Abstrak
Studi ini menyelidiki pengaruh inulin dalam makanan terhadap pertumbuhan dan kesehatan udang muda Litopenaeus vannamei selama uji coba 8 minggu, dengan fokus pada pertumbuhan, kesehatan, dan dampak metabolik karena variasi makanan. Sebanyak 240 udang dengan berat rata-rata awal 0,26 ± 0,01 g didistribusikan ke 12 tangki resirkulasi dan diberi makanan dengan berbagai konsentrasi inulin: 0% (Kontrol), 0,5% (T1), 1,0% (T2) dan 1,5% (T3). Hasil penelitian menunjