untuk tingkat mutasi (atau frekuensi alel yang diinisialisasi), tingkat rekombinasi, dominasi alel, dan efek kebugaran mutasi atau alel (baik yang diambil dari distribusi probabilitas atau tetap), termasuk efek netral, menguntungkan, atau merusak. Ada batasan pada parameter mana yang dapat dikalibrasi dan divalidasi dengan data empiris, dan kami membahas topik ini di bawah ini menggunakan studi kasus marsupial Australia yang terancam dalam Penggunaan data untuk memandu pengembangan dan penerapan model .
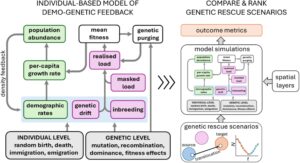
Program perangkat lunak yang tersedia mampu memodelkan berbagai macam proses ekologi dan evolusi. Oleh karena itu, simulasi skenario penyelamatan genetik untuk menginformasikan tindakan konservasi tertentu memerlukan pengambilan keputusan penting tentang parameter mana yang akan disertakan atau dikecualikan untuk mengembangkan model parsimonius yang menangkap dinamika penting sistem (lihat García-Díaz et al. 2019 untuk panduan ringkas tentang pengembangan dan penggunaan model kuantitatif dalam manajemen konservasi). Kami berpendapat bahwa implementasi penyelamatan genetik dalam konservasi harus dipandu oleh model yang mereplikasi penguatan timbal balik dari stokastisitas demografi, pergeseran genetik, dan perkawinan sedarah (misalnya, Gambar 2 ). Proses-proses ini menyebabkan populasi menurun ke dalam pusaran kepunahan (Gambar 1 ) dan menciptakan ketidakpastian bagi para pembuat keputusan yang memilih di antara serangkaian intervensi potensial yang bersaing.
3.2 Mengintegrasikan Dinamika Spasial dan Lingkungan
Sebagian besar populasi menunjukkan variasi spasial baik dalam tingkat demografis maupun sifat genomik (White dan Smith 2018 ), sehingga model penyelamatan genetik yang eksplisit secara spasial cenderung lebih realistis daripada model non-spasial. Lebih jauh lagi, individu yang ditranslokasi dari populasi sumber ke populasi target mungkin cenderung menyebar secara berlebihan dari lokasi pelepasan (Bilby dan Moseby 2024 ), dengan implikasi untuk pencampuran genetik berikutnya. Model spasial dapat memperhitungkan dinamika penyebaran (misalnya, kecenderungan penyebaran dan jarak), yang mungkin dipengaruhi oleh variasi spasial dalam kesesuaian dan konfigurasi habitat, serta permeabilitas (atau resistensi) terhadap pergerakan.
3.3 Kemampuan Perangkat Lunak Open-Source yang Ada untuk Membangun Simulasi Demo-Genetik Penyelamatan Genetik
Di sini kami menyediakan deskripsi singkat dari lima program perangkat lunak sumber terbuka yang dapat digunakan untuk mengembangkan dan menjalankan simulasi demo-genetik penyelamatan genetik. Setiap program memiliki dokumentasi terperinci dan materi pendukung yang berguna bagi peneliti terapan dan praktisi konservasi. Ringkasan tertulis dari setiap program dimaksudkan untuk memberikan konteks daripada perbandingan terperinci tentang kemampuan dan fleksibilitas mereka untuk mengembangkan model demo-genetik untuk mensimulasikan penyelamatan genetik, yang kami sediakan dalam Tabel 3. Di sana, kami mengidentifikasi jenis parameter spesifik yang berkaitan dengan mekanisme dan proses yang kami jelaskan di bagian atas (yaitu, parameter yang memunculkan umpan balik demo-genetik dalam populasi virtual). Untuk setiap program, kami nyatakan apakah program tersebut memiliki atau tidak memiliki kemampuan atau fleksibilitas yang kami jelaskan untuk setiap jenis parameter. Misalnya, kami membedakan di antara program-program berdasarkan apakah mereka memodelkan mutasi/alel virtual sebagai varian ‘abstrak’ atau ‘urutan yang dipetakan’ (atau keduanya). Kami mengkategorikan mutasi/alel sebagai abstrak jika diperlakukan sebagai varian yang ditentukan oleh serangkaian properti (misalnya, koefisien seleksi dan dominasi), yang tidak mencakup lokasi fisik (misalnya, dalam genom virtual atau peta rekombinasi). Sebaliknya, kami mengkategorikan mutasi/alel sebagai urutan yang dipetakan jika diperlakukan sebagai varian nukleotida dengan lokasi yang ditentukan pada genom virtual atau peta rekombinasi. Ini adalah salah satu contoh pendekatan yang kami ambil untuk membandingkan program-program dalam Tabel 3. Kami mulai di bawah ini dengan ringkasan deskriptif singkat dari setiap program.
SLiM (‘ Pemilihan S pada Mutasi Terkait ‘; messerlab.org/slim ) (Haller dan Messer 2023 ) adalah simulator berbasis individu, maju dalam waktu yang dirancang untuk studi genetika evolusi. SLiM eksplisit secara genetik karena mensimulasikan mutasi pada posisi tertentu sepanjang genom virtual daripada frekuensi alel atau rata-rata dan varians sifat genetik kuantitatif. Secara default, SLiM memodelkan genom sebagai ‘batu tulis kosong’ dari posisi kromosom dan hanya mutasi yang dilacak untuk meningkatkan efisiensi komputasi. Namun, dimungkinkan untuk mensimulasikan urutan nukleotida empiris dalam SLiM menggunakan file FASTA (format berbasis teks untuk merepresentasikan urutan nukleotida sebagai kode huruf tunggal) dan VCF (format panggilan varian, file teks standar untuk menyimpan variasi urutan gen) untuk merepresentasikan urutan yang disimulasikan. Proses genetik lain yang dapat dimodelkan mencakup laju rekombinasi variabel di seluruh genom (meniru hubungan genetik dan kromosom diskret) dan variasi dalam dominasi alel. Yang terakhir memungkinkan pemodelan alel-alel merugikan resesif parsial, yang merupakan kontributor penting bagi beban genetik (Charlesworth dan Willis 2009 ; Hedrick dan Garcia-Dorado 2016 ). Interaksi epistatik (ketika ekspresi gen dimodifikasi oleh ekspresi ≥ 1 gen lainnya) juga dapat dimodelkan dalam SLiM dan simulasi menawarkan alat yang ampuh untuk memahami pengaruh epistasis pada akumulasi, ekspresi, dan pembersihan mutasi yang merugikan dalam populasi kecil. Namun, peran epistasis dalam kepunahan dan penyelamatan genetik populasi kecil berada di luar cakupan makalah saat ini. Versi terbaru SLiM (saat ini V4.3) melonggarkan asumsi model Wright-Fisher klasik (lihat Glosarium) genetika populasi dan dapat mensimulasikan berbagai proses yang realistis secara ekologis, termasuk generasi yang tumpang tindih, struktur usia, umpan balik kepadatan pada pertumbuhan populasi, variasi dalam kesuburan dan kelangsungan hidup individu, serta interaksi spesies yang berguna untuk memodelkan dinamika predator-mangsa atau penyakit menular (Haller dan Messer 2019 , 2023 ). Model SLiM dapat eksplisit secara spasial, dengan individu yang dimodelkan dalam ruang kontinu (hingga 3 dimensi), dan lapisan variabel lingkungan yang terkotak-kotak.
