
Abstrak
- Kualitas lingkungan selama perkembangan individu secara umum dianggap memiliki efek jangka panjang terhadap kinerja di masa dewasa, tetapi hal ini terutama didasarkan pada studi observasi yang tidak dapat menunjukkan jalur kausal di balik efek jangka panjang tersebut.
- Dalam penelitian ini, kami melakukan uji coba terkontrol acak untuk menguji efek kausal dari pola makan yang buruk terhadap kinerja pertumbuhan, reproduksi, dan kelangsungan hidup selama masa hidup burung puyuh Jepang betina ( Coturnix japonica ). Semua individu ditempatkan dalam kondisi standar saat dewasa untuk memungkinkan pemisahan efek lingkungan perkembangan dari efek lingkungan dewasa.
- Pola makan yang buruk menyebabkan pertumbuhan yang sangat berkurang, yang menunda permulaan reproduksi dan mengakibatkan ukuran tubuh yang berkurang sepanjang masa dewasa, dibandingkan dengan pola makan standar. Meskipun tidak ada efek yang terdeteksi pada tingkat bertelur dan kelangsungan hidup pada usia tertentu, meskipun terjadi penuaan yang kuat pada ciri-ciri kebugaran ini, betina yang dibesarkan dengan pola makan yang buruk menghasilkan telur yang lebih kecil.
- Oleh karena itu, meskipun tidak ada pengaruh lingkungan perkembangan yang buruk terhadap tingkat bertelur dan kelangsungan hidup betina, lingkungan perkembangan seekor induk memengaruhi ukuran tubuhnya saat dewasa dan lingkungan yang ia sediakan bagi keturunannya selama perkembangan embrio.
- Kami menduga bahwa efek pada ukuran betina dewasa dan ukuran telur dapat menyebabkan efek ‘sendok perak’ di alam liar, jika ukuran yang lebih besar memberikan keuntungan dalam persaingan atas sumber daya. Namun, kami tidak dapat mengesampingkan bahwa ukuran yang lebih kecil akan menyebabkan kebutuhan makanan yang lebih rendah, sehingga bertindak sebagai ‘respons adaptif prediktif’ terhadap lingkungan yang buruk.
1. PENDAHULUAN
Kondisi yang buruk selama perkembangan individu umumnya dianggap memiliki efek negatif jangka panjang pada performa di masa dewasa, di seluruh taksa, yang juga dapat bermanifestasi sebagai peningkatan penuaan di akhir kehidupan (misalnya, Cooper & Kruuk, 2018 ; Hamel et al., 2009 ; Hayward et al., 2013 ; Lummaa & Clutton-Brock, 2002 ; Nussey et al., 2007 ; Reid et al., 2003 ; van de Pol et al., 2006 ; Victora et al., 2008 ). Namun, karena menilai efek jangka panjang tersebut (sering disebut sebagai efek ‘sendok perak’; Grafen, 1988 ) memerlukan studi jangka panjang, pandangan umum ini terutama didasarkan pada studi korelatif yang tidak dirancang apriori untuk secara khusus menunjukkan jalur kausal yang mengarah pada korelasi positif antara kualitas kondisi perkembangan dan kinerja di akhir kehidupan.
Dari perspektif riwayat hidup, korelasi positif antara kualitas lingkungan perkembangan dan kinerja di masa dewasa dapat muncul karena berbagai proses. Lingkaran umpan balik positif dapat menciptakan proses di mana kondisi buruk individu dalam satu tahap kehidupan secara langsung menyebabkan kondisi buruk pada tahap berikutnya (Fokkema et al., 2021 ). Ketika ada persaingan untuk sumber daya, tertinggal selama perkembangan dapat membatasi kemampuan kompetitif individu, menyebabkan individu tersebut semakin tertinggal dan, jika bertahan hidup hingga mandiri, menetap di lingkungan yang tidak optimal dan akibatnya mencapai keberhasilan reproduksi atau kelangsungan hidup dewasa yang lebih rendah (Fokkema et al., 2021 ; van de Pol et al., 2006 ; Verhulst et al., 1997 ). Misalnya, pada rusa roe ( Capreolus capreolus ), kebugaran dewasa yang berkurang pada anak rusa yang lahir akhir sebagian besar dapat dijelaskan oleh massa tubuh mereka yang lebih rendah sepanjang hidup (Plard et al., 2015 ).
Pada saat yang sama, keterbatasan sumber daya dapat menyebabkan trade-off dalam individu antara kinerja saat ini dan kinerja masa depan (misalnya, Lemaître et al., 2015 ; Metcalfe & Monaghan, 2001 ). Ketika alokasi untuk pertumbuhan mengorbankan alokasi untuk sifat-sifat yang lebih penting di kemudian hari (misalnya, perbaikan DNA), individu yang menghadapi kekurangan sumber daya selama perkembangan dapat secara strategis mengalokasikan sumber daya mereka untuk menanggung beban biaya kebugaran di akhir hidup (Kirkwood, 1977 ). Studi eksperimental jangka pendek yang memanipulasi kondisi perkembangan dan menilai efeknya pada proksi fisiologis kualitas somatik jangka panjang, seperti panjang telomer, memang menunjukkan bahwa ini adalah fenomena yang sering terjadi (Chatelain et al., 2020 ; Monaghan, 2014 ). Namun, mengasumsikan konsekuensi kebugaran jangka panjang berdasarkan hasil tersebut mungkin tidak tepat (Froy et al., 2021 ; Vedder, Moiron, et al., 2022 ). Oleh karena itu, studi eksperimental yang benar-benar mengukur performa jangka panjang, dalam hal kebugaran, diperlukan untuk menunjukkan apakah kondisi perkembangan yang buruk memiliki konsekuensi kebugaran jangka panjang akibat adanya pertukaran dalam individu.
Selain itu, hipotesis ‘respons adaptif prediktif’ menyatakan bahwa kondisi buruk yang dialami selama perkembangan secara adaptif memprogram metabolisme individu untuk mengatasi kondisi serupa saat dewasa (Gluckman & Hanson, 2004 ). Namun, jika kondisi perkembangan yang buruk tidak diimbangi dengan kondisi buruk saat dewasa, hal ini dapat menyebabkan penurunan kesehatan yang berdampak pada kebugaran di masa dewasa (Rickard & Lummaa, 2007 ). Meskipun asumsi yang diperlukan agar mekanisme ini berkembang dikritik, terutama pada aspek kondisi perkembangan yang memberikan perkiraan akurat tentang kondisi masa depan (misalnya, Nettle et al., 2013 ; Rickard & Lummaa, 2007 ; Wells, 2007 ), hipotesis ini mungkin juga memberikan penjelasan untuk efek negatif dari kondisi perkembangan yang buruk pada kinerja di lingkungan dewasa yang tidak cocok (Monaghan, 2008 ).
