Abstrak
- Bunga Campanula (Campanulaceae) dikunjungi oleh lebah generalis dan spesialis dari berbagai famili. Sementara lebah generalis mengandalkan volatil umum dalam aroma bunga Campanula , lebah spesialis Megachilidae terutama menggunakan spiroacetal yang tidak biasa untuk pengenalan bunga inang, yang biasanya hanya dilaporkan dalam komunikasi serangga. Meskipun evolusi spesialisasi dalam Megachilidae dan adaptasi sensorik yang mendasarinya telah lama menjadi topik yang menarik, evolusi sensitivitas penciuman reseptor antena untuk spiroacetal belum pernah diselidiki sebelumnya.
- Dalam penelitian ini, kami memadukan elektrofisiologi dan filogenetik untuk mengevaluasi evolusi kepekaan penciuman terhadap spiroasetal pada megachilid spesialis Campanula dibandingkan dengan lebah lain, termasuk generalis dan spesialis yang mencari makan pada bunga selain Campanula .
- Kami menunjukkan bahwa spesialis Campanula megachilid sangat sensitif terhadap spiroacetal dan bahwa senyawa ini dapat dideteksi oleh sebagian besar Megachilidae lainnya dan Andrena bicolor (Andrenidae) yang umum. Lebih jauh, analisis filogenetik kami menunjukkan bahwa kemampuan untuk mendeteksi spiroacetal mungkin merupakan sifat leluhur di Megachilidae dan kesamaan dalam sensitivitas antena tidak dapat dijelaskan hanya dengan keterkaitan filogenetik.
- Kami menyimpulkan bahwa kapasitas untuk merasakan spiroacetals merupakan eksaptasi dalam Megachilidae, yang merupakan suatu kondisi sebelum evolusi spesialis Campanula yang diakibatkan oleh peran spiroacetals dalam komunikasi serangga, sementara kepekaan tinggi pada spesialis Campanula telah berevolusi kemudian sebagai respons terhadap tingkat emisi rendah senyawa ini pada bunga inangnya.
Baca Ringkasan Bahasa Sederhana gratis untuk artikel ini di blog Jurnal.
1. PENDAHULUAN
Perilaku lebah, yang termasuk dalam banyak spesies dan genus yang berbeda, merupakan dasar penyerbukan sekitar 280.000 tanaman berbunga dari seluruh dunia (Kearns et al., 1998 ; Michener, 2007 ). Lebah-lebah ini memanfaatkan sumber daya bunga, terutama serbuk sari dan nektar dan dengan demikian secara tidak sengaja menyerbuki bunga. Makanan serbuk sari yang dikumpulkan oleh lebah untuk keturunannya sebagai sumber makanan utama sangat bervariasi. Sementara beberapa spesies lebah mengumpulkan serbuk sari dari spektrum tanaman yang sangat luas yang tidak selalu berhubungan dekat (disebut ‘generalis serbuk sari’ atau ‘polilega’; misalnya lebah madu), yang lain terspesialisasi dan sangat bergantung pada serbuk sari dari sangat sedikit spesies tanaman yang termasuk dalam genus atau famili yang sama (misalnya Cane & Sipes, 2006 ) atau bahkan satu spesies. Penelitian telah menunjukkan bahwa larva ‘spesialis serbuk sari’ atau ‘oligolega’ ini tidak dapat berkembang pada serbuk sari dari tanaman bukan inang (Praz, Müller & Dorn, 2008 ).
Untuk menemukan dan memilih tanaman inang, penyerbuk menggunakan isyarat bunga yang berbeda (Leonard & Masek, 2014 ), termasuk warna, bentuk, tekstur (Chittka et al., 2005 ), aroma (Raguso, 2008 ) dan medan elektrostatik (Clarke et al., 2013 ). Namun, aroma bunga diyakini sebagai sinyal utama untuk daya tarik lebah terhadap bunga, terutama dalam asosiasi yang melibatkan penyerbuk khusus (Dötterl & Vereecken, 2010 ). Aroma bunga sering kali mengandung banyak senyawa organik volatil yang termasuk dalam beberapa kelompok kimia dan jalur metabolisme. Buket aroma sangat berbeda tergantung pada spesies tanaman baik karena rasio relatif yang unik dari berbagai senyawa yang umum terjadi atau karena adanya senyawa yang sangat spesifik, yang jarang ditemukan di alam (Knudsen et al., 2006 ). Oleh karena itu, mereka dapat bertindak sebagai sumber informasi yang hampir tak terbatas bagi penyerbuk. Cara lebah mencari makan memanfaatkan informasi ini berbeda-beda menurut preferensi serbuk sari mereka. Sementara para generalis menggunakan volatil umum untuk membedakan bunga-bunga yang termasuk dalam spesies tanaman yang berbeda atau untuk mengevaluasi ketersediaan serbuk sari dan nektar pada bunga-bunga dari spesies yang sama (lihat Ayasse et al., 2000 ; Burger et al., 2010 ; Milet-Pinheiro et al., 2012 ), para spesialis menggunakan volatil bunga yang tidak biasa untuk mengidentifikasi tanaman inangnya (Burger et al., 2012 ; Milet-Pinheiro et al., 2013 ). Biasanya, spesies lebah yang terspesialisasi memiliki adaptasi spesifik dalam sirkuit penciuman, yang dapat diekspresikan hanya melalui kehadiran (atau jumlah yang lebih tinggi) reseptor penciuman yang sensitif terhadap volatil bunga spesifik inang (Renou, 2014 ). Hal ini menghasilkan sensitivitas tinggi terhadap senyawa-senyawa dalam karangan bunga inang, sementara spesies lebah lainnya jauh kurang sensitif atau tidak dapat merasakannya sama sekali (Brandt et al., 2017 ; Schäffler et al., 2015 ).