quantiNemo (Neuenschwander et al. 2019 ) adalah perluasan dari program Nemo asli (Guillaume dan Rougemont 2006 ; nemo2.sourceforge.io ), yang juga telah diperluas sebagai Nemo-age (Cotto et al. 2020 ). Di sini, kami fokus pada quantiNemo karena telah memperluas kapasitas yang relevan untuk simulasi penyelamatan genetik. quantiNemo adalah program simulasi stokastik yang eksplisit secara genetik dan berbasis individu. Program ini dikembangkan untuk menyelidiki efek seleksi, mutasi, rekombinasi, dan pergeseran pada sifat kuantitatif dengan berbagai arsitektur dalam populasi terstruktur yang terhubung oleh migrasi dan berlokasi di habitat heterogen. quantiNemo fleksibel dalam beberapa komponen: populasi, seleksi, arsitektur sifat, peta genetik untuk lokus dan/atau penanda sifat kuantitatif, lingkungan, demografi, sistem perkawinan, dll. Program ini adalah program konsol berorientasi objek yang dikodekan dalam C++, berjalan pada platform komputer apa pun, dan didistribusikan di bawah lisensi sumber terbuka. Berbeda dengan SLiM , yang memodelkan mutasi pada kromosom virtual, quantiNemo mengimplementasikan peta rekombinasi tempat lokus yang mengkode berbagai jenis sifat dapat ditempatkan bersama-sama. Proses genetik (dan evolusi sifat kuantitatif) yang dimodelkan dalam quantiNemo yang relevan dengan simulasi penyelamatan genetik mencakup mutasi yang merusak, penanda netral (mikrosatelit, polimorfisme nukleotida tunggal), lokus sifat kuantitatif (QTL), tingkat penyebaran khusus jenis kelamin, dan sifat riwayat hidup, antara lain. Kerangka kerja ini mendukung struktur spasial, serta berbagai arsitektur genetik yang berguna untuk mensimulasikan pergeseran genetik, aliran gen, dan seleksi. Tidak seperti SLiM dan CDMetaPop (dijelaskan di bawah), quantiNemo hanya dapat memodelkan spesies tunggal, tetapi jumlah lokus, individu, atau populasi yang disimulasikan hanya dibatasi oleh daya komputasi. Kami tidak mengetahui adanya penelitian yang membandingkan efisiensi komputasi model quantiNemo dengan kompleksitas yang bervariasi. Oleh karena itu, kami tidak dapat memberikan indikasi batas atas yang realistis untuk parameter ini, yang perlu dioptimalkan berdasarkan kasus per kasus.
CDMetaPOP (‘ Cost- Distance Meta- POPulation ‘ ; github.com/ComputationalEcologyLab/CDMetaPOP ) (Day et al. 2023 ; Landguth et al. 2017 ) adalah program simulasi yang digunakan untuk memprediksi pengaruh struktur lanskap dan pergerakan, perkembangbiakan, dan penyebaran berbasis individu pada kemunculan pola spasial dalam data genetik populasi (yaitu, genetika lanskap). Seperti quantiNemo , CDMetaPop memperlakukan lanskap sebagai kisi petak-petak di mana individu berbagi lingkungan yang sama, dengan opsi untuk menentukan petak-petak pada skala individu. Menentukan variabel lingkungan antar-petak menciptakan lanskap heterogen, dengan pergerakan individu melintasi lanskap ini ditentukan oleh permukaan resistensi (atau permeabilitas). Di dalam petak-petak, individu tumbuh, bereproduksi, bermigrasi, dan mati. Proses ekologi ini menimbulkan perubahan netral atau adaptif yang eksplisit secara spasial dalam frekuensi alel pada jumlah lokus dan alel yang tidak terbatas. Tidak seperti SLiM (genotipe sebagai genom virtual) dan quantiNemo (genotipe sebagai peta rekombinasi), genotipe individual dalam CDMetaPop terdiri dari daftar lokus dan alel yang ditentukan pengguna. Model diinisialisasi dengan frekuensi alel tingkat populasi (baik nilai hipotetis maupun estimasi empiris) dan keturunan mewarisi genotipe berdasarkan pewarisan Mendel. Pengguna dapat memilih dari sejumlah model mutasi (misalnya, k -alel, bertahap). Polimorfisme nukleotida tunggal dapat diakomodasi dalam model sebagai lokus bi-alel. Tingkat demografi (misalnya, kelahiran, kematian, penyebaran) ditentukan sebagai probabilitas, dengan hasil individual dialokasikan secara binomial, yang berarti bahwa stokastisitas demografi tersirat. Variabel lingkungan dapat bervariasi baik di ruang maupun waktu, dan menentukan rata-rata dan varians menggabungkan stokastisitas lingkungan. Seperti SLiM , inkarnasi terbaru ( CDMetaPop V2) memperluas kapasitas untuk memodelkan dinamika eko-evolusi dalam komunitas multi-spesies. Jumlah lokus, alel, individu, dan spesies yang dapat dimodelkan hanya dibatasi oleh daya komputasi, meskipun direkomendasikan bahwa lokus × alel < 10 5 (Day et al. 2023 ; Landguth et al. 2017 ). Kami tidak mengetahui adanya penelitian yang membandingkan efisiensi komputasi model CDMetaPop dengan kompleksitas yang bervariasi. Oleh karena itu, kami tidak dapat memberikan indikasi batas atas yang realistis untuk semua kombinasi parameter, yang perlu dioptimalkan berdasarkan kasus per kasus.
RangeShifter ( rangeshifter.github.io ) (Bocedi et al. 2021 ) adalah alat pemodelan yang dirancang untuk mensimulasikan dinamika ekologi dan evolusi spesies individu dalam menanggapi pergeseran lingkungan. Platform ini menggabungkan bentang alam yang bervariasi secara temporal, pemodelan genetik eksplisit, dan evolusi mekanisme penyebaran dengan fokus khusus pada identifikasi risiko populasi dan pemahaman proses yang mendorong dinamika jangkauan spesies, termasuk dampak struktur bentang alam pada keragaman genetik dan adaptasi. Aspek genetik RangeShifter menawarkan pemodelan terperinci tentang sifat-sifat yang dapat diwariskan, mendukung evolusi strategi penyebaran dan menyediakan dasar untuk studi lanjutan tentang genetika bentang alam. Sementara RangeShifter menawarkan wawasan yang kuat tentang respons spesies terhadap perubahan lingkungan, utilitas aspek genetik diredam oleh tidak adanya fungsi tertentu, seperti representasi bernuansa proses genetik yang ditemukan dalam perangkat lunak lain seperti SLiM, quantiNemo , dan CDMetaPop . RangeShifter juga tidak memiliki kapasitas untuk memodelkan interaksi antarspesies seperti SLiM dan CDMetaPop . Karena RangeShifter menyediakan kerangka kerja baru untuk pemodelan penyebaran dan menghubungkan ekologi pergerakan dengan dinamika spasial, ini masih merupakan alat yang berpotensi berguna untuk simulasi penyelamatan genetik.
HexSim ( hexsim.net ) (Schumaker dan Brookes 2018 ) adalah model berbasis individu yang eksplisit secara spasial yang dirancang untuk mempelajari dinamika populasi satwa liar di berbagai lanskap dan skenario. Model ini berfokus pada kompleksitas spasial yang memengaruhi perilaku individu seperti pergerakan, kelangsungan hidup, reproduksi, dan penyebaran, sehingga memengaruhi dinamika dan distribusi populasi secara keseluruhan. HexSim tidak hanya dapat mensimulasikan dinamika populasi yang kompleks, termasuk fluktuasi ukuran populasi, struktur usia, dan dampak gangguan lingkungan, tetapi juga menggabungkan faktor genetik untuk memeriksa aliran gen, keragaman genetik, dan dampak lanskap pada struktur genetik. Komponen genetik ini dapat mencakup pelacakan alel atau genotipe dari waktu ke waktu, menilai dampak pergeseran dan seleksi, dan mengevaluasi dampak konektivitas lanskap pada keragaman genetik. Model ini menghubungkan sifat riwayat hidup dengan komponen demografi dan genetik, yang memungkinkan simulasi tekanan selektif pada sifat genetik dan konsekuensi demografinya. HexSim mengintegrasikan proses demografi dengan fitur lanskap spasial untuk menawarkan wawasan tentang kelangsungan hidup populasi, kesehatan genetik, dan persyaratan konservasi.