Meskipun jalur-jalur yang disebutkan di atas untuk konsekuensi kebugaran jangka panjang dari lingkungan perkembangan yang buruk mungkin tidak saling eksklusif, jalur-jalur tersebut berbeda berkenaan dengan pentingnya lingkungan dewasa. Sementara manipulasi eksperimental dari lingkungan perkembangan dapat menetapkan efek kausal dari variabel yang dimanipulasi (misalnya, Spagopoulou et al., 2020 ), lingkungan dewasa tidak boleh dikorelasikan dengan lingkungan perkembangan untuk mengesampingkan kemungkinan bahwa efek jangka panjang dari lingkungan perkembangan hanya bertindak dengan menentukan kualitas lingkungan dewasa, alih-alih melalui efek langsung melalui trade-off dalam individu atau ‘respons adaptif prediktif’ yang tidak cocok. Dalam hal ini, penelitian pada hewan dalam penangkaran memiliki manfaat karena mereka dapat lebih mudah memanipulasi lingkungan perkembangan terlepas dari lingkungan dewasa masa depan seorang individu. Namun, melakukan eksperimen semacam itu pada spesies vertebrata di mana semua komponen kebugaran dapat diukur pada tingkat individu sepanjang rentang hidup lengkap semua individu yang menjadi sasaran eksperimen, dengan ukuran sampel yang memadai, merupakan tantangan logistik.
Burung puyuh Jepang ( Coturnix japonica ) adalah spesies burung galliform kecil yang sangat cocok untuk percobaan semacam itu. Anak burung bersifat precocial dan dapat dengan mudah dibesarkan secara mandiri dari induknya, yang memungkinkan manipulasi kualitas lingkungan perkembangan tanpa variabel pengganggu apa pun. Untuk seekor burung, mereka memiliki riwayat hidup yang sangat cepat, dengan individu yang mampu mencapai reproduksi dalam waktu 2 bulan setelah menetas (Vedder, Bichet, et al., 2022 ). Selain itu, karena burung puyuh peliharaan telah dipilih terutama untuk kinerja bertelur (Lukanov & Pavlova, 2020 ) ‘kebugaran’ mereka (yaitu, apa yang dimaksimalkan oleh seleksi) dan kinerja reproduksi dapat dengan mudah diukur dari kelangsungan hidup mereka dan jumlah serta ukuran telur yang mereka taruh (Vedder, Bichet, et al., 2022 ). Penuaan dalam kelangsungan hidup dan bertelur dimulai segera setelah dimulainya bertelur, dan masa hidup mereka jarang melebihi 3 tahun (Beccardi et al., 2024 ; Vedder, Bichet, et al., 2022 ; Woodard & Abplanalp, 1971 ), yang memungkinkan pemantauan kinerja dan tingkat penuaan seumur hidup dalam jangka waktu yang dapat diperkirakan.
Dalam studi ini kami secara eksperimental memanipulasi kondisi perkembangan pascanatal anak burung puyuh Jepang hingga kematangan seksual, dengan secara acak memberi mereka makanan pemeliharaan standar atau berkualitas buruk (lihat juga Vedder et al., 2023 ). Dari anak burung ini, 218 betina kemudian dipelihara secara komunal dalam satu lingkungan hingga mereka mati secara alami. Kami memantau semua komponen kebugaran (awal reproduksi, kelangsungan hidup harian, dan kesuburan spesifik usia), alokasi per kapita untuk reproduksi (massa telur segar), dan ukuran dewasa dan massa tubuh spesifik usia selama rentang hidup mereka yang lengkap. Ini memungkinkan kami untuk menguji apakah kondisi buruk selama perkembangan memiliki efek jangka panjang pada kinerja dewasa atau tingkat penuaan, terlepas dari lingkungan dewasa, dan apakah ini memiliki efek yang bertahan lama pada massa tubuh sepanjang masa dewasa.
2 METODE
2.1 Pembiakan
Bahasa Indonesia: Untuk memperoleh anak ayam untuk percobaan, kami mengawinkan dengan burung puyuh Jepang dewasa berusia sekitar 1 tahun pada bulan Maret–Juni 2019 dan 2020 di Institut Penelitian Unggas di Wilhelmshaven, Jerman. Pada kedua tahun tersebut, kami melakukan empat putaran pembiakan, dengan total 96 pasang pada tahun 2019 dan total 80 pasang pada tahun 2020. Setiap putaran, pasangan-pasangan ditempatkan di kandang pembiakan tempat mereka menghabiskan 10–14 hari bersama-sama. Pada usia ini, betina bertelur sekitar 0,8 butir telur per hari (Vedder, Bichet, et al., 2022 ), dan kami mengumpulkan dan menandai semua telur setiap hari. Jantan mudah kawin ketika ditempatkan bersama dengan betina, menyebabkan sebagian besar telur sudah subur setelah pasangan tersebut menghabiskan 2 hari di kandang pembiakan (Vedder, 2022 ). Sebelum berpasangan, betina ditempatkan bersama dan diisolasi dari jantan setidaknya selama empat belas hari untuk mencegah sperma yang disimpan dari jantan sebelumnya membuahi telur apa pun (lihat Birkhead & Fletcher, 1994 ). Telur yang dikumpulkan disimpan pada suhu 12°C dan diinkubasi secara artifisial per putaran, selalu dalam jangka waktu 14 hari setelah bertelur. Inkubasi dilakukan pada suhu 37,7°C dan kelembaban relatif 50% dengan inkubator otomatis penuh (Grumbach, ProCon automatic systems GmbH & Co. KG, Mücke, Jerman). Telur secara otomatis dibalik setiap jam selama 14 hari pertama inkubasi. Setelah 14 hari inkubasi, telur ditempatkan di kompartemen individual yang ditandai agar anakan telur dapat dihubungkan dengan telur tempat menetasnya. Inkubasi berikutnya dilakukan pada suhu 37,2°C dan kelembaban relatif 70%, tanpa pembalikan telur, dalam inkubator penetasan (Favorit, HEKA Brutgeräte, Rietberg, Jerman). Setelah 16 hari inkubasi, inkubator penetasan diperiksa sekali sehari untuk mencari telur baru hingga tidak ada lagi telur yang layak (setelah sekitar 19 hari inkubasi).