Megachilidae adalah famili lebah berlidah panjang yang sangat beragam, mencakup lebih dari 4000 spesies dan 76 genera yang dideskripsikan di seluruh dunia (Michener, 2007 ). Dalam famili tersebut, spesialisasi telah berkembang dalam klade yang berbeda yang melibatkan kelompok tanaman inang yang berbeda, seperti Asteraceae, Ranunculaceae, dan Campanulaceae (Praz, Müller, Danforth et al., 2008 ). Lebah megachilid yang beradaptasi dengan serbuk sari Campanula (‘ Campanula specialist’) termasuk dalam genera yang berbeda, seperti Chelostoma , Osmia , dan Hoplitis . Meskipun bunga Campanula juga dikunjungi dan diserbuki oleh spektrum generalis yang luas (misalnya lebah madu dan lebah) yang termasuk dalam famili lain (Blionis & Vokou, 2001 ; Westrich, 1989 ), spesialis megachilid Campanula adalah penyerbuk terpenting dalam beberapa spesies Campanula (Schlindwein et al., 2005 ). Senyawa utama yang hadir dalam aroma bunga Campanula adalah beberapa konstituen yang paling banyak ditemukan yang dilaporkan dalam bunga (yaitu ( E )-β-ocimene, 2-phenylethanol dan linalool; lihat, misalnya Dudareva & Pichersky, 2006 ). Senyawa-senyawa ini memicu respons antena dan perilaku yang kuat pada spesies lebah yang berbeda (Dötterl & Vereecken, 2010 ; Milet-Pinheiro et al., 2016 ; Pecetti et al., 2002 ), termasuk pengunjung Campanula , terlepas dari tingkat spesialisasi mereka (Brandt et al., 2017 ; Milet-Pinheiro et al., 2016 ). Selain itu, rangkaian aroma juga mengandung spiroasetal, yang jarang terdeteksi sebagai konstituen aroma bunga selain spesies Campanula (Milet-Pinheiro et al., 2013 ), dalam jumlah relatif sangat rendah (Milet-Pinheiro et al., 2015 ). Senyawa yang sangat spesifik ini memainkan peran penting dalam pengenalan inang pada spesialis Campanula Chelostoma rapunculi (Milet-Pinheiro et al., 2015 ). Selain itu, spiroasetal memainkan peran yang sangat penting dalam komunikasi serangga. Misalnya, campuran spiroasetal disekresikan sebagai feromon seks oleh banyak spesies Coleoptera, Hymenoptera (tawon, lebah) dan Diptera (Booth et al., 2009 ; Francke & Kitching, 2001 ) atau sebagai feromon agregasi (misalnya pada lebah soliter Andrena wilkella ; Francke & Kitching, 2001 ). Selain itu, mereka dapat bertindak sebagai penolak (Francke & Kitching, 2001), juga ditemukan dalam racun spesies tawon umum (yaitu Vespula vulgaris dan V. germanica ; Weston et al., 1997 ).
Dalam studi baru-baru ini, Brandt et al. ( 2017 ) membandingkan sensitivitas antena pengunjung khusus dan umum Campanula , mengungkap bahwa empat spiroacetals berbeda dapat dirasakan oleh beberapa spesialis Campanula (yaitu Ch. rapunculi , Chelostoma campanularum dan Hoplitis mitis ) bahkan dalam konsentrasi yang sangat rendah satu bagian per miliar (0,000001 μL·mL −1 ). Sebaliknya, generalis (yaitu Andrena bicolor , Apis mellifera dan Bombus terrestris ) tampaknya tidak merasakan senyawa kimia ini sama sekali atau hanya dalam konsentrasi yang jauh lebih tinggi daripada yang terjadi secara alami pada bunga Campanula (Brandt et al., 2017 ). Secara keseluruhan, hasil ini mendukung hipotesis kendala yang diusulkan oleh Sedivy et al. ( 2008 ), yang menunjukkan bahwa pilihan lebah terhadap tanaman inang terutama ditentukan oleh kendala evolusi yang terkait dengan adaptasi sensorik, fisiologis atau perilaku dan membatasi jangkauan tanaman yang dapat dimanfaatkan lebah secara efektif untuk serbuk sari. Akibatnya, spesialisasi pada bunga Campanula mungkin didorong tidak hanya oleh faktor ekologi tetapi juga oleh keterbatasan intrinsik, seperti sensitivitas spesies lebah tertentu yang berevolusi terhadap spiroacetals. Selain itu, hasilnya menimbulkan pertanyaan tentang kemampuan untuk melihat spiroacetals dalam spesies Megachilidae yang berbeda dibandingkan dengan pengunjung Campanula yang termasuk dalam famili lebah lain (yaitu Apidae dan Andrenidae; Michener, 2007 ). Berbeda dengan asumsi awal dan yang telah lama berlaku, garis keturunan Megachilidae generalis telah diturunkan dari nenek moyang yang terspesialisasi (Danforth et al., 2006 ; Larkin et al., 2008 ; Sedivy et al., 2008 ). Akan tetapi, tidak diketahui apakah hal ini terkait dengan kemampuan untuk merasakan aroma bunga spesifik inang Campanula melalui reseptor penciuman di antena yang sensitif terhadap spiroasetal. Tidak diketahui pula apakah kemampuan ini merupakan sifat dasar dalam Megachilidae atau apakah kemampuan ini telah berevolusi beberapa kali secara terpisah sebagai respons terhadap spesialisasi pada bunga Campanula .
Dalam penelitian ini, kami memadukan analisis perbandingan elektrofisiologi dan filogenetik untuk (1) memperluas pengetahuan tentang sensitivitas antena spesies lebah yang berbeda terhadap spiroacetal dan (2) menyelidiki evolusi sensitivitas antena terhadap spiroacetal dalam Megachilidae dengan spesies Andrenidae dan Apidae sebagai kelompok luar. Lebih khusus lagi, kami mengajukan pertanyaan-pertanyaan berikut: (1) Apakah spesialis Campanula megachilid lebih sensitif terhadap spiroacetal spesifik inang daripada spesies lebah dengan preferensi serbuk sari lain yang termasuk dalam Megachilidae dan famili lain (yaitu Andrenidae dan Apidae)? (2) Apakah evolusi sensitivitas antena terhadap aroma bunga spesifik inang dibentuk oleh spesialisasi pada serbuk sari Campanula atau oleh keterkaitan filogenetik? (3) Apakah kapasitas untuk merasakan spiroacetal berevolusi sebelum atau bersamaan dengan perkembangan spesialisasi pada serbuk sari Campanula di Megachilidae?
2 BAHAN DAN METODE
2.1 Spesies lebah
Kami menguji 11 spesies lebah dengan preferensi serbuk sari yang berbeda (Gambar 1 ). Selain tiga spesialis Campanula , kami menguji satu spesies yang mengkhususkan diri pada Echium (Boraginaceae), satu spesialis Ranunculus (Ranunculaceae), satu spesialis Asteraceae, serta lima spesies generalis yang diketahui mengunjungi bunga Campanula (selanjutnya diklasifikasikan sebagai ‘lebah dengan preferensi serbuk sari lainnya’). Lebah ditangkap selama dua tahun berikutnya menggunakan jaring serangga (BugDorm, MegaView Science Supplies Co., Ltd., Taichung, Taiwan) dan disimpan dalam kotak pendingin hingga analisis elektrofisiologis. Untuk setiap spesies, kami menguji 12 individu, kecuali Ho. mitis , yang menggunakan 13 individu (lihat Tabel 1 ). Informasi lebih lanjut tentang spesies lebah yang diuji termasuk waktu dan lokasi pengumpulan disediakan dalam Tabel S1 di Informasi Tambahan S01 . Studi ini tidak memerlukan persetujuan etis untuk penggunaan serangga.
| Skala inferensi | Skala di mana perlakuan yang menarik diterapkan | Jumlah replikasi pada skala yang sesuai |
|---|---|---|
| Individu | Jenis | 12–13 |

2.2 Pengukuran elektroantografi
Untuk pengukuran elektroantennografi (EAGs) kami mengikuti pendekatan Brandt et al. ( 2017 ). Kami mencatat respons antena masing-masing lebah terhadap empat spiroacetal berbeda yang umumnya terdapat pada bunga Campanula : (1) 1,6-dioxaspiro[4,5]decane [alias, 1,6-dioxaspirodecane], (2) ( E )-7-methyl-1,6-dioxaspiro[4.5]decane [alias, ( E )-conophthorin], (3) (2R,6S)-2-metil-1,7-dioxaspiro[5.5]undecane dan (4) (5 S ,7 S )-7-metil-1,6-dioxaspiro[4.5]decane [alias, (5 S ,7 S )-conophthorin]. Semua spiroasetal disintesis oleh Wittko Francke di Institut Kimia Organik, Universitas Hamburg. Selain itu, kami memperoleh dan menguji secara komersial tiga senyawa yang dilaporkan sebagai komponen aroma bunga Campanula , yang juga umum ditemukan pada tumbuhan angiospermae lainnya (Knudsen et al., 2006 ): satu senyawa aromatik, yaitu 2-feniletanol (99%; Sigma-Aldrich) dan dua monoterpena, yaitu ß-osimena (campuran isomer, 90%; Sigma-Aldrich) dan linalool (campuran rasemat, 97%; Sigma-Aldrich). Larutan masing-masing zat kimia pada empat konsentrasi berbeda disiapkan: 1 μL·mL −1 (10 −3 ), 0,01 μL·mL −1 (10 −5 ), 0,0001 μL·mL −1 (10 −7 ) dan 0,000001 μL·mL −1 (10 −9 ). Kami menggunakan n -heksana (99%; Merck) sebagai pelarut. Stimulus aroma diterapkan pada setiap sediaan antena pada interval 90 detik, dimulai dan diakhiri dengan n -heksana sebagai referensi. Bahan kimia diuji dalam urutan acak (ditentukan di RStudio) dengan konsentrasi yang meningkat secara progresif. Untuk setiap stimulus, strip kertas saring berukuran 0,5 cm × 1 cm diresapi dengan 10 μL larutan uji dan ditempatkan dalam pipet Pasteur berukuran 15 cm (VWR International, Darmstadt, Jerman). Semprotan udara standar (0,3 detik, 25 mL/detik) diaplikasikan menggunakan pengontrol stimulus (CS-05; Syntech, Hilversum, Belanda). Untuk pengukuran elektroantenografi, kami menguji N = 12 individu untuk semua spesies kecuali Ho. mitis ( N = 13) dan menganalisis respons antena menggunakan perangkat lunak Syntech EAG (EAGPro, v.2.1; Ockenfels SYNTECH EAGPro, nd). Perangkat lunak merekam potensi antena yang dijumlahkan setelah stimulasi. Ini divisualisasikan sebagai hiperpolarisasi garis dasar. Amplitudo maksimum, diukur dalam mV, didefinisikan sebagai ‘respons antena’ dan bervariasi berdasarkan keberadaan dan jumlah neuron reseptor penciuman yang merespons pada spesies yang berbeda. Informasi terperinci tentang pengaturan, persiapan, dan penerapan rangsangan aroma dapat ditemukan di Brandt et al. ( 2017 ) dan di Informasi Pendukung S02 .
2.3 Analisis statistik respons antena terhadap volatil umum dan spiroasetal
2.3.1 Perbandingan dalam spesies
Dengan menggunakan perangkat lunak Syntech, respons dinormalkan berdasarkan kekuatan respons terhadap n -heksana untuk mengoreksi penurunan sensitivitas antena selama pengukuran. Berikut ini, respons antena yang dinormalkan ini akan disebut sebagai ‘respons absolut’. Untuk menguji perbedaan di antara empat konsentrasi yang diuji dan n -heksana dalam setiap spesies, kami melakukan analisis model efek campuran linier univariat [LMM; faktor tetap: mudah menguap dalam setiap konsentrasi yang diuji; faktor acak: individu lebah] berdasarkan kumpulan data masing-masing spesies yang diuji. Kami mempertimbangkan perbedaan yang signifikan antara respons EAG dari stimulus uji sebagai bukti bahwa zat tersebut dapat dideteksi pada konsentrasi tersebut. LMM dan perbandingan berpasangan berdasarkan estimasi rata-rata marginal (EMM) dilakukan di RStudio (Tim RStudio, 2024 ) menggunakan paket lme4 v.1.1.34 (Bates et al., 2014 ) dan emmeans v.1.8.9 (Lenth et al., 2021 ).