Kami mengakui bahwa ada program perangkat lunak berbasis individu dan eksplisit secara genetis yang mungkin sama-sama berlaku untuk pemodelan penyelamatan genetik. Misalnya, SimAdapt (Rebaudo et al. 2013 ) berdasarkan lingkungan pemodelan NetLogo (Wilensky 1999 ) dan Geonomics (Hart et al. 2021 ) adalah dua contoh, dan mungkin ada yang lain. Pengguna mungkin berbeda dalam elemen program mana yang mereka anggap lebih berguna dalam setiap konteks. Di luar cakupan makalah kami untuk memberikan tinjauan menyeluruh tentang kemampuan semua program yang tersedia. Sebaliknya, fokus kami di sini adalah untuk membandingkan kemampuan dari serangkaian lima program representatif untuk mendukung pesan utama kami—bahwa implementasi penyelamatan genetik dalam konservasi harus dipandu oleh model yang mereplikasi penguatan timbal balik dari stokastisitas demografis, pergeseran genetik, dan perkawinan sedarah.
4 Contoh Model Demo-Genetik untuk Simulasi Penyelamatan Genetik
4.1 Pengembangan Model
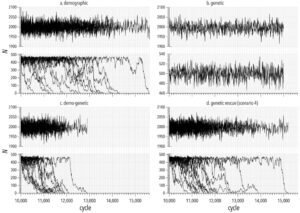
Di sini, kami mengembangkan simulasi model heuristik untuk menunjukkan pengaruh umpan balik demo-genetik pada dinamika kepunahan dan hasil penyelamatan genetik populasi kecil. Kami memodelkan populasi hipotetis yang telah mengalami penurunan tiba-tiba dalam ukuran populasi. Riwayat hidup spesies hipotetis kami secara longgar terinspirasi oleh koala ( Phascolarctos cinereus ), tetapi parameter spesifik yang kami gunakan tidak diinformasikan secara empiris atau dikalibrasi karena tujuan kami adalah untuk menunjukkan satu contoh tentang cara membangun model di mana umpan balik demo-genetik muncul dari mekanisme tingkat individu dan genetik yang mendasarinya (Gambar 2 ) dan memengaruhi hasil penyelamatan genetik virtual. Kami mulai dengan mengembangkan satu set tiga model dasar di mana dinamika populasi yang disimulasikan muncul dari stokastisitas demografis saja (model demografis), pergeseran genetik dan perkawinan sedarah saja (model genetik), dan penguatan bersama dari stokastisitas demografis, pergeseran genetik, dan perkawinan sedarah (model demo-genetik) (Gambar 3 ; lihat Informasi Pendukung untuk deskripsi terperinci tentang bagaimana kami membangun model).
Kami menggunakan SLiM ( v 4.0; Haller dan Messer 2023 ) untuk membangun model. Untuk masing-masing dari tiga model, kami mensimulasikan dua populasi hipotetis—satu yang mempertahankan ukuran populasi ekuilibrium pada daya dukung ( K1 ) 2000 individu (P1 = populasi kontrol) untuk keseluruhan simulasi, dan yang lain yang mengalami penurunan demografis mendadak (misalnya, karena hilangnya habitat) ke daya dukung ( K2 ) 500 individu (P2 ; populasi fokus). Penurunan mendadak terjadi setelah periode burn-in 10.000 siklus (direplikasi dalam setiap iterasi), yang kira-kira setara dengan tahun dalam model (lihat Resolusi temporal siklus dan iterasi simulasi ). Setelah penurunan mendadak dalam kelimpahan pada populasi fokus, kami menjalankan setiap iterasi simulasi hingga populasi tersebut punah. Untuk setiap iterasi, kami mencatat waktu kepunahan T ext yang diukur dalam siklus simulasi (≈ tahun), yang darinya kami hitung mean T ext di semua iterasi untuk model tertentu. Untuk setiap model, kami juga menghitung probabilitas kepunahan Pr ( ext ) yang dihitung sebagai 1 dikurangi fraksi iterasi di mana populasi fokus tetap ada setelah waktu acak 1500 siklus (≈ tahun) setelah populasi menurun. Kami menggunakan T ext dan Pr ( ext ) untuk membandingkan pengaruh stokastisitas demografi, pergeseran dan perkawinan sedarah, dan umpan balik demo-genetik, masing-masing, pada kelangsungan hidup populasi.
Berikutnya, kami menggunakan model demo-genetik dasar untuk memeriksa efek dari berbagai skenario penyelamatan genetik pada T ext dan Pr ( ext ). Kami menekankan bahwa latihan ini dimaksudkan sebagai bukti konsep dan bukan untuk memandu keputusan manajemen. Kami menggunakan model demo-genetik untuk memeriksa hasil dari empat skenario penyelamatan genetik utama pada T ext dan Pr ( ext ). Dalam setiap skenario, kami memvariasikan: ( i ) jumlah individu yang berpindah dari populasi kontrol (P 1 dengan K 1 = 2000) ke dalam populasi fokus (P 2 dengan K 2 = 500) dalam setiap peristiwa ‘translokasi’ tunggal (ukuran kohort), dan ( ii ) jumlah peristiwa translokasi. Keempat skenario tersebut adalah: Skenario 1 : 50 individu pindah sekali pada 250 tahun setelah penurunan demografi; Skenario 2 : 100 individu pindah sekali pada 250 tahun setelah penurunan demografi; Skenario 3 : 50 orang pindah tiga kali pada 250, 255, dan 260 tahun setelah penurunan demografi; dan Skenario 4 : 100 orang pindah tiga kali pada 250, 255, dan 260 tahun setelah penurunan demografi.
Dalam keempat skenario, kami hanya mempertimbangkan efek ukuran kelompok dan jumlah kejadian translokasi. Ada sejumlah variabel lain yang dapat dipertimbangkan oleh para pengambil keputusan saat menerapkan penyelamatan genetik pada populasi alami. Di bawah ini, dalam Menggunakan simulasi untuk menginformasikan penyelamatan genetik terapan , kami memberikan pembahasan terperinci tentang bagaimana simulasi dapat digunakan untuk membandingkan dan memberi peringkat skenario penyelamatan genetik yang berbeda dalam berbagai variabel. Di sini, cukup untuk fokus pada faktor-faktor yang umumnya bervariasi di antara implementasi penyelamatan genetik. Semua kode SLiM yang diperlukan untuk menjalankan simulasi tersedia di https://zenodo.org/doi/10.5281/zenodo.10939288 .
4.2 Hasil Simulasi Model Heuristik
Dalam model demografi, varians dalam probabilitas survival menyebabkan kelimpahan berfluktuasi secara acak dari waktu ke waktu (Gambar 3 ) dan memberikan probabilitas kepunahan Pr ( ext ) = 0,24 untuk populasi fokus (Gambar 4 ). Dalam model genetik, mutasi merusak yang sebagian resesif (yaitu, alel dengan dominasi tidak lengkap; Glosarium, Tabel 1 ) terakumulasi (pada frekuensi rendah) di populasi fokus dan kontrol, tetapi pengurangan kelimpahan saja tidak cukup untuk menyebabkan perkawinan sedarah dan pergeseran genetik untuk mengalahkan dinamika kompensasi (yaitu, pertumbuhan populasi menuju daya dukung, K = 500) untuk membuka kedok beban genetik dan mengurangi kebugaran rata-rata dan laju pertumbuhan populasi (Gambar 3 ). Dengan kata lain, tanpa stokastisitas dalam survival, mutasi merusak dibersihkan melalui seleksi yang bekerja pada individu yang homozigot untuk mutasi merusak (depresi perkawinan sedarah). Oleh karena itu, tanpa efek tambahan stokastisitas demografi (dan dengan K = 500), tidak ada kepunahan dalam simulasi model genetik (Gambar 4 ).