2.2 Prosedur percobaan dan pengumpulan data umum
Setelah menetas, semua anak ayam ditandai dengan cincin kaki plastik bernomor dan ditimbang hingga 0,01 g terdekat. Mereka didistribusikan secara acak di kandang pemeliharaan yang dipanaskan (109 × 57 × 25 cm, Kükenaufzuchtbox Nr 4002/C, HEKA Brutgeräte, Rietberg, Jerman) dengan salah satu dari dua diet pemeliharaan yang berbeda kualitasnya, sedemikian rupa sehingga keturunan dari pasangan yang sama dibagi secara kasar pada dua perlakuan diet. Secara total, kami menggunakan 8 kandang pemeliharaan (4 per perlakuan), dengan maksimum 30 anak ayam dari kedua jenis kelamin digabungkan secara bersamaan di kandang yang sama. Kami menggunakan diet pemeliharaan unggas standar dengan 21,0% protein, 4,0% lemak, dan 1,1% kalsium sebagai perlakuan pemeliharaan ‘standar’. Untuk perawatan pemeliharaan ‘buruk’, kami menggunakan diet yang mengandung 14,5% protein, 4,0% lemak, dan 1,0% kalsium (lihat Informasi Pendukung Online untuk tinjauan rinci kedua diet). Kedua diet memiliki nilai kalori yang sama sebesar 11,4 MJ/Kg dan merupakan pakan unggas yang tersedia secara komersial (GoldDott, DERBY Spezialfutter GmbH, Münster, Jerman), namun yang standar ditujukan untuk anak ayam yang sedang tumbuh, sedangkan yang ‘buruk’ ditujukan untuk ayam yang berbulu lengkap, tetapi belum bertelur. Kandungan protein yang lebih rendah dari diet ‘buruk’ diantisipasi untuk secara substansial mengurangi laju pertumbuhan tanpa mengorbankan kelangsungan hidup anak ayam (Weber & Reid, 1967 ). Kandungan protein dari diet standar diantisipasi untuk menghasilkan laju pertumbuhan normal tanpa menyebabkan gangguan yang terkait dengan pertumbuhan yang terlalu cepat (Weber & Reid, 1967 ). Makanan dan air disediakan sepuasnya, dan makanan digiling menjadi bubuk homogen untuk mencegah makan selektif. Suhu semua kandang pemeliharaan ditetapkan pada 37,0°C saat menetas dan secara bertahap diturunkan hingga 20–25°C selama 3 minggu. Kandang dipelihara dalam siklus terang–gelap 16–8 jam untuk kedua perlakuan, dan cincin kaki plastiknya diganti dengan cincin aluminium bernomor unik saat anak ayam berusia antara 14 dan 35 hari, tergantung pada ukuran kakinya. Semua anak ayam dipindahkan ke kandang burung luar ruangan saat berusia 21 hari. Untuk ini, kami menggunakan dua kandang burung yang terpisah tetapi identik, di mana mereka dipelihara dengan diet pemeliharaan yang sama seperti sebelumnya. Kandang burung ini dilengkapi dengan lampu pemanas dan menerima cahaya minimal 16 jam.
Kematian anak ayam rendah (5,7%), sebagian besar terjadi dalam beberapa hari pertama setelah menetas, dan tidak berbeda antara perlakuan diet (Vedder et al., 2023 ). Semua anak ayam yang bertahan hidup ditimbang setiap minggu setelah tanggal penetasan masing-masing, hingga 0,01 g terdekat pada 7 hari, dan hingga g terdekat pada semua umur berikutnya hingga berumur 84 hari ketika semua individu telah dewasa sepenuhnya. Pada umur 84 hari, tarsus dewasa masing-masing individu juga diukur, hingga 0,01 mm terdekat. Kami menentukan jenis kelamin semua anak ayam berdasarkan karakteristik bulu setelah berumur sekitar 28 hari. Sementara anak ayam jantan juga dipantau hingga berumur 84 hari sebagai bagian dari penelitian lain (Vedder et al., 2023 ), dalam penelitian ini kami hanya fokus pada betina karena hanya kesuburan mereka yang dipantau selama seluruh rentang hidup mereka. Dari umur 35 hari dan seterusnya, mereka diperiksa satu per satu untuk mengetahui permulaan bertelur setiap 2–3 hari. Hal ini dilakukan dengan memeriksa keberadaan telur di saluran telur melalui palpasi. Pada awal bertelur, setiap betina ditempatkan di kandang pembiakan selama seminggu. Di kandang-kandang ini, mereka semua menerima diet ayam petelur dewasa, dengan protein 19,0%, lemak 4,6%, dan kalsium 4,8%, dan nilai kalori 9,8 MJ/Kg (GoldDott, DERBY Spezialfutter GmbH, Münster, Jerman). Dengan cara ini, tingkat bertelur individu ditetapkan dengan memeriksa keberadaan telur yang baru diletakkan setiap hari selama 7 hari berturut-turut. Semua telur baru dikumpulkan dan ditimbang hingga 0,01 g terdekat dalam waktu satu hari setelah bertelur. Kemudian, pada tahun 2019 semua betina ( n = 151) dari kedua kelompok perlakuan dipindahkan ke satu kandang burung luar ruangan di mana semuanya menerima makanan ayam petelur dewasa yang sama (sepuasnya yang disediakan oleh beberapa tempat makan yang ditempatkan di seluruh kandang burung), dengan minimal 16 jam cahaya per hari untuk memastikan kelanjutan aktivitas reproduksi (Kovach, 1974 ). Pada tahun 2020, setengah dari semua betina juga ditempatkan dalam kondisi tersebut saat dewasa ( n = 66), sementara separuh lainnya ditempatkan dalam kondisi berbeda di musim dingin sebagai bagian dari eksperimen lain dan tidak disertakan dalam penelitian ini. Betina yang dikecualikan dipilih secara acak pada tingkat individu (menggunakan perangkat lunak untuk menghasilkan angka acak per individu, dan nilai batas dipilih untuk mengecualikan setengah dari betina dewasa secara seksual). Pada tahun 2019, tiga betina tidak pernah mulai bertelur (1 dengan diet standar, dan 2 dengan diet buruk) dan mereka dipindahkan ke kandang burung dengan diet dewasa saat sudah dewasa sepenuhnya dan tetap menjadi bagian dari penelitian. Semua kematian dewasa dipantau setiap hari, dan semua betina ditimbang dan dipantau sementara untuk kinerja reproduksi seperti yang dijelaskan di atas (tingkat bertelur dan massa telur selama 7 hari) pada interval sekitar 0,5 tahun selama masa hidup lengkap mereka. Penelitian ini tidak memerlukan pengambilan sampel invasif, dan semua prosedur yang melibatkan burung puyuh dilakukan di bawah lisensi “Veterinäramt JadeWeser” (izin no. 42508_03122020).
2.3 Analisis statistik
Meskipun kami sebelumnya telah melaporkan efek besar dari manipulasi diet pada pertumbuhan pascanatal untuk semua individu dari kedua jenis kelamin (Vedder et al., 2023 ), kami di sini menguji efek untuk subset betina yang ditempatkan sebagai orang dewasa dalam kondisi yang sama ( n = 217), yang mana 105 (48,4%) telah menerima diet buruk, dan 112 (51,6%) diet standar. Untuk tujuan ini, kami menggunakan model aditif Umum (GAM, fungsi ‘gam()’, paket ‘mgcv’), dengan distribusi kesalahan normal, dengan semua pengukuran massa tubuh mingguan sebagai variabel dependen. Untuk menguji apakah pertumbuhan berbeda antara kedua perlakuan diet, kami menambahkan perlakuan diet pemeliharaan (dua tingkat: standar/buruk) sebagai efek tetap dan interaksinya dengan usia sebagai variabel yang dihaluskan. Kami juga mengendalikan perbedaan potensial antar tahun, dengan menambahkan tahun (dua tingkat: 2019/2020) sebagai efek tetap. Sebagai efek acak, kami menambahkan ‘identitas pasangan’ induk, dan ‘identitas individu’, untuk memperhitungkan non-independensi saudara kandung, dan pengukuran berulang dari individu yang sama. Sementara GAM memiliki keuntungan karena tidak membatasi kurva pertumbuhan ke bentuk tertentu, ia memiliki kerugian karena tidak memungkinkan perbandingan langsung parameter pertumbuhan tertentu dengan penelitian lain. Oleh karena itu, kami juga menguji fungsi pertumbuhan yang umum digunakan (Gompertz, logistik, log-logistik, Weibull) yang paling sesuai dengan pertumbuhan yang diamati (dengan fungsi ‘drm()’ dari paket R ‘drc’) dan memberikan perbandingan parameter pertumbuhan antara diet untuk fungsi yang paling sesuai.
Untuk variabel dependen yang hanya diukur satu kali per individu (panjang tarsus dewasa, usia kematangan seksual, dan rentang hidup dewasa), kami menerapkan tiga model campuran Linear terpisah (LMM, fungsi ‘lmer()’, paket ‘lme4’), untuk menguji efek manipulasi diet pemeliharaan. Dalam model ini, perlakuan diet dan tahun penetasan dimasukkan sebagai efek tetap, dan ‘identitas pasangan’ induk sebagai efek acak.
Untuk menguji perbedaan spesifik usia dalam massa tubuh dewasa, dan massa telur, antara dua diet pemeliharaan, kami melakukan dua GAM terpisah dengan distribusi kesalahan normal dan massa tubuh, dan massa telur, sebagai variabel dependen, masing-masing. Model-model ini memasukkan perlakuan diet pemeliharaan sebagai efek tetap dan interaksinya dengan usia sebagai variabel yang dihaluskan. Juga dalam kasus ini, kami mengendalikan perbedaan potensial antara kelompok, dengan memasukkan tahun penetasan sebagai efek tetap. Selain itu, ‘identitas pasangan’ dan ‘identitas individu’ dimasukkan sebagai efek acak. Karena kami secara khusus tertarik pada perbedaan dalam pola massa tubuh/massa telur dalam individu dengan usia, kami memperhitungkan mortalitas selektif potensial dalam kaitannya dengan massa tubuh dewasa/massa telur dengan memasukkan rentang hidup dewasa sebagai prediktor linier kontinu (mengikuti van de Pol & Verhulst, 2006 ). Namun, jika diet memiliki efek yang sama pada rentang hidup seperti pada massa tubuh dewasa atau massa telur, mengoreksi rentang hidup dapat menutupi efek tersebut. Oleh karena itu, kami memusatkan variabel rentang hidup di sekitar rentang hidup rata-rata per perlakuan diet (dengan mengurangi rata-rata dari nilai mentah), sehingga tidak ada perbedaan keseluruhan dalam rentang hidup antara perlakuan diet. Selain itu, kami mengulang model pada massa telur dengan massa tubuh dewasa selama bertelur ditambahkan sebagai variabel yang dihaluskan, untuk menguji apakah ada juga efek diet pemeliharaan pada massa telur saat mengoreksi perbedaan dalam massa tubuh dewasa.
Efek dari manipulasi diet pemeliharaan pada probabilitas bertelur harian diuji dengan GAM dengan distribusi kesalahan binomial dan fungsi hubungan logit. Untuk analisis ini, variabel dependen terdiri dari 7 catatan binomial (1 = bertelur, 0 = tidak bertelur) per periode pengambilan sampel khusus usia. Perlakuan diet ditambahkan sebagai efek tetap dan kami secara khusus memasukkan istilah interaksi dengan usia sebagai variabel yang dihaluskan untuk menguji apakah diet pemeliharaan yang buruk dapat mempercepat penuaan reproduksi. Sekali lagi, kami memasukkan tahun penetasan sebagai efek tetap, dan ‘identitas pasangan’ dan ‘identitas individu’ sebagai efek acak. Umur dewasa (berpusat pada rata-rata per diet) dimasukkan sebagai prediktor linier berkelanjutan, untuk secara khusus menguji efek dalam individu dari perlakuan diet.