2.3.2 Perbandingan antar spesies
Karena respons EAG absolut berbeda di antara spesies dan dalam spesies di antara individu, kami membakukan respons antena. Oleh karena itu, kami menggunakan pengukuran antena setiap lebah terhadap berbagai konsentrasi senyawa tertentu untuk menghitung regresi linier dan mengekstrak intersep dan kemiringan menggunakan fungsi ‘lm’ dan ‘summary’ dari R. Intersep menunjukkan posisi saat garis regresi melintasi sumbu Y dan menunjukkan kekuatan respons secara keseluruhan. Kemiringan menunjukkan seberapa kuat respons EAG meningkat dengan meningkatnya konsentrasi setiap senyawa yang diuji. Mengenai intersep dan kemiringan, kami mengharapkan hasil yang serupa dengan hasil dari Brandt et al. ( 2017 ), dengan nilai yang lebih tinggi untuk spesialis Campanula dibandingkan dengan semua spesies lain yang diuji untuk spiroacetal, sementara tidak ada perbedaan yang diharapkan untuk volatil umum. Nilai intersep dan kemiringan dibandingkan untuk spiroacetal dan volatil umum antara spesialis Campanula dan spesies lebah yang diuji dengan preferensi serbuk sari lainnya dalam uji jumlah peringkat Wilcoxon non-parametrik dengan koreksi kontinuitas menggunakan paket stats yang diimplementasikan dalam R.
Berdasarkan matriks Bray Curtis multivariat (menggunakan empat spiroacetal yang diuji atau tiga volatil umum sebagai variabel), kami melakukan empat analisis PERMANOVA dua faktorial yang berbeda dengan perbandingan berpasangan (faktor: preferensi serbuk sari [yaitu Campanula spp., generalis, Ranunculus spp., Asteraceae spp. dan Echium spp.] dan spesies [bersarang dalam preferensi serbuk sari]) untuk menguji perbedaan dalam intersep dan kemiringan dalam respons terhadap spiroacetal dan volatil umum, masing-masing (Anderson et al., 2008 ).
Untuk mengilustrasikan variasi respons antena di antara spesies dengan preferensi serbuk sari yang berbeda, kami menggunakan penskalaan multidimensi non-metrik (nMDS; Clarke & Gorley, 2006 ), berdasarkan kesamaan Bray-Curtis yang dihitung untuk data intersepsi (rata-rata per spesies). Kami melakukan analisis SIMPER untuk menentukan senyawa yang paling ditanggapi spesies tersebut.
Matriks dan analisis untuk perbandingan antarspesies dihitung dalam perangkat lunak ‘PRIMER 6’ (versi 6.1.15; PRIMER-E Ltd. 2012) dengan add-on ‘PERMANOVA +’ (versi 1.0.5; PRIMER-E Ltd. 2012). Tingkat signifikansi ditetapkan pada α ≤ 0,05 untuk semua analisis statistik. Untuk semua PERMANOVA, kami menggunakan jumlah kuadrat tipe III (parsial), efek tetap berjumlah nol untuk suku campuran, permutasi residual dalam model tereduksi, dan 9999 permutasi.
2.4 Evolusi sensitivitas terhadap zat volatil dan spiroasetal umum
Untuk menyelidiki pola evolusi respons antena terhadap spiroasetal dan volatil umum di seluruh filogeni spesies lebah yang diuji, kami merekonstruksi pohon filogenetik yang mencakup semua 11 spesies. Kami menggunakan satu gen mitokondria ( CO1 ) dan tiga gen nuklir ( EF1-a , LWR dan CAD ) sebagai penanda, yang tersedia dalam basis data GenBank (NCBI: National Center for Biotechnology Information, 2024 ). Nomor aksesi untuk setiap penanda DNA yang digunakan diberikan dalam Informasi Pendukung S01 . Kami memperoleh beberapa penyelarasan menggunakan MUSCLE (Edgar, 2004 ) yang diimplementasikan dalam MEGA v.8.0 (Kumar et al., 2018 ). Filogeni direkonstruksi menggunakan BEAST v.1.10.4 (Suchard et al., 2018 ) dan diproses dengan menggunakan Tracer v.1.7.1 , LogCombiner v.1.10.4 , dan TreeAnnotator v.1.10.4. Untuk visualisasi pohon kredibilitas klade maksimum (MCC), kami menggunakan FigTree v.1.3.1 (Rambaut et al., 2018 ). Rincian lebih lanjut tentang langkah-langkah ini disediakan dalam Informasi Pendukung S03 dan dalam Dataset 1 (Brandt et al., 2025 ) yang berisi data penyelarasan yang dirangkai dan berkas masukan BEAST.
Bahasa Indonesia: Untuk lebih memahami adaptasi evolusioner sensitivitas terhadap volatil bunga umum dan spiroasetal pada spesies lebah yang kami uji, kami menggabungkan filogeni yang direkonstruksi dan matriks data berdasarkan respons antena terstandarisasi (pengurangan respons absolut terhadap n -heksana dari respons absolut terhadap setiap senyawa yang diuji untuk setiap individu yang diuji) untuk semua senyawa dalam konsentrasi 1 μL·mL −1 , yang menunjukkan keberadaan reseptor penciuman (yaitu kemampuan untuk merasakan suatu senyawa). Kami melakukan analisis filogenetik komparatif dalam R menggunakan paket ape v .5.7.1 (Paradis et al., 2022 ), phytools v .1.9.16 (Revell, 2012 ) dan geiger v .2.0.7 (Pennell et al., 2014 ).
Mengikuti Martel et al. ( 2019 ), kami menghitung apa yang disebut ‘sinyal filogenetik untuk sifat-sifat berkelanjutan’ berdasarkan respons antena multivariat menggunakan uji statistik K Blomberg (lihat Blomberg et al., 2003 ) berdasarkan 1000 pengacakan (fungsi ‘phylosig’). Dalam uji ini, sinyal filogenetik diukur berdasarkan varians sifat yang diharapkan berdasarkan model gerak Brown yang mirip dengan pergeseran genetik (Kamilar & Cooper, 2013 ).