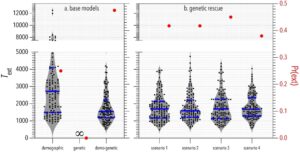
Dalam model demo-genetik, stokastisitas demografi berarti bahwa pengurangan periodik dalam ukuran populasi < K (yaitu, kemacetan) mengungkap mutasi yang merusak melalui peningkatan perkawinan sedarah dan pergeseran genetik. Umpan balik demo-genetik mengurangi kebugaran individu rata-rata dan laju pertumbuhan populasi (Gambar 3 ) dan meningkatkan probabilitas kepunahan sebesar 0,24 dan 0,48 dibandingkan dengan yang diamati dalam model demografi dan genetik, masing-masing (Gambar 4 ). Pilihan model dasar juga memengaruhi T ext dalam populasi yang disimulasikan (Gambar 4 ). Rata-rata, kepunahan terjadi 1444 siklus (≈ tahun) lebih cepat dalam simulasi model dasar demo-genetik dibandingkan dengan model dasar demografi (Gambar 4 ). Kami tidak membandingkan T ext antara model dasar demo-genetik dan genetik karena kami secara sewenang-wenang membatalkan simulasi model dasar genetik pada 5000 siklus setelah penurunan demografi (di mana semua iterasi model dasar genetik tetap ada).
Kami kemudian menghitung waktu menuju kepunahan dan probabilitas kepunahan dalam simulasi empat skenario penyelamatan genetik dan membandingkannya dengan nilai yang diperoleh dari model dasar demo-genetik. Waktu menuju kepunahan tidak berbeda di antara skenario penyelamatan genetik dan model dasar demo-genetik. Namun, penyelamatan genetik yang disimulasikan sedikit mengurangi probabilitas kepunahan sebesar 3%–9% relatif terhadap model dasar demo-genetik, dengan skenario 4 memiliki dampak terbesar (Gambar 4 ).
4.3 Resolusi Temporal Siklus Simulasi dan Iterasi
Dalam model demonstrasi ini, ‘siklus’ simulasi mendekati tahun, tetapi ini tidak harus menjadi kasusnya. Durasi yang diwakili oleh satu siklus akan bergantung pada sistem yang dimodelkan dan tujuan studi; suatu siklus dapat mewakili periode < 1 tahun jika, misalnya, variasi musiman dalam mortalitas diduga memengaruhi struktur demografis suatu populasi. Siklus juga dapat > 1 tahun ketika, misalnya, spesies menunjukkan variasi tahunan yang rendah dalam mortalitas dan berkembang biak secara episodik (misalnya, terkait dengan peristiwa lingkungan multi-tahunan). Kerangka waktu di mana hasil dari setiap iterasi simulasi terjadi (yaitu, waktu hingga kepunahan T ext dan probabilitas kepunahan pada waktu t di masa depan Pr ( ext ); Gambar 3 dan Gambar 4 ) sangat bergantung pada kombinasi spesifik nilai parameter yang digunakan dalam model tertentu. Tujuan dari latihan pemodelan ini bukanlah untuk memprediksi hasil spesifik dari penyelamatan genetik dalam hal T ext dan Pr ( ext ), melainkan untuk membandingkan dan memberi peringkat berbagai skenario penyelamatan genetik berdasarkan pengaruh relatifnya pada T ext dan Pr ( ext ) dalam populasi virtual (lihat Menggunakan simulasi untuk menginformasikan penyelamatan genetik terapan ).
Di bawah ini, kami membahas bagaimana model demo-genetik dapat dikembangkan dan diterapkan menggunakan data genetik yang tersedia untuk populasi spesies terancam yang menjadi sasaran penyelamatan genetik. Kami menggunakan studi kasus marsupial asli Australia yang terancam untuk menunjukkan jenis data genetik yang tersedia dan menyarankan serangkaian strategi pemodelan dan bagaimana data dapat digunakan untuk mengembangkan dan menerapkan model.
5 Marsupial Australia yang Terancam Punah: Studi Kasus dan Panduan untuk Mengukur Model Menggunakan Data Genetik
5.1 Ketersediaan Data Genetik untuk Marsupial Australia yang Terancam
Australia adalah salah satu negara dengan keanekaragaman hayati yang tinggi yang dicirikan oleh endemismenya yang tinggi (misalnya, 87% fauna mamalianya endemik) (Woinarski et al. 2015 ). Australia juga memiliki catatan kepunahan mamalia terburuk di antara negara mana pun—setidaknya 17 dari ~179 spesies marsupial asli telah punah dalam 200 tahun terakhir (Legge, Hayward et al. 2023a ; Woinarski et al. 2015 ). Sebaliknya, tidak satu pun dari ~130 spesies marsupial Dunia Baru telah punah selama periode yang sama (Martin et al. 2022 ). Kepunahan marsupial terutama disebabkan oleh tekanan predator baru dari kucing ( Felis catus silvestris ) dan rubah merah Eropa ( Vulpes vulpes ), serta dari hilangnya habitat, perubahan rezim kebakaran, dan perubahan iklim, yang semuanya saling berinteraksi (Legge, Rumpff et al. 2023b ; Legge, Hayward et al. 2023a ). Saat ini, ada tambahan 110 marsupial Australia (~65% dari spesies yang masih ada) yang terdaftar sebagai terancam berdasarkan Undang-Undang Perlindungan Lingkungan dan Konservasi Keanekaragaman Hayati Australia 1999 (Undang-Undang EPBC; jumlah total terdiri dari 55 spesies, subspesies, atau populasi yang didefinisikan secara geografis yang Rentan, 46 Terancam Punah, dan 9 Sangat Terancam Punah). Banyak dari spesies ini sekarang hanya terdapat dalam populasi kecil (< 1000 individu) dan terisolasi yang menempati < 10% dari rentang geografis sebelumnya (Legge, Hayward et al. 2023a ; Woinarski dan Fisher 2022 ). Efek gabungan dari perkawinan sedarah dan pergeseran genetik sekarang diakui sebagai ancaman tambahan terhadap kelangsungan hidup banyak spesies marsupial yang terancam, menjadikan prospek penyelamatan genetik sebagai pertimbangan manajemen yang serius (Cowen et al. 2023 ; Farquharson et al. 2021 ; Hoffmann et al. 2021 ; Nilsson et al. 2023 ; Nistelberger et al. 2023 ).
Kemajuan pesat dalam teknologi sekuensing dan bioinformatika telah meningkatkan kelayakan untuk mendapatkan data genomik beresolusi tinggi (dan berbiaya rendah) untuk spesies yang terancam (Bernatchez et al. 2024 ; Hogg et al. 2022 ). Data genomik ini, serta banyak penelitian berdasarkan DNA mikrosatelit (misalnya, DeWoody et al. 2021 ) dapat dimanfaatkan untuk mengembangkan dan menerapkan model simulasi penyelamatan genetik. Di bagian ini, kami fokus pada marsupial Australia yang terancam untuk menyarankan pendekatan berdasarkan data yang tersedia untuk umum untuk membuat parameter, mengkalibrasi, dan memvalidasi model demo-genetik. Tujuannya adalah untuk memandu pengambilan keputusan dalam implementasi penyelamatan genetik (lih. memandu pengambilan keputusan tentang perlunya penyelamatan genetik). Kami berfokus pada spesies marsupial yang terdaftar sebagai Terancam Punah dan Sangat Terancam Punah berdasarkan Undang-Undang Federal (EPBC) 1999 sebagai contoh pengambilan keputusan berbasis model dalam penyelamatan genetik (dijelaskan di bawah ini dalam Menggunakan simulasi untuk menginformasikan penyelamatan genetik terapan ). Pendekatan kami dapat diterapkan pada spesies atau populasi apa pun tanpa memandang status ancaman.