Akhirnya, untuk mempelajari efek manipulasi diet pada tingkat mortalitas harian saat dewasa, kami menggunakan model campuran aditif eksponensial sepotong-sepotong (PAMM), yang menggabungkan paket ‘pammtools’ (Bender et al., 2018 ) dengan model GAM yang dipasangkan dengan paket ‘mgcv’. Model tersebut menyertakan perlakuan diet dan tahun penetasan sebagai faktor tetap, interaksi antara perlakuan diet dan usia (variabel yang dihaluskan) sebagai istilah interaksi, dan ‘identitas pasangan’ sebagai efek acak. Dalam kasus ini, distribusi kesalahan Poisson diterapkan. Meskipun pendekatan ini direkomendasikan untuk data kelangsungan hidup spesifik usia yang tidak mengikuti bentuk yang terbentuk sebelumnya seiring bertambahnya usia (Bender et al., 2018 ), kami juga menguji perbedaan kelangsungan hidup spesifik usia antara perlakuan diet dengan model kelangsungan hidup parametrik. Karena kami sebelumnya telah menunjukkan bahwa fungsi Weibull memberikan kecocokan terbaik untuk data kelangsungan hidup burung puyuh Jepang (Beccardi et al., 2024 ), kami menjalankan model yang menguji perbedaan dalam skala fungsi Weibull (angka kematian awal) dan parameter bentuk (angka kematian berdasarkan usia) antara pola makan, dengan fungsi ‘flexsurvreg()’ dari paket R ‘flexsurv’. Model ini memperhitungkan perbedaan kohort potensial dalam kelangsungan hidup dengan membiarkan parameter skala bervariasi sesuai tahun penetasan.
Bahasa Indonesia: Untuk semua GAM yang dilakukan, kami menggunakan spline pelat tipis default fungsi gam() dengan 10 knot, yang diperkirakan menggunakan pendekatan kemungkinan maksimum (ML) (massa tubuh anak ayam, massa tubuh dewasa, massa telur) atau pendekatan kemungkinan maksimum terbatas (REML) (probabilitas bertelur harian, tingkat kematian dewasa harian). Karena GAM tidak menyediakan nilai-p untuk efek interaksi, kami menilai signifikansi efek spesifik usia dari manipulasi diet dengan memperkirakan penghalus terpisah untuk dua tingkat prediktor diet yang berbeda dan menghitung perbedaan antara penghalus dengan perkiraan 95% CI (fungsi ‘difference_smooths()’, paket ‘gratia’; Wood, 2017 , hlm. 294). Dengan tidak adanya efek keseluruhan dari diet pemeliharaan, ini seharusnya memungkinkan kita untuk menyimpulkan bahwa ada perbedaan antara diet pemeliharaan dalam rentang usia di mana interval kepercayaan tidak tumpang tindih dengan nol (lihat Informasi Pendukung Online ). Semua analisis dilakukan dalam perangkat lunak R (versi 4.1.3, R Foundation for Statistical Computing).
3 HASIL
3.1 Pertumbuhan dan ukuran dewasa
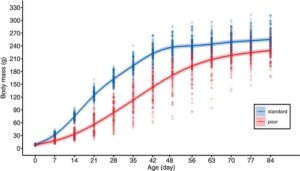
Dua diet pemeliharaan yang berbeda menyebabkan perbedaan ekstrem dalam pertumbuhan, dengan diet yang buruk menyebabkan massa tubuh yang lebih rendah dari pengukuran pertama setelah menetas, pada usia 7 hari, dan seterusnya (Gambar 1 , Gambar S1 , Tabel 1 ). Pada puncak pertumbuhan, betina yang menerima diet yang buruk beratnya sekitar 55% lebih rendah daripada betina yang menerima diet standar (Gambar 1 ). Meskipun perbedaan ini menjadi lebih kecil menjelang akhir tahap pertumbuhan, betina dengan diet yang buruk tidak dapat mengejar sepenuhnya dan tetap sekitar 10% lebih ringan saat dewasa (Gambar 1 ). Panjang tarsus mereka sekitar 5% lebih pendek saat dewasa pada usia 12 minggu (Gambar 2a , Tabel 2 ).
| Massa tubuh anak ayam (g) | |||
|---|---|---|---|
| Koefisien parametrik | Memperkirakan | Bahasa Inggris | nilai p |
| Mencegat | 175.748 | 1.958 | <0,001 |
| Pola makan (buruk) | -45.909 | 2.166 | <0,001 |
| Tahun penetasan (2020) | 1.299 | 2.760 | 0.638 |
| Istilah halus | Bahasa Inggris | Nilai F | nilai p |
| Usia: Diet (standar) | 8.597 | 6281.06 | <0,001 |
| Usia: Pola makan (buruk) | 7.682 | 5408.77 | <0,001 |
| Efek acak | Varians (SD) | 95% CI | nilai p |
| Identitas pasangan | 54 | 44,22–65,8 | 0,005 |
| Identitas individu | 211 | 182,6–260,7 | <0,001 |
| Sisa | 188 | 134,0–243,1 |
| Massa tubuh orang dewasa (g) | |||
|---|---|---|---|
| Koefisien parametrik | Memperkirakan | Bahasa Inggris | nilai p |
| Mencegat | 256.987 | 2.628 | <0,001 |
| Pola makan (buruk) | -13.046 | 2.755 | <0,001 |
| Tahun penetasan (2020) | -1,363 | 3.786 | 0.719 |
| Jangka hidup | 0,005 | 0,005 | 0,285 |
| Istilah halus | Bahasa Inggris | Nilai F | nilai p |
| Usia: Diet (standar) | 3.948 | 13.004 | <0,001 |
| Usia: Pola makan (buruk) | 4.377 | 10.862 | <0,001 |
| Efek acak | Varians (SD) | 95% CI | nilai p |
| Identitas pasangan | 131 | 122,9–147,3 | <0,001 |
| Identitas individu | 219 | 206,7–236,9 | <0,001 |
| Sisa | 399 | 380.1–420.1 |

| Efek acak | Perbedaan | Efek tetap | Memperkirakan | Bahasa Inggris | nilai p |
|---|---|---|---|---|---|
| Panjang tarsus (mm) | |||||
| Identitas pasangan | 0.400 | Mencegat | 39.461 | 0,145 | <0,001 |
| Pola makan (buruk) | -2,048 | 0.152 | <0,001 | ||
| Tahun Penetasan (2020) | 0.114 | 0.207 | 0,583 | ||
| Usia kematangan (hari) | |||||
| Identitas pasangan | 24.820 | Mencegat | 52.246 | 1.348 | <0,001 |
| Pola makan (buruk) | 22.652 | 1.529 | <0,001 | ||
| Tahun Penetasan (2020) | 0.920 | 1.904 | 0.630 | ||
| Umur (hari) | |||||
| Identitas pasangan | 8852 | Mencegat | 727.680 | 31.450 | <0,001 |
| Pola makan (buruk) | -38.700 | 37.300 | 0.301 | ||
| Tahun Penetasan (2020) | -128.570 | 44.550 | 0,005 | ||
Pertumbuhan massa tubuh paling baik dijelaskan oleh fungsi Gompertz (Tabel S1 ), yang menunjukkan bahwa konstanta pertumbuhan intrinsik (k) betina dengan pola makan buruk sekitar 37% lebih rendah, dan usia puncak pertumbuhan ( t i ) sekitar 75% lebih lambat dibandingkan dengan betina dengan pola makan standar (Gambar S2 , Tabel S2 ). Massa tubuh asimtotik (A) diperkirakan hanya sekitar 2% lebih rendah untuk betina dengan pola makan buruk (Tabel S2 ), tetapi asimtot untuk betina ini tidak ditangkap dengan sempurna oleh fungsi Gompertz (Gambar S2 ).