Kami mengamati pola filogenetik respons antena dengan mencocokkan dan membandingkan dua model evolusi sifat yang berbeda: (1) model Gerak Brown (BM) multivariat laju tunggal (simulasi pergeseran genetik; probabilitas divergensi respons antena meningkat secara seragam dari waktu ke waktu) dan (2) model Orenstein-Uhlenbeck (OU) optimum tunggal (simulasi optimum global; varians respons antena menurun dari waktu ke waktu). Dengan demikian, kami menggunakan kumpulan data univariat (yaitu respons terhadap senyawa tertentu) dan fungsi ‘fitContinuous’ untuk menghitung estimasi parameter dan nilai kemungkinan (kriteria informasi Akaike yang dikoreksi; AICc).
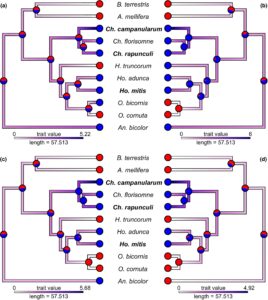
Untuk memahami apakah dan bagaimana semua spesies lebah yang diuji dapat mendeteksi senyawa yang diuji, kami merekonstruksi status leluhur menggunakan dua sifat: (1) keberadaan reseptor yang sensitif terhadap senyawa (karakter diskret) dan (2) sensitivitas terhadap senyawa tersebut (karakter kontinu; diukur dengan respons antena yang terstandarisasi). Kami melakukan Estimasi Kemungkinan Maksimum dari kedua jenis karakter tersebut dengan pohon MCC untuk setiap senyawa yang diuji.
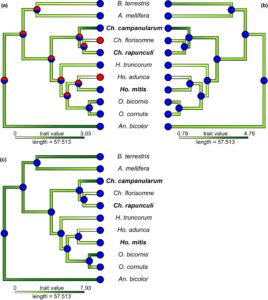
Untuk representasi grafis dari analisis gabungan, kami (1) membuat pohon filogenetik disertai dengan peta panas yang mencerminkan respons antena rata-rata terstandarisasi (fungsi ‘phylo.heatmap’) dan (2) menghitung filogeni yang menunjukkan perkiraan riwayat evolusi nilai sifat sebagai variabel kontinu (fungsi ‘contMap’) yang dikombinasikan dengan status karakter (yaitu kemampuan untuk memahami zat volatil yang diuji seperti yang ditawarkan dalam LMM) sebagai variabel kualitatif (fungsi ‘plotTree’) dalam rekonstruksi status leluhur.
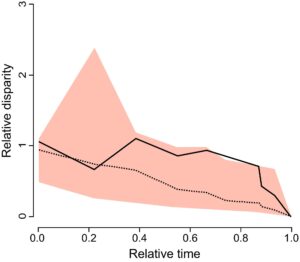
Selain itu, kami membuat plot disparitas melalui waktu (DTT) (fungsi ‘dtt’). Plot ini menunjukkan bagaimana respons antena menempati ruang sifat sepanjang sejarah evolusi garis keturunan yang disertakan dalam studi kami (Revell, 2012 ) dengan membandingkan lintasan DTT yang diamati di seluruh filogeni dengan respons antena terhadap senyawa pada konsentrasi tertentu yang disimulasikan dalam model BM (Harmon et al., 2003 ). Pengukuran area antara perbedaan DTT rata-rata yang diamati dan disimulasikan dilakukan setelah Harmon et al. ( 2003 ) menunjukkan disparitas yang diamati dan disimulasikan dengan menggunakan statistik indeks keanekaragaman morfologi (MDI) (interval kepercayaan 95% untuk total 1000 simulasi; tingkat signifikansi: α ≤ 0,05).
3 HASIL
3.1 Respon antena terhadap volatil umum dan spiroasetal
Hasil perbandingan antarspesies dari respons EAG absolut terhadap n -heksana dibandingkan dengan setiap senyawa kimia dalam empat konsentrasi berbeda memberikan indikasi yang baik tentang apakah senyawa yang diuji dapat dideteksi oleh setiap spesies yang diuji atau tidak. Sebagian besar spesies, baik spesialis Campanula atau bukan, merespons secara signifikan terhadap tiga volatil umum pada konsentrasi serendah 0,01 μL·mL −1 (untuk sebagian besar spesies) atau 0,0001 μL·mL −1 ( An. bicolor ). Sementara respons signifikan terhadap spiroacetals ditunjukkan untuk spesialis Campanula pada konsentrasi serendah 0,0001 μL·mL −1 ( Ho. mitis dan Ch. rapunculi ) atau 0,000001 μL·mL −1 ( Ch. rapunculi ), lebah dengan preferensi serbuk sari lainnya menunjukkan respons signifikan terhadap spiroacetals hanya pada konsentrasi tertinggi yang diuji (1 μL·mL −1 ; An. bicolor , Ch. florisomne , H. truncorum , Ho. adunca dan O. bicornis ) atau tidak sama sekali ( A. mellifera , B. terrestris dan O. cornuta ). Hasil tersebut diberikan dalam Informasi Pendukung S04 .
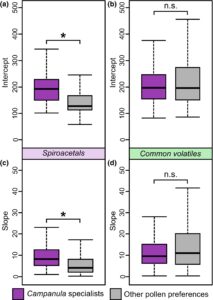
Intersep garis regresi linier berbeda secara signifikan antara spesialis Campanula dan lebah dengan preferensi serbuk sari lainnya untuk spiroacetals (uji jumlah peringkat Wilcoxon: W = 12.419, p < 0,0001; Gambar 2a ), tetapi tidak untuk volatil umum ( W = 16.651; p = 0,52; Gambar 2b ). Demikian pula, ada perbedaan yang signifikan dalam kemiringan yang membandingkan spesialis Campanula dan semua spesies lain yang diuji untuk spiroacetals ( W = 17.225; p < 0,0001; Gambar 2c ) tetapi tidak untuk volatil umum ( W = 16.733; p = 0,47; Gambar 2d ).