Bahasa Indonesia: Kami menelusuri Web of Science pada 7 September 2023 untuk publikasi yang telah ditinjau sejawat yang menyediakan estimasi ukuran keragaman genetik/genomik populasi dan perkawinan sedarah pada marsupial menggunakan empat istilah penelusuran berikut: ( i ) marsupial* AND genetic* AND microsatellite*; ( ii ) marsupial* AND genomic* AND snp; ( iii ) marsupial* AND genomic* OR genetic* AND bottleneck; dan ( iv ) marsupial* AND genomic* AND divers* OR demog*. Penelusuran kami menghasilkan 173 hasil, yang 135 di antaranya unik. Kami kemudian mengelompokkan daftar tersebut ke artikel utama (yaitu, bukan artikel tinjauan) yang menyertakan nama-nama marsupial asli Australia yang terdaftar sebagai Terancam Punah atau Sangat Terancam Punah berdasarkan Undang-Undang EPBC (Tabel S1 ). Berikutnya, kami menghapus makalah yang membahas tentang pengembangan sumber daya molekuler (misalnya, pengembangan mikrosatelit, genom referensi) dan tidak menyertakan perkiraan keragaman genetik populasi, struktur, atau perkawinan sedarah.
Prosedur yang dijelaskan di atas menghasilkan sampel sebanyak 73 artikel yang menyediakan data genetik untuk 21 spesies. Kami kemudian mengulangi prosedur yang sama pada tanggal 19 November 2024 untuk memperbarui hasil kami agar mencakup artikel yang baru-baru ini diterbitkan. Kami juga menyelesaikan beberapa pencarian oportunistik tambahan untuk menemukan artikel yang kami tahu telah terlewatkan dalam pencarian awal kami. Kami menerapkan prosedur pemangkasan yang sama seperti yang dijelaskan di atas pada daftar makalah yang dihasilkan dari pencarian yang diperbarui. Sampel akhir kami yang dihasilkan dari semua pencarian mencakup 79 makalah untuk 21 spesies (Tabel 4 ).
| Jenis | Tipe data | Lokasi | Analisis | Hilangnya keberagaman | Referensi |
|---|---|---|---|---|---|
| Bettongia penicillata | Mikrosatelit, mtDNA, DNA purba, SNP | 4+ pop, 9 lokasi | Keanekaragaman, aliran gen, struktur populasi, filogeni, keanekaragaman purba vs. modern, pemodelan kelangsungan hidup populasi (berbasis mikrosatelit), desain penangkaran, perkawinan sedarah (berbasis SNP) | Ya; perbandingan DNA purba | Farquharson dkk. 2021 ; Pacioni dkk. 2011 , 2013 , 2015 , 2018 , 2020 |
| Bettongia tropis | Mikrosatelit | 1 buah | Sistem perkawinan; keterkaitan dan penyebaran individu | Tidak dilaporkan secara langsung | Paus dkk. 2012 |
| Burung pipit Burramys parvus | Mikrosatelit, mtDNA | 13 lokasi, ~ 4 populasi | Struktur populasi, keanekaragaman, N e , kemacetan, filogeni | Tanda-tanda kemacetan genetik | Mitrovski dkk. 2007 ; Minggu dkk. 2016 |
| Dasyurus halusatus | Mikrosatelit, SNP, mtDNA | 10+ lokasi | Struktur pop, keragaman, N e (perbandingan temporal), seleksi | Ya, dalam translokasi pulau vs daratan , dan dalam sejarah vs kontemporer | Cardoso dkk. 2009 ; Hohnen dkk. 2016 ; Bagaimana dkk. 2009 ; Spencer dkk. 2017 ; von Takach dkk. 2022 , 2024 ; Minggu dkk. 2016 |
| Dasyurus maculatus | Mikrosatelit | 12+ lokasi | Struktur pop, keberagaman, filogeografi historis | Mungkin | Firestone dkk. 1999 ; Ruibal dkk. 2009 , 2010 ; Minggu dkk. 2016 |
| Dasyurus viverrinus | Mikrosatelit | 9 daerah | Struktur pop, keberagaman, hambatan, N e | Tanda-tanda kemacetan genetik | Cardoso dkk. 2014 |
| Gymnobelideus leucocephalus | Mikrosatelit, mtDNA, SNP | 4+ populer | Struktur populasi, keanekaragaman (termasuk tren temporal), N e , perkawinan sedarah, depresi perkawinan sedarah (pewarisan sifat kebugaran), simulasi kelangsungan hidup populasi/rekomendasi penyelamatan genetik | Ya; misalnya, penurunan temporal dalam heterozigositas | (Hansen dkk. 2009 ; Hansen dan Taylor 2008 ; Zilko dkk. 2020 , 2021 |
| Isoodon obesulus | Mikrosatelit | 10+ lokasi | Struktur populasi, keanekaragaman, aliran gen | Tidak secara langsung, namun struktur pop diduga terpengaruh oleh fragmentasi | Li dkk. 2013 , 2014 , 2015 , 2016 ; Ramalho dkk. 2018 ; Zenger dkk. 2005 ; Zenger dan Johnston 2001 |
| Lagorchestes hirsutus | Mikrosatelit, mtDNA | 3 muncul | Struktur populasi, keanekaragaman, filogeni | Tidak dilaporkan secara langsung | Eldridge dkk. 2004 ; Eldridge dan Potter 2019 |
| Lasiorhinus krefftii | Mikrosatelit | 1 buah | keterkaitan dan penyebaran individu | Tidak dilaporkan secara langsung | Taylor dan kawan-kawan 1997 |
| Myrmecobius fasciatus | Mikrosatelit | 1 buah | Keturunan, keberagaman | Tidak dilaporkan secara langsung | Spencer dan kawan-kawan 2020 |
| Onikogalea fraenata | Mikrosatelit | Sisa pop, hasil penangkaran, kelompok yang dipindahkan | Struktur populasi, keanekaragaman, keturunan, keberhasilan reproduksi | Keanekaragaman yang berkurang setelah translokasi | Sigg 2006 ; Sigg dkk. 2005 |
| Paratechinus apikal | Mikrosatelit, mtDNA | Daratan +3 pulau pop | Struktur populasi, keanekaragaman, hambatan, filogeografi, analisis kelangsungan hidup populasi untuk memandu translokasi | Tanda-tanda kemacetan dan penurunan N e di pulau-pulau | Aisya dkk. 2022 ; Pabrik dkk. 2004 ; Thavornkanlapachai dkk. 2019 , 2021 |
| Peramal
gunnii |
Mikrosatelit, mtDNA | 7+ | Pop. struktur, keanekaragaman, aliran gen | Keragaman yang lebih rendah pada sampel modern dibandingkan dengan sampel historis | Hitam dkk. 2024 ; Minggu dkk. 2013 |
| Bunga bugenvil Perameles | Mikrosatelit, mtDNA | 5 pulau (baik pulau alami + pulau pop pendatang) | Struktur pop, keragaman, uji kemacetan | Tidak baru-baru ini; keragaman rendah jangka panjang diikuti oleh pencampuran dalam penangkaran = keragaman lebih tinggi | Smith dan Hughes 2008 ; White dkk. 2018 |
| Burung pipit volans | Mikrosatelit | 11 situs | Struktur pop dan konektivitas, asal usul | bukti lemah | Taylor dan kawan-kawan 2007 |
| Petrogale lateralis | Mikrosatelit, mtDNA | 4+ liar dan berbagai penangkaran | Struktur dan penyebaran populasi, keanekaragaman, garis keturunan, taksonomi, analisis kelangsungan hidup populasi | Berbagai temuan, namun bukti adanya kemacetan daratan baru-baru ini dan rendahnya keragaman jangka panjang di pulau-pulau | Eldridge dkk. 1999 , 2004 ; Eldridge dan Potter 2019 ; Nilsson dkk. 2023 ; Potter dkk. 2017 ; Ruykys dan Lancaster 2015 ; West dkk. 2018 |
| Burung pipit Phascolarctos cinereus | Mikrosatelit, mtDNA, SNP, MHC, WGS, transkriptom | 91+ lokasi, 5 klaster utama | Banyak dan beragam | Tanda-tanda kemacetan genetik | Cheng dkk. 2018 ; Cristescu dkk. 2010 ; Hobbs dkk. 2014 ; Johnson dkk. 2018 ; Kjeldsen dkk. 2019 ; Lott dkk. 2022 , 2024 |
| Bunga gilbertii yang potorous | Mikrosatelit, mtDNA | 1 pop yang masih ada, ditambah tawanan | Struktur pop, keragaman, uji kemacetan, N e | Ya; tanda-tanda kemacetan genetik | Sinclair dkk. 2002 |
| Sarcophilus harrisii | Mikrosatelit, mtDNA, SNP, MHC, WGS, transkriptom | 10+ lokasi | Banyak dan beragam, termasuk demografi penduduk | Ya; tanda genomik dari kemacetan yang berulang | Brandi dkk. 2019 ; Cheng dkk. 2012 ; Cheng dan Belov 2012 ; Cui dkk. 2015 ; Deakin dan Belov 2012 ; Farquharson dkk. 2022 ; Grueber dkk. 2021 ; Jones dkk. 2003 , 2004 ; Miller dkk. 2011 ; Murchison dkk. 2012 ; Perak dkk. 2021 ; Stahlke dkk. 2021 ; Ujvari dkk. 2014 ; Hutan dkk. 2018 ; Wright dkk. 2019 |
| Sminthopsis psammophila | Mikrosatelit, mtDNA | 11+ Situs, ~ 3 pop. | Struktur pop, keberagaman, uji kemacetan, N e , keterkaitan | Tanda-tanda kemacetan genetik | McLean dkk. 2014 , 2018 |
Singkatan: het, heterozigot; MHC, kompleks histokompatibilitas utama; mtDNA, DNA mitokondria; N c , ukuran populasi sensus; N e , ukuran populasi efektif; pop, populasi; SNP, polimorfisme nukleotida tunggal; WGS, pengurutan genom keseluruhan.