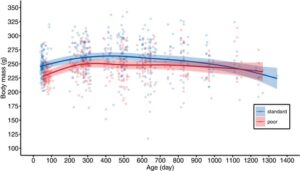
Memang, secara keseluruhan massa tubuh dewasa betina dengan pola makan buruk diperkirakan sekitar 5% lebih rendah daripada betina dengan pola makan standar, dengan perbedaan ini tetap terdeteksi hingga sekitar 2,5 tahun (Gambar 3 , Gambar S3 , Tabel 1 ). Setelah 2,5 tahun, perbedaan massa tubuh menghilang, tetapi ini kemungkinan besar karena hanya sedikit betina yang mencapai usia tersebut, yang menyebabkan peningkatan 95% CI di usia lanjut (Gambar 3 , Gambar S3 ). Tidak ada mortalitas selektif berkenaan dengan massa tubuh, karena rentang hidup dan massa tubuh dewasa tidak berhubungan (Tabel 1 ).
3.2 Reproduksi
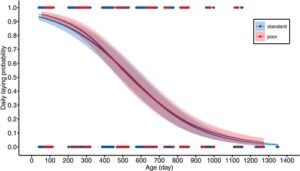
Permulaan bertelur tertunda sekitar 23 hari pada betina yang menerima diet pemeliharaan yang buruk (sekitar 40% kemudian, Gambar 2b , Tabel 2 ). Namun, tingkat bertelur tidak terpengaruh oleh diet pemeliharaan (Gambar 4 , Gambar S4 , Tabel 3 ). Betina dari kedua perlakuan diet mulai bertelur hampir satu butir per hari, tetapi cepat menua, dengan betina berusia tiga tahun hampir tidak bertelur lagi (Gambar 4 ). Penurunan ini tidak tergantung pada diet pemeliharaan (Gambar 4 , Gambar S4 , Tabel 3 ). Rentang hidup berhubungan positif dengan tingkat bertelur, yang menunjukkan bahwa betina yang paling subur juga yang paling dapat hidup (Tabel 3 ).
| Kemungkinan bertelur setiap hari | |||
|---|---|---|---|
| Koefisien parametrik | Memperkirakan | Bahasa Inggris | nilai p |
| Mencegat | 0.615 | 0,149 | <0,001 |
| Pola makan (buruk) | 0,168 | 0.181 | 0.353 |
| Tahun penetasan (2020) | 0,057 tahun | 0.214 | 0.790 |
| Jangka hidup | 0,001 | 0,001 | 0,003 |
| Istilah halus | Bahasa Inggris | Chi. persegi | nilai p |
| Usia: Diet (standar) | 1.002 | 573.1 | <0,001 |
| Usia: Pola makan (buruk) | 2.443 | 479.6 | <0,001 |
| Efek acak | Varians (SD) | 95% CI | nilai p |
| Identitas pasangan | 0,147 tahun | -0,236–0,530 | 0.703 |
| Identitas individu | 1.300 | 0,969–1,340 | <0,001 |
| Massa telur (g) | |||
|---|---|---|---|
| Koefisien parametrik | Memperkirakan | Bahasa Inggris | nilai p |
| Mencegat | 11.578 | 0.113 | <0,001 |
| Pola makan (buruk) | -0,191 | 0,090 | 0,033 |
| Tahun penetasan (2020) | -0,101 | 0,168 | 0,547 tahun |
| Jangka hidup | -0,001 | 0,001 | 0.256 |
| Istilah halus | Bahasa Inggris | Nilai F | nilai p |
| Usia: Diet (standar) | 8.487 | 307.78 | <0,001 |
| Usia: Pola makan (buruk) | 7.360 | 162.40 | <0,001 |
| Efek acak | Varians (SD) | 95% CI | nilai p |
| Identitas pasangan | 0.463 | 0,356–0,580 | <0,001 |
| Identitas individu | 0,267 tahun | 0,146–0,498 | 0,003 |
| Sisa | 0,529 | 0,328–0,745 |
| Massa telur (g) (dikoreksi untuk massa tubuh orang dewasa) | |||
|---|---|---|---|
| Koefisien parametrik | Memperkirakan | Bahasa Inggris | nilai p |
| Mencegat | 11.497 | 0,105 | <0,001 |
| Pola makan (buruk) | -0,035 | 0,087 tahun | 0,687 tahun |
| Tahun penetasan (2020) | -0,075 | 0,155 | 0.628 |
| Jangka hidup | -0,002 | 0,001 | 0.198 |
| Istilah halus | Bahasa Inggris | Nilai F | nilai p |
| Usia: Diet (standar) | 8.287 | 151.58 | <0,001 |
| Usia: Pola makan (buruk) | 7.191 | 62.96 | <0,001 |
| Massa tubuh | 5.216 | 13.85 | <0,001 |
| Efek acak | Varians (SD) | 95% CI | nilai p |
| Identitas pasangan | 0.383 | 0,219–0,503 | <0,001 |
| Identitas individu | 0.239 | 0,121–0,368 | <0,001 |
| Sisa | 0.521 | 0,405–0,739 |
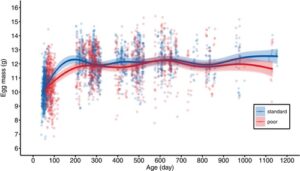
Betina yang menerima diet pemeliharaan yang buruk memang bertelur lebih kecil (Gambar 5 , Tabel 3 ). Efek ini tidak hadir pada minggu pertama bertelur, ketika semua betina masih bertelur relatif kecil, tetapi muncul pada usia selanjutnya, ketika betina telah mencapai ukuran telur puncaknya (Gambar 5 , Gambar S5 ). Ukuran telur tetap relatif konstan setelah puncaknya dan tidak terkait dengan umur (Gambar 5 , Tabel 3 ). Efek diet pada ukuran telur menghilang setelah dikoreksi dengan massa tubuh dewasa selama bertelur (Tabel 3 ), yang menunjukkan bahwa efek diet pada ukuran telur tidak independen dari efeknya pada ukuran tubuh dewasa.