Hasil analisis permutasi mengungkapkan perbedaan signifikan dalam kekuatan respons antena terhadap spiroacetal di antara spesies dengan preferensi serbuk sari yang berbeda (PERMANOVA: Pseudo- F 4,122 = 13,96, p < 0,001) tanpa perbedaan di antara spesies dalam kelompok ini (PERMANOVA: Pseudo- F 6,122 = 1,66, p = 0,12; Gambar 3a ). Ketika membandingkan respons terhadap volatil umum, kekuatannya berbeda baik di antara preferensi serbuk sari (PERMANOVA: Pseudo- F 4,122 = 3,95, p < 0,01) dan spesies dalam kelompok ini (PERMANOVA: Pseudo- F 6,122 = 2,72, p < 0,05; Gambar 3b ).

Tidak ada tingkat sinyal filogenetik yang signifikan dalam respons antena terhadap salah satu spiroacetals [1,6-dioxaspirodecane: K = 0,29, p = 0,74; ( E )-conophthorin: K = 0,32, p = 0,66; (2 R ,6 S )-2-methyl-1,7-dioxaspiro[5.5]undecane: K = 0,29, p = 0,75; (5S,7R)-conophthorin: K = 0,32, p = 0,61; n = 77 untuk semua] maupun terhadap zat volatil umum (β-ocimene: K = 0,31, p = 0,66; 2-phenylethanol: K = 0,31, p = 0,67; linalool: K = 0,32, p = 0,64; n = 77 untuk semua) pada konsentrasi 1 μL·mL −1 berdasarkan uji statistik K Blomberg.
Dengan demikian, estimasi parameter terhitung dan kemungkinan evolusi karakter berkelanjutan, berdasarkan spesies yang kami uji, menunjukkan bahwa baik kekuatan respons antena terhadap spiroacetals (model BM: sigSq <0,0001 dan AICc >46,8) maupun terhadap volatil umum (model BM: sigSq <0,0001 dan AICc >40,5) tidak dapat dijelaskan oleh keterkaitan filogenetik saja (Gambar 4 ). Respons antena optimal tidak disarankan untuk senyawa kimia yang diuji (sigSq <0,0001 dan AICc >40 untuk semua senyawa yang diuji; model OU).
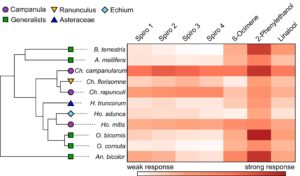
Berdasarkan analisis kami, semua spesialis Campanula yang diuji serta spesialis non- Campanula Ch. florisomne dan Ho. adunca mampu merasakan keempat spiroacetal yang diuji (Gambar 6 ). Sebaliknya, sebagian besar generalis yang mencari makan pada berbagai spesies tanaman mampu merasakan tidak ada atau hanya satu dari spiroacetal yang diuji. Andrena bicolor dengan demikian merupakan pengecualian karena spesies generalis ini mampu merasakan 1,6-dioxaspirodecane, (2 R ,6 S )-2-methyl-1,7-dioxaspiro[5.5]undecane dan ( E )-conophthorin. Rekonstruksi status leluhur untuk 11 spesies yang kami uji mengindikasikan bahwa kemampuan untuk merasakan empat spiroacetals yang diuji adalah sifat dasar dalam Megachilidae yang dijadikan sampel (meskipun tidak sepenuhnya teratasi untuk 1,6-dioxaspirodecane) dan kondisi tersebut telah hilang setidaknya dalam dua kesempatan untuk merasakan ( E )-conophthorin dan setidaknya dalam tiga kesempatan untuk merasakan (2 R ,6 S )-2-methyl-1,7-dioxaspiro[5.5]undecane dan (5S,7R)-conophthorin. Sementara kapasitas untuk merasakan spiroacetals tampaknya merupakan leluhur dalam Chelostoma dan Hoplitis , itu tidak terjadi pada Osmia (Gambar 6 ). Dengan demikian, rekonstruksi kami mengindikasikan bahwa kepekaan terhadap spiroacetals terbatas pada leluhur Megachilidae yang diuji, tetapi sangat berevolusi pada spesialis Campanula atau spesies yang terkait erat dengan mereka (misalnya Ch. florisomne ; Gambar 6 ).
4 DISKUSI
Studi kami mengungkap bahwa sebagian besar spesies yang diuji, terlepas dari preferensi serbuk sari, menunjukkan respons antena yang kuat terhadap tiga volatil umum yang diuji. Namun, hanya spesialis Campanula yang mampu merasakan spiroacetals spesifik inang dalam konsentrasi rendah, sementara beberapa spesies dengan preferensi serbuk sari lainnya tidak merespons sama sekali terhadap spiroacetals. Pemodelan evolusi kami menunjukkan bahwa sensitivitas antena terhadap spiroacetals dan volatil umum tidak terkait dengan keterkaitan filogenetik dan oleh karena itu sensitivitas diferensial adalah hasil adaptasi evolusi. Lebih penting lagi, hasil kami menunjukkan bahwa kemampuan untuk merasakan spiroacetals (yaitu keberadaan reseptor penciuman yang sensitif) mungkin merupakan kondisi leluhur yang berevolusi sebelum evolusi spesialis Campanula di Megachilidae dan oleh karena itu mungkin mewakili eksaptasi yang dapat menyebabkan evolusi spesialisasi ke Campanula di Megachilidae.