Jenis data genetik yang tersedia untuk spesies yang menjadi fokus kami berkisar dari penanda mikrosatelit hingga polimorfisme nukleotida tunggal di seluruh genom yang diperoleh dari sekuensing representasi tereduksi (RRS; misalnya, DArTseq, ddRAD) hingga genom utuh. Dari 21 spesies yang data genetiknya kami temukan, semuanya memiliki data mikrosatelit, 13 spesies memiliki DNA mitokondria (mtDNA), sembilan spesies memiliki SNP di seluruh genom, dan dua spesies memiliki sekuens genom utuh— Phascolarctos cinereus (koala) dan Sarcophilus harrisii (setan Tasmania). Ada juga satu spesies yang DNA purbanya telah disekuensing— Bettonia penicillata (woylie). Sebagian besar spesies (15) direpresentasikan oleh satu atau dua jenis data sekuens, sementara lebih sedikit (6) yang memiliki tiga atau lebih jenis data. Spesies dengan data genetik terbanyak termasuk Bettongia penicillata , Dasyurus hallucatus (quoll utara), Perameles bougainville (bandicoot berpalang barat), Gymnobelideus leadbeateri (possum Leadbeater), Phascolarctos cinereus , dan Sarcophilus harrisii .
5.2 Penggunaan Data dalam Pengembangan dan Aplikasi Model
Di sini kami menghubungkan tinjauan pustaka data genetik pada marsupial dengan bagian pemodelan di atas dengan membahas strategi potensial untuk simulasi penyelamatan genetik. Data genetik dapat digunakan pada berbagai tahap pengembangan model dan dapat diterapkan pada satu atau semua tahap parameterisasi, kalibrasi, dan/atau validasi. Mengingat fleksibilitas platform pemodelan berbasis individu dan berbagai macam tujuan dan tipe data spesifik konteks, kami tidak dapat memberikan preskriptif tentang strategi untuk penggunaan data dalam pengembangan dan aplikasi model. Sebaliknya, kami memberikan beberapa contoh bagaimana data dapat dikombinasikan dengan model berbasis individu yang eksplisit secara genetik untuk mensimulasikan penyelamatan genetik. Sensitivitas hasil model terhadap ketidakpastian parameter dapat diukur menggunakan analisis sensitivitas global (Prowse et al. 2016 ) yang memiliki manfaat ganda untuk mengukur ketidakpastian dalam hasil model serta memprioritaskan data mana yang penting untuk prediksi model yang realistis. Evaluasi kuantitatif nilai parameter dan pengaruhnya terhadap hasil simulasi harus menghindari penyalahgunaan estimasi kesalahan Neyman-Pearson Tipe I ( nilai p ) dan sebaliknya berfokus pada ukuran efek (lihat White et al. 2018 untuk penjelasan mengapa uji hipotesis statistik frekuentis tidak tepat untuk menafsirkan hasil model simulasi).
5.2.1 Strategi 1: Simulasi Penyelamatan Genetik Menggunakan Variasi Urutan Virtual
Strategi ini melibatkan simulasi mutasi de novo yang muncul dan berubah dalam frekuensi selama periode burn-in, diikuti oleh penurunan populasi sebelum simulasi penyelamatan genetik. Di sini, nilai parameter perlu didefinisikan untuk mekanisme yang menghasilkan variasi urutan virtual, termasuk laju mutasi, laju rekombinasi, serta efek kebugaran dan dominasi mutasi de novo virtual . Karena populasi yang membutuhkan penyelamatan genetik tidak mungkin memiliki data yang diperlukan untuk memperkirakan parameter mekanistik ini, estimasi dapat digunakan dari (dalam urutan preferensi) populasi lain dari spesies yang sama, spesies terkait, atau spesies yang tidak terkait untuk mengisi celah tersebut. Misalnya, laju mutasi untuk koala baru-baru ini diperkirakan (T. Kovacs et al., dalam persiapan), yang seharusnya menggantikan estimasi untuk Drosophila atau manusia jika simulasi penyelamatan genetik populasi marsupial. Demikian pula, pada subspesies tikus ( Mus musculus castaneus ), distribusi efek kebugaran dari mutasi nonsinonim bersifat bimodal: sebagian besar mutasi hampir netral, dan beberapa mutasi sangat merusak (Kousathanas dan Keightley 2013 ). Parameter yang tidak ada datanya dapat diperkirakan dari hubungan allometrik atau berdasarkan asumsi wajar lainnya yang diinformasikan oleh prediksi teoritis (misalnya, model geometri Fisher; Orr 2006 ). Seperti yang dinyatakan di atas, interpretasi yang cermat dari keluaran model harus dipandu oleh analisis sensitivitas global untuk mengukur dan menyoroti ketidakpastian.
Data yang lebih umum tersedia untuk populasi target mencakup estimasi berbasis urutan keragaman genetik dan perkawinan sedarah (misalnya, kekayaan alel, heterozigositas/homozigositas, perkawinan sedarah atau koefisien terkait), dan yang lebih jarang, beban genetik. Informasi tersebut dapat digunakan untuk mengkalibrasi (dan idealnya, memvalidasi) parameter mekanistik dalam model yang menimbulkan variasi urutan virtual. Misalnya, tingkat mutasi dan rekombinasi, efek kebugaran, dan dominasi dapat dikalibrasi sehingga estimasi parameter genetik dalam populasi yang disimulasikan cocok dengan estimasi empiris yang diperoleh dari populasi target. Dikenal sebagai ‘pemodelan berorientasi pola’, pendekatan ini banyak digunakan untuk mengkalibrasi dan memvalidasi model berbasis individu (Gallagher et al. 2021 ; Grimm et al. 2006 ). Setelah mekanisme genetik yang menimbulkan variasi urutan virtual dikalibrasi, pengguna kemudian dapat menjalankan simulasi penyelamatan genetik dan membandingkannya berdasarkan seberapa besar pengaruhnya terhadap keragaman genetik virtual, perkawinan sedarah, atau beban genetik.