3.3 Kelangsungan Hidup
Betina hidup, rata-rata, selama ca. 2 tahun, dengan kelompok tahun 2020 hidup lebih pendek daripada kelompok tahun 2019 (ca. 17% lebih pendek, Tabel 2 ). Diet pemeliharaan tidak berpengaruh pada harapan hidup (Gambar 2c , Tabel 2 ). Ini dikonfirmasi dengan analisis tingkat mortalitas spesifik usia mereka (Gambar 6 dan 7 , Tabel 4 ). Mortalitas meningkat seiring bertambahnya usia, tetapi ini tidak tergantung pada diet pemeliharaan (Gambar 7 , Gambar S6 , Tabel 4 ). Selain itu, ketika mortalitas dewasa dimodelkan dengan fungsi Weibull, tidak ada indikasi bahwa diet pemeliharaan memiliki efek pada mortalitas, karena interval kepercayaan 95% untuk efek diet pada parameter skala (mortalitas dasar) dan bentuk (mortalitas spesifik usia) tumpang tindih dengan nol (Tabel S3 ).
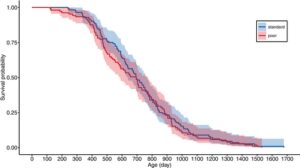
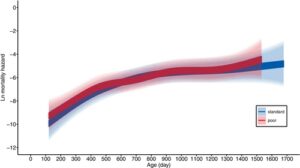
| Bahaya kematian orang dewasa | |||
|---|---|---|---|
| Koefisien parametrik | Memperkirakan | Bahasa Inggris | nilai p |
| Mencegat | -6.639 | 0,146 tahun | <0,001 |
| Pola makan (buruk) | 0.192 | 0,168 | 0.254 |
| Tahun penetasan (2020) | 0,535 | 0.183 | 0,003 |
| Istilah halus | Bahasa Inggris | Chi. persegi | nilai p |
| Usia: Diet (standar) | 3.711 | 88.08 | <0,001 |
| Usia: Pola makan (buruk) | 4.131 | 80.29 | <0,001 |
| Efek acak | Varians (SD) | 95% CI | nilai p |
| Identitas pasangan | 0.171 | 0,093–0,313 | 0,027 |
4 DISKUSI
Lingkungan yang buruk selama perkembangan individu umumnya dianggap memiliki efek negatif jangka panjang pada kinerja di masa dewasa (misalnya, Cooper & Kruuk, 2018 ; Hamel et al., 2009 ; Hayward et al., 2013 ; Lummaa & Clutton-Brock, 2002 ; Nussey et al., 2007 ; Reid et al., 2003 ; van de Pol et al., 2006 ; Victora et al., 2008 ). Namun, keberadaan efek ‘sendok perak’ tersebut di mana-mana (Grafen, 1988 ) dipertanyakan, karena tidak adanya efek jangka panjang juga sering dilaporkan (Cam et al., 2003 ; Drummond & Rodríguez, 2013 ; Vedder, Bichet, et al., 2022 ; Vedder & Bouwhuis, 2018 ; Wilkin & Sheldon, 2009 ). Pengetahuan lebih lanjut tentang jalur kausal yang mengarah pada efek ‘sendok perak’ dapat membantu menjelaskan mengapa kemunculan efek tersebut bervariasi antara penelitian. Dalam penelitian ini, kami melakukan uji coba terkontrol acak untuk menguji efek kausal dari diet pemeliharaan yang buruk terhadap kinerja pertumbuhan, reproduksi, dan kelangsungan hidup, selama seluruh perjalanan hidup burung puyuh Jepang betina. Kami secara khusus menampung semua betina dalam kondisi standar sebagai orang dewasa, untuk menguji efek lingkungan perkembangan yang independen dari efek lingkungan dewasa.
Meskipun pola makan yang buruk menyebabkan pertumbuhan yang sangat berkurang dan mengakibatkan ukuran tubuh yang mengecil sepanjang masa dewasa, tidak ada efek yang terdeteksi pada reproduksi (bertelur) dan kelangsungan hidup berdasarkan usia sepanjang masa hidup mereka. Satu-satunya efek perlakuan pola makan yang penting pada komponen ‘kebugaran’ pada spesies burung peliharaan ini, adalah keterlambatan permulaan reproduksi pada betina yang dipelihara dengan pola makan yang buruk. Rata-rata, betina ini mulai bertelur 23 hari lebih lambat daripada betina yang dipelihara dengan pola makan standar (Tabel 2 ). Mempertimbangkan bahwa tingkat bertelur pada burung puyuh Jepang dimulai tinggi dengan hampir satu telur per hari (Gambar 4 , Gambar S7 ), ini setara dengan bertelur sekitar 21 butir lebih sedikit dalam periode ini. Karena jumlah telur seumur hidup yang dikeluarkan betina rata-rata dapat diperkirakan sebesar 395 (lihat Informasi Pendukung Daring ), keterlambatan permulaan reproduksi ini sudah merupakan pengurangan sekitar 5% dalam ‘kebugaran’. Hal ini menggambarkan seleksi untuk performa di awal kehidupan pada spesies model burung ini, dan kesesuaiannya untuk mempelajari secara eksperimental proses eko-evolusi yang dapat memengaruhi performa selama rentang hidup lengkap, dalam jangka waktu yang dapat diperkirakan, meskipun tidak ada spesies burung liar yang dapat bertelur sebanyak itu.