Sesuai dengan Brandt et al., ( 2017 ), kami menemukan sensitivitas yang jauh lebih tinggi terhadap spiroacetals pada Campanula specialist dibandingkan pada spesies yang diuji dengan preferensi serbuk sari lain yang tampaknya tidak dapat merasakan senyawa kimia ini sama sekali atau hanya dalam konsentrasi tinggi 1 μL·mL −1 (lihat juga Brandt et al., 2017 ). Meskipun ketiga spesialis Campanula , spesialis Ranunculus Ch. florisomne dan spesialis Echium Ho. adunca mampu mendeteksi semua spiroacetals yang diuji, hanya spesialis Campanula yang merespons konsentrasi rendah di mana aroma bunga spesifik inang ini benar-benar dipancarkan di alam (Milet-Pinheiro et al., 2013 , 2015 ). Untuk mempersepsikan jumlah yang rendah ini (Milet-Pinheiro et al., 2013 , 2015 ), sirkuit penciuman perifer dari spesialis Campanula harus sangat beradaptasi dengan isyarat spesifik dari bunga inangnya dalam bentuk keberadaan dan/atau jumlah reseptor penciuman atau protein pengikat (Hansson & Stensmyr, 2011 ; Schäffler et al., 2015 ). Ini menggarisbawahi apa yang disebut ‘hipotesis kendala yang meyakinkan’ atau ‘memaksa’ dari Sedivy et al. ( 2008 ) yang menyatakan bahwa, pilihan tanaman inang lebah terutama ditentukan oleh kendala evolusi yang terkait dengan adaptasi sensorik, fisiologis atau perilaku dan membatasi jangkauan tanaman yang dapat dimanfaatkan lebah secara efektif untuk serbuk sari. Hal ini sering kali menghasilkan preferensi untuk garis keturunan tanaman tertentu (misalnya berbagi sifat kimia atau morfologi tertentu) pada spesies yang mengeksploitasi tanaman inang yang tidak mungkin dilakukan oleh yang lain yang pada akhirnya dapat mengarah pada spesialisasi. Pentingnya sensitivitas yang kuat disorot oleh ketergantungan lebah spesialis Campanula yang baru muncul dan belum pernah mencari makan, yang sepenuhnya bergantung pada bahan kimia ini untuk menemukan bunga inangnya (misalnya Ch. rapunculi ; Milet-Pinheiro et al., 2013 ). Namun, ketika mendapatkan lebih banyak pengalaman mencari makan dari waktu ke waktu, lebah juga belajar untuk mengasosiasikan volatil yang lebih umum, seperti linalool dan β-ocimene, dengan nektar dan serbuk sari bunga Campanula . Dengan demikian, lebah Ch. rapunculi yang lebih berpengalaman dapat membatasi kunjungan mereka ke bunga yang lebih bermanfaat dan meningkatkan efisiensi mencari makan mereka (Milet-Pinheiro et al., 2013 ).
Dalam analisis kami, volatil umum membangkitkan respons yang kuat pada sebagian besar spesies terlepas dari preferensi serbuk sari dan keterkaitan filogenetiknya. Ketika disajikan kepada penyerbuk secara individual, volatil umum sering menunjukkan tingkat respons antena yang optimal dalam dua model kami (model BM dan OU) yang menunjukkan seleksi stabilisasi yang bertindak untuk mempertahankan respons antena yang kuat atau lemah terhadap senyawa kimia tertentu di seluruh filogeni (Hansen, 1997 ). Misalnya, Brandt et al. ( 2021 ) menunjukkan bagaimana senyawa yang ditemukan sangat umum dalam aroma bunga parfum, di antaranya linalool, mengungkapkan respons tingkat optimal di seluruh pola respons antena penyerbuk lebah euglossine jantannya. Namun, untuk semua spesies lebah dalam penelitian kami saat ini, β-ocimene, 2-phenyl ethanol dan linalool kemungkinan besar penting untuk daya tarik penyerbuk hanya jika disajikan bersama dengan senyawa kimia lainnya. Memang, penelitian sebelumnya telah menunjukkan bahwa proporsi relatif dan campuran spesifik memainkan peran kunci untuk mencapai spesifisitas dalam sinyal penciuman dengan penggunaan senyawa yang relatif sederhana dan ada di mana-mana saja (Ayasse & Dötterl, 2014 ; Proffit et al., 2020 ). Ketika menafsirkan hasil kami, penting untuk mengakui bahwa sementara elektroantenografi adalah alat yang berharga untuk menganalisis keberadaan reseptor penciuman dan sensitivitas spesies terhadap volatil tertentu (Wadhams, 1990 ), itu tidak memberikan informasi langsung tentang perilaku lebah. Meskipun senyawa yang memicu respons perilaku pada lebah biasanya menimbulkan respons antena (Schorkopf et al., 2011 ), signifikansi ekologis volatil spesifik dalam interaksi lebah-tanaman harus ditafsirkan dengan hati-hati tanpa adanya informasi tentang pemrosesan saraf tingkat tinggi di otak serangga dan data perilaku yang sesuai.
Analisis filogenetik kami menunjukkan bahwa dua spesialis Campanula Ch. campanularum dan Ch. rapunculi lebih dekat hubungannya dengan spesialis Ranunculus Ch. florisomne daripada dengan spesialis Campanula Ho. mitis . Hubungan yang lebih dekat ini menunjukkan bahwa sejarah evolusi bersama dapat menjelaskan mengapa Ch. florisomne dapat mendeteksi spiroacetal meskipun tidak mengkhususkan diri pada Campanula . Meskipun demikian, profil respons antena ternyata lebih mirip di antara lebah dengan preferensi serbuk sari yang sama daripada di antara spesies yang lebih dekat hubungannya. Megachilid lain yang diuji dapat mendeteksi hanya satu (yaitu spesialis Asteraceae H. truncorum dan generalis O. bicornis ) atau tidak ada spiroacetal sama sekali (yaitu generalis O. cornuta ), bahkan pada konsentrasi yang sangat tinggi. Ini berarti bahwa meskipun semua spesies ini termasuk dalam famili yang sama, sensitivitas terhadap spiroacetal sangat bervariasi tergantung pada preferensi serbuk sari. Selain itu, An. bicolor dari famili Andrenidae mampu merasakan tiga dari empat spiroacetals dan karena itu memiliki reseptor penciuman yang sensitif terhadap konstituen aroma bunga yang jarang terjadi ini. Alasan yang mungkin untuk hasil di An. bicolor bisa jadi karena spesies bivoltine ini terutama mencari makan di Campanula pada generasi keduanya (Milet-Pinheiro et al., 2016 ) yang juga digunakan untuk percobaan. Temuan ini digarisbawahi oleh statistik K Blomberg yang menunjukkan tidak ada sinyal filogenetik yang terkait dengan sensitivitas diferensial terhadap spiroacetals pada spesies megachilid dan non-megachilid An. bicolor . Semua hasil tersebut menyiratkan bahwa faktor-faktor lain selain dari keterkaitan filogenetik harus dipertimbangkan untuk sepenuhnya menjelaskan jalur evolusi yang mengarah pada kemampuan (atau ketidakmampuan) untuk merasakan spiroacetals pada spesies yang kami uji.