Kalibrasi parameter mekanistik yang berorientasi pada pola harus dipandu oleh jenis data sekuens yang tersedia. Misalnya, data yang tersedia mungkin mencakup estimasi pra-intervensi keragaman genetik yang diperoleh dari mikrosatelit atau polimorfisme nukleotida tunggal di seluruh genom. Estimasi empiris dari rangkaian homozigositas dan beban genetik dapat diperoleh dari populasi target jika data sekuensing genom secara keseluruhan tersedia. Sekuensing atau genotipe polimorfisme nukleotida tunggal di seluruh genom adalah cara yang paling mudah untuk menangkap elemen penting keragaman genetik tanpa perlu mendefinisikan peran adaptif gen fungsional pada populasi yang terancam (misalnya, Kardos et al. 2021 ). Data di seluruh genom mungkin juga menggambarkan wilayah dengan fungsi adaptif yang tidak diketahui. Simulasi bagaimana penyelamatan genetik menutupi kembali beban genetik yang terealisasi dan memecah rangkaian homozigositas adalah area menarik yang akan semakin berharga karena biaya dan tantangan analitis yang terkait dengan sekuensing genom secara keseluruhan terus menurun. Demikian pula, simulasi menawarkan alat yang ampuh untuk memandu penyelamatan genetik, yang menargetkan keragaman genetik pada daerah pengkodean tertentu, tetapi ini akan terbatas pada sistem di mana fungsi adaptif dari berbagai varian telah dikarakterisasi dengan baik (misalnya, gen kompleks imunohistokompatibilitas yang alel-alel tertentunya diketahui dapat meningkatkan ketahanan terhadap penyakit menular). Apa pun jenis data yang tersedia untuk populasi target, tujuannya adalah untuk mengembangkan model yang menghasilkan variasi sekuens dengan jenis dan resolusi yang sama dengan data yang darinya estimasi keragaman genetik (atau beban) diturunkan dalam populasi target.
5.2.2 Strategi 2: Simulasi Penyelamatan Genetik Menggunakan Variasi Urutan Empiris
Strategi lain melibatkan simulasi penyelamatan genetik populasi di mana frekuensi alel telah diinisialisasi dari data sekuens (penanda) (‘alel empiris’). Di sini, data sekuens dari populasi target dapat diimpor ke populasi yang disimulasikan (misalnya, melalui berkas FASTA atau VCF) pada waktu yang ditentukan pengguna berdasarkan saat data dikumpulkan. Misalnya, jika data dikumpulkan pada beberapa titik waktu selama penurunan populasi target, data dari peristiwa pengambilan sampel sebelumnya dapat diimpor ke simulasi pada waktu tertentu, dan efek dinamika populasi yang disimulasikan pada frekuensi alel (atau heterozigositas, beban genetik, dll.) dapat divalidasi dengan membandingkan hasil simulasi dengan data dari peristiwa pengambilan sampel selanjutnya. Ketika data untuk populasi target hanya tersedia dari sampel kontemporer, data tersebut dapat digunakan untuk menginisialisasi frekuensi alel pada titik dalam simulasi ketika kelimpahan dalam populasi virtual cocok dengan populasi nyata.
5.2.3 Strategi 3: Pendekatan Hibrida
Strategi ketiga menggabungkan elemen dari dua pendekatan sebelumnya dengan mengkalibrasi parameter mekanistik yang memediasi pengaruh umpan balik demo-genetik pada variasi sekuens virtual sebelum mengimpor data sekuens empiris ke dalam populasi yang disimulasikan. Setelah mekanisme genetik umpan balik demo-genetik dikalibrasi, pengguna dapat menjalankan simulasi burn-in tanpa alel virtual (untuk mensimulasikan kelimpahan ekuilibrium historis yang diikuti oleh penurunan), dan kemudian data sekuens dari populasi target dapat diimpor ke dalam populasi yang disimulasikan pada waktu yang tepat, seperti ketika ukuran populasi yang disimulasikan mirip dengan kelimpahan populasi nyata (jika diketahui) saat sampel dikumpulkan.
6 Menggunakan Simulasi untuk Menginformasikan Penyelamatan Genetika Terapan
6.1 Simulasi Skenario Alternatif Penyelamatan Genetik
Di sini, kami menggambarkan pendekatan untuk menggunakan simulasi guna membandingkan dan memberi peringkat skenario potensial alternatif penyelamatan genetik. Skenario alternatif yang dihadapi para pengambil keputusan mungkin berbeda untuk berbagai variabel, misalnya: ( i ) kelimpahan populasi ( N ) dari populasi target dan sumber (yaitu, estimasi ukuran populasi sensus Nc ); ( ii ) ukuran populasi efektif ( Ne ) dari populasi target dan sumber; ( iii ) daya dukung ( K ) dari populasi target; ( iv ) heterozigositas rata – rata dari populasi target; ( v ) diferensiasi genetik dari populasi target dan sumber; (vi) jumlah individu yang ditranslokasi dari populasi sumber (kohort yang ditranslokasi) ke populasi target; ( vii ) jumlah kejadian translokasi; ( viii ) waktu antara kejadian translokasi (frekuensi); ( ix ) rasio jenis kelamin dari kohort yang ditranslokasi; dan ( x ) distribusi usia (atau struktur-tahap) dari kohort yang ditranslokasi. Dalam simulasi spasial, variabel tambahan dapat mencakup: ( xi ) kepadatan populasi dan penyebaran populasi target, dan ( xii ) kemungkinan penyebaran berlebihan pasca-pelepasan individu yang ditranslokasi. Ini bukan daftar variabel yang lengkap untuk membandingkan skenario penyelamatan genetik, tetapi ini mencakup variabel yang paling mudah diubah di antara intervensi alternatif.
Variabel-variabel yang tercantum di atas dapat diubah untuk mensimulasikan skenario alternatif penyelamatan genetik, yang kemudian dapat dibandingkan dan diberi peringkat berdasarkan bagaimana mereka mempengaruhi hasil pada dinamika populasi yang disimulasikan dan komposisi genetik. ‘Mata uang’ dari simulasi adalah probabilitas kepunahan Pr(kepunahan) pada waktu tertentu t di masa depan, dan waktu rata-rata kepunahan ( T ext ) yang dihitung untuk skenario genetik yang berbeda. Kami memperingatkan terhadap penggunaan model proyeksi ke depan untuk memprediksi hasil tertentu dari penyelamatan genetik dan kelangsungan hidup populasi, yang bukan merupakan tujuan yang dimaksudkan—model proyeksi ke depan dapat digunakan untuk memberi peringkat keberhasilan relatif dari skenario penyelamatan genetik virtual dan memprioritaskan strategi dunia nyata berdasarkan probabilitas relatif untuk mengurangi depresi perkawinan sedarah dan meningkatkan kelangsungan hidup populasi dan keragaman genetik (Kotak 1 ).
KOTAK 1. Panduan langkah demi langkah untuk mengembangkan dan menerapkan simulasi demo-genetik penyelamatan genetik dalam konservasi.
- Tentukan tujuan intervensi penyelamatan genetik
- ○Identifikasi populasi sasaran dan tentukan tujuan strategis (misalnya, ukuran efek dan kerangka waktu untuk peningkatan keragaman genetik dan peningkatan pertumbuhan populasi per kapita).