Meskipun tidak adanya efek lain pada ‘kebugaran’, hasil bahwa betina yang dibesarkan dengan pola makan yang buruk tidak dapat sepenuhnya mengejar ukuran dan massa tubuh di masa dewasa menunjukkan bahwa efek ‘sendok perak’ dapat disebabkan oleh lingkaran umpan balik positif, di mana keuntungan ukuran dalam persaingan untuk sumber daya akan mengarah pada penguatan positif dalam akses ke sumber daya (Fokkema et al., 2021 ). Memang, jika efek ‘sendok perak’ muncul terutama karena daya saing yang berkurang, atau daya pegang sumber daya (Parker, 1974 ), tidak adanya efek pada reproduksi dan kelangsungan hidup orang dewasa dalam kondisi dewasa yang jinak sesuai dengan harapan. Sementara penelitian pada hewan di alam liar telah menunjukkan ini sebagai jalur yang mungkin di balik efek ‘sendok perak’ (Plard et al., 2015 ; van de Pol et al., 2006 ; Verhulst et al., 1997 ), penelitian tersebut tidak dapat mengesampingkan mekanisme tambahan yang bertanggung jawab atas efek ‘sendok perak’. Misalnya, jika individu mempertahankan pertumbuhan dan kelangsungan hidup dalam kondisi yang buruk dengan mengorbankan alokasi untuk perbaikan DNA (misalnya, Lemaître et al., 2015 ; Metcalfe & Monaghan, 2001 ), kinerja yang berkurang di akhir kehidupan juga harus terlihat dalam kondisi dewasa yang terstandarisasi. Namun, karena kami tidak menemukan bukti peningkatan penuaan dalam kinerja reproduksi dan kelangsungan hidup pada betina yang dibesarkan dengan pola makan yang buruk, efek tertunda yang diakibatkan oleh trade-off dalam individu mungkin relatif kecil.
Demikian pula, hipotesis ‘respons adaptif prediktif’ menyatakan bahwa efek jangka panjang yang negatif dapat terjadi karena ketidaksesuaian antara lingkungan perkembangan dan lingkungan dewasa, karena selama perkembangan individu mungkin disiapkan untuk lingkungan masa depan yang sesuai dengan lingkungan perkembangan (Gluckman & Hanson, 2004 ; Monaghan, 2008 ). Karena kondisi agar mekanisme tersebut adaptif, dan dengan demikian mampu berevolusi, relatif ketat (misalnya, Nettle et al., 2013 ; Rickard & Lummaa, 2007 ; Uller et al., 2013 ; Wells, 2007 ), kami tidak secara khusus menguji hipotesis ini dengan dua lingkungan dewasa yang berbeda. Meskipun demikian, diet unggas dewasa standar kami kemungkinan besar akan lebih cocok dengan diet anak ayam standar dan seharusnya menyebabkan penurunan kinerja dewasa untuk betina yang dibesarkan dengan diet yang buruk, jika mekanisme ‘respons adaptif prediktif’ telah bertindak. Fakta bahwa kami tidak menemukan bukti terjadinya hal ini dapat dijelaskan oleh efek yang lebih besar dari kualitas lingkungan saat ini. Akan tetapi, kami tidak dapat mengesampingkan bahwa ukuran tubuh yang lebih kecil saat dewasa akan menghasilkan kinerja yang lebih baik di lingkungan yang buruk saat dewasa dengan menurunkan kebutuhan makanan, dengan ukuran tubuh yang kecil bertindak sebagai ‘respons adaptif prediktif’ terhadap ketersediaan makanan yang buruk di masa dewasa (Monaghan, 2008 ).
Pentingnya lingkungan saat ini juga ditunjukkan oleh perbedaan kelangsungan hidup dewasa antara kedua kelompok. Hal ini kemungkinan besar disebabkan oleh efek lingkungan saat ini, karena kedua kelompok dibesarkan secara setara, tetapi sebagai orang dewasa yang ditempatkan di kandang burung luar ruangan yang relatif tidak terlindungi dari fluktuasi suhu sekitar. Sebagai alternatif dari hipotesis ‘respons adaptif prediktif’, telah dilaporkan bahwa efek ‘sendok perak’ diperburuk dalam lingkungan dewasa yang buruk (Briga et al., 2017 ), setiap efek ‘sendok perak’ yang tidak disebabkan oleh korelasi positif antara kualitas lingkungan perkembangan dan dewasa bisa jadi lebih kuat dalam kelompok kami dengan rentang hidup terpendek. Namun, analisis post hoc yang menguji efek interaksi antara perlakuan diet dan tahun penetasan pada rentang hidup tidak mendeteksi efek tersebut ( p = 0,696, Tabel S4 ). Oleh karena itu, terlepas dari kondisi lingkungan dewasa yang tepat, pendekatan eksperimental kami tidak memberikan bukti untuk efek jangka panjang yang dapat dideteksi dari lingkungan perkembangan yang buruk itu sendiri.
Karena ukuran betina yang lebih kecil yang dibesarkan dengan pola makan yang buruk juga menyebabkan mereka bertelur lebih kecil di masa dewasa, hasil kami memberikan bukti adanya dampak lingkungan perkembangan induk terhadap lingkungan perkembangan embrio keturunannya. Ketika dampak positif berlanjut ke semua tahap kehidupan keturunan berikutnya (seperti dibahas di atas), ini dapat menyebabkan dampak transgenerasional berjenjang dengan dampak dari generasi ke generasi (Aiken et al., 2015 ; Pick et al., 2019 ). Namun, potensi dampak transgenerasional tersebut kemungkinan besar dimodulasi oleh lingkungan. Penelitian sebelumnya pada burung puyuh Jepang menunjukkan korelasi yang kuat antara ukuran telur dan ukuran keturunan saat menetas, tetapi dampak ukuran telur digantikan oleh efek genetik dan lingkungan saat ini dengan cepat setelahnya, ketika anak ayam tumbuh dan makanan disediakan sepuasnya (Vedder et al., 2023 ). Di alam liar, keunggulan ukuran awal tersebut kemungkinan besar dapat dipertahankan dengan meningkatnya persaingan untuk mendapatkan makanan (Fokkema et al., 2021 ; Krist, 2011 ; Oddie, 2000 ).
Singkatnya, kami tidak menemukan bukti adanya efek jangka panjang dari lingkungan perkembangan yang buruk terhadap kelangsungan hidup dan reproduksi orang dewasa. Akan tetapi, karena kami menemukan efek yang terus berlanjut pada ukuran orang dewasa dan ukuran telur, kami tidak dapat mengesampingkan bahwa di alam liar, keunggulan ukuran dalam persaingan untuk mendapatkan sumber daya dapat menciptakan korelasi positif antara kualitas lingkungan dalam berbagai tahap kehidupan. Hal ini akan menyebabkan efek ‘sendok perak’, meskipun kualitas lingkungan saat ini merupakan faktor lingkungan yang paling penting dalam menentukan kinerja individu. Sebaliknya, ukuran yang lebih kecil dapat menyebabkan kebutuhan makanan yang lebih rendah, sehingga bertindak sebagai ‘respons adaptif prediktif’ terhadap lingkungan yang buruk.