Secara umum, telah lama diasumsikan bahwa status leluhur garis keturunan lebah adalah generalis dan bahwa faktor-faktor yang berbeda, seperti persaingan antarspesies (Robertson, 1899 ) atau efisiensi mencari makan (Lovell, 1914 ), telah memilih spesialisasi tanaman inang dan juga bertindak untuk mempertahankannya (Müller, 1996 ). Namun, Sedivy et al. ( 2008 ) menilai tanaman inang dari 35 spesies Chelostoma yang berbeda dan menunjukkan bahwa setidaknya dua generalis serbuk sari telah berevolusi dari leluhur spesialis dalam Chelostoma . Kebanyakan megachilid dari penelitian kami yang tidak terspesialisasi pada Campanula memang merasakan spiroacetals, semua atau beberapa, tetapi pada saat yang sama kurang sensitif terhadapnya. Ini menunjukkan bahwa reseptor penciuman yang sensitif terhadap spiroacetals lebih merupakan sifat dasar untuk sebagian besar genus Chelostoma daripada sifat yang telah berkembang sebagai adaptasi untuk secara khusus menemukan bunga Campanula , yang juga mendukung temuan Sedivy et al. ( 2008 ). Khususnya, rekonstruksi leluhur dan model evolusi kita, berdasarkan spesies yang diuji, menunjukkan hilangnya kepekaan terhadap spiroacetals dalam genera Osmia , Heriades dan Hoplitis . Spesies-spesies ini bisa saja beralih dari mencari makan di Campanula ke tanaman lain, seperti Asteraceae (yaitu H. truncorum ), atau bisa saja memperluas spektrum tanaman inangnya menjadi generalis seperti O. bicornis dan O. cornuta . Semua ini mendukung gagasan bahwa keberadaan reseptor penciuman yang peka terhadap spiroacetals telah berevolusi sebelum evolusi spesialisasi Campanula di Megachilidae dan bahwa ini telah menjadi salah satu prasyarat evolusi yang penting bagi evolusi spesialis Campanula . Karena spiroacetals banyak dilaporkan dalam komunikasi serangga, keberadaan reseptor penciuman yang sensitif terhadap spiroacetals dapat mewakili sebuah eksaptasi: Sementara senyawa-senyawa spesifik ini bertindak baik sebagai feromon seks dan agregasi atau sebagai pengusir dalam banyak keluarga serangga yang tidak berhubungan (Booth et al., 2009 ; Francke & Kitching, 2001 ; Weston et al., 1997 ), bunga Campanula mungkin telah berevolusi untuk mengeksploitasi bias sensorik yang ada pada lebah Megachilid, menggunakan kembali spiroacetals untuk menarik mereka untuk penyerbukan.
Untuk pemahaman yang lebih baik, evolusi aroma Campanula dan, khususnya, evolusi spiroacetals dalam aroma juga harus dipertimbangkan. Dalam banyak sistem tanaman, preferensi penciuman bawaan spesifik dari penyerbuk masing-masing membentuk evolusi komposisi aroma bunga (Milet-Pinheiro & Gerlach, 2017 ; Schiestl & Johnson, 2013 ). Namun, bunga Campanula dikunjungi oleh berbagai pengunjung yang termasuk dalam genera dan famili berbeda yang menunjukkan preferensi aroma yang berbeda (Brandt et al., 2017 ). Oleh karena itu, tekanan selektif yang bekerja pada aroma bunga Campanula oleh penyerbuk mungkin tidak sekuat dalam sistem lain. Dalam penelitian terbaru, Milet-Pinheiro et al. ( 2021 ) membandingkan aroma dan sifat bunga lain dari spesies Campanula yang berbeda untuk menyelidiki pengaruh filogeni pada evolusi sifat. Hasil mereka menunjukkan bahwa evolusi aroma bunga di Campanula , setidaknya sebagian, dibatasi oleh keterkaitan filogenetik. Pada saat yang sama, keberadaan senyawa lain serta seluruh komposisi rangkaian aroma tampaknya dibentuk oleh faktor eksternal, seperti penyerbuk atau herbivora. Jelas, untuk sepenuhnya mendukung asumsi bahwa kepekaan terhadap spiroacetal mungkin telah mendorong evolusi spesialisasi ke Campanula di Megachilidae, analisis elektroantennografi tambahan dari lebih banyak spesies megachilid, pengambilan sampel filogenetik yang lebih luas, dan eksperimen perilaku diperlukan. Secara khusus, menggabungkan lebih banyak spesies lebah dari genera Andrena , Chelostoma dan Osmia ke dalam analisis filogenetik akan secara signifikan meningkatkan pemahaman kita. Namun, hasil kami memberikan titik awal yang penting untuk penyelidikan ini dan menyoroti kebutuhan untuk menghubungkan temuan ini dengan perkembangan evolusi aroma bunga pada spesies Campanula lainnya .
Secara keseluruhan, hasil kami menunjukkan bahwa spesialis Campanula sangat sensitif terhadap aroma bunga spesifik inang dibandingkan dengan spesies lebah dengan preferensi serbuk sari lainnya, dengan demikian mendukung hipotesis kendala yang kuat mengenai evolusi jangkauan inang pada lebah dalam genus Chelostoma . Selain itu, kapasitas untuk merasakan spiroacetal tampaknya merupakan kondisi leluhur yang dapat berevolusi sebelum evolusi spesialis Campanula di Megachilidae; namun, hasil kami menunjukkan bahwa kepekaan terhadap aroma bunga inang tertentu telah berevolusi secara independen dari filogeni dan dengan demikian kesamaan dalam kepekaan adalah hasil dari evolusi konvergen ke spesialisasi Campanula . Secara keseluruhan, kami menawarkan wawasan pertama tentang evolusi kepekaan penciuman untuk spiroacetal di Megachilidae, yang tampaknya menjadi eksaptasi utama untuk evolusi spesialis Campanula megachilid .