- Kumpulkan data yang relevan
- ○Mengumpulkan atau mengumpulkan data demografi (misalnya, ukuran populasi, kelangsungan hidup dan kesuburan, struktur usia, rasio jenis kelamin)
- ○Mengumpulkan atau menghimpun data genetik (misalnya, frekuensi alel, koefisien perkawinan sedarah, heterozigositas, beban genetik)
- ○Mencakup data spasial: kesesuaian habitat, permeabilitas lanskap, dan distribusi populasi
- Pilih dan pelajari perangkat lunak pemodelan
- ○Pilih platform perangkat lunak yang sesuai berdasarkan kemampuan yang dibutuhkan (Tabel 3 untuk perbandingan serangkaian program yang representatif)
- ○Memahami dokumentasi perangkat lunak dan antarmuka pengguna atau bahasa pengkodean (jika berlaku)
- Mengembangkan kerangka kerja model
- ○Tentukan model demografi dasar, termasuk mekanisme untuk stokastisitas demografi dan umpan balik kepadatan
- ○Tambahkan proses genetik (misalnya, simulasi mutasi, rekombinasi, pergeseran, efek seleksi)
- ○Menghubungkan proses demografi dan genetik untuk memungkinkan adanya umpan balik demografi dan genetik
- Parameterisasi model
- ○Gunakan data empiris untuk menetapkan parameter proses demografi dan genetik
- ○Jika data empiris tidak tersedia, gunakan nilai dari spesies terkait, prediksi teoritis, atau hubungan alometrik.
- ○Lakukan analisis sensitivitas untuk mengidentifikasi parameter kritis dan mengukur ketidakpastian
- Simulasikan skenario dasar dan intervensi
- ○Jalankan simulasi dasar untuk menetapkan skenario kontrol (misalnya, tidak ada intervensi)
- ○Mengembangkan dan mensimulasikan skenario penyelamatan genetik dengan memvariasikan faktor-faktor seperti ukuran translokasi, frekuensi, dan populasi sumber (lihat Simulasi skenario alternatif penyelamatan genetik untuk serangkaian variabel yang perlu dipertimbangkan)
- ○Sertakan komponen spasial jika diperlukan, seperti konektivitas habitat atau dinamika penyebaran
- Menganalisis keluaran simulasi
- ○Bandingkan hasil skenario alternatif menggunakan metrik seperti kemungkinan kepunahan, waktu kepunahan, keragaman genetik, dan heterozigositas.
- ○Fokus pada ukuran efek (bukan pada uji hipotesis Neyman-Pearson) untuk memberi peringkat skenario dan mengidentifikasi intervensi yang optimal
- Validasi model (jika memungkinkan)
- ○Bandingkan hasil simulasi dengan data observasi dari populasi target atau populasi serupa
- ○Gunakan pemodelan berorientasi pola untuk menyempurnakan parameter jika ditemukan ketidaksesuaian
- Menafsirkan dan menerapkan hasil
- ○Merangkum temuan dalam hal kinerja skenario relatif (misalnya, perubahan risiko kepunahan)
- ○Memberikan rekomendasi yang jelas untuk menerapkan penyelamatan genetik (misalnya, ukuran kelompok yang optimal, jumlah translokasi)
- Ulangi dan adaptasikan
- ○Perbarui model dengan data baru saat tersedia
- ○Menilai ulang skenario untuk menyempurnakan rekomendasi sebagai respons terhadap perubahan kondisi atau wawasan tambahan
7 Kesimpulan
Kami telah menyoroti bagaimana umpan balik demo-genetik dapat dimasukkan ke dalam model simulasi yang menguji manfaat relatif penyelamatan genetik. Sementara teori memprediksi bahwa umpan balik demo-genetik memainkan peran penting dalam probabilitas kepunahan (yaitu, mendorong populasi ke dan menjaga mereka dalam pusaran kepunahan) (Benson et al. 2016 ; Caughley 1994 ; Gilpin dan Soulé 1986 ; Melbourne dan Hastings 2008 ), dan beberapa validasi empiris ada (Fagan dan Holmes 2006 ; Willi dan Hoffmann 2009 ), tidak ada contoh yang diterbitkan yang menunjukkan bagaimana umpan balik demo-genetik memengaruhi hasil penyelamatan genetik virtual. Kurangnya bukti ini terjadi meskipun perangkat lunak yang tersedia sekarang memiliki kemampuan untuk memasukkan umpan balik demo-genetik dalam prediksi kelangsungan hidup populasi relatif.
Ada beberapa peluang dan keterbatasan untuk pengembangan model simulasi penyelamatan genetik, dan bagaimana model tersebut dapat diterapkan untuk menginformasikan manajemen konservasi. Beberapa keterbatasan tersebut muncul karena tantangan dalam membuat parameter model dengan data empiris, yang dalam banyak kasus akan menjadi tantangan atau tidak mungkin dikumpulkan (terutama untuk spesies yang terancam). Kami menyarankan bahwa memilih untuk tidak melanjutkan penyelamatan genetik karena keterbatasan data berisiko menimbulkan rasa puas diri dan bahwa risiko dan manfaat dari pencampuran atau tidak mencampur perlu dipertimbangkan dalam konteks ancaman langsung terhadap populasi kecil (< 1000 individu yang disensus). Dengan kata lain, tidak adanya tindakan masih merupakan keputusan manajemen yang berpotensi memengaruhi persistensi spesies yang terancam. Dalam kasus data yang hilang, kami menganjurkan penggunaan model yang diparameterisasi dengan hubungan alometrik, spesies analog (misalnya, kongenerik), atau asumsi wajar lainnya dan interpretasi yang cermat dari keluaran model yang dipandu oleh analisis sensitivitas global untuk mengukur dan menyoroti ketidakpastian.
Populasi spesies yang terancam semakin berisiko punah akibat saling memperkuatnya proses demografi dan genetik yang terkikis oleh hilangnya dan degradasi habitat, spesies invasif, eksploitasi, perubahan iklim, dan interaksi sinergisnya. Bahkan jika perlindungan habitat dan pemulihan ekologi tumbuh pesat sesuai kebutuhan, konsekuensi genetik dari kemacetan populasi dapat terus berlanjut dan merusak pemulihan populasi. Mengembangkan model simulasi untuk menginformasikan penyelamatan genetik berpotensi meningkatkan kepercayaan diri praktisi konservasi untuk menerapkan intervensi dan menawarkan panduan tentang cara meningkatkan kemungkinan keberhasilan sambil menghindari konsekuensi negatif yang tidak diinginkan. Simulasi juga menawarkan jalan yang menarik untuk penelitian masa depan, dengan banyak pertanyaan luar biasa yang masih perlu diperhatikan (Kotak 2 ).
KOTAK 2. Pertanyaan-pertanyaan yang masih belum terjawab.
Model penyelamatan genetik harus bertujuan untuk menjawab pertanyaan-pertanyaan berikut:
- Seberapa sensitifkah probabilitas kepunahan terhadap jumlah individu yang ditranslokasi ke populasi target dan frekuensi translokasi tersebut?
- Berapa lama (dalam skala generasi) yang dibutuhkan untuk mengamati dampak dalam hal menutupi beban genetik dan meningkatkan kelangsungan hidup populasi target? Hal ini khususnya relevan dalam konteks garis waktu pengelolaan yang realistis dan saran untuk membedakan antara dampak jangka pendek (F 1 , F 2 ) versus jangka panjang (F 3 ) dari penyelamatan genetik (Hoffmann et al. 2021 ).
- Seberapa intensif populasi target perlu diambil sampelnya untuk memperkirakan efek penyelamatan genetik pada heterozigositas dan keragaman genetik yang diamati?
- Apakah konfigurasi spasial translokasi memberikan dampak yang nyata pada hasil penyelamatan genetik? Pertimbangan ini mungkin hanya relevan dalam keadaan tertentu (misalnya, spesies dengan penyebaran rendah, cagar alam berpagar kecil atau suaka pulau).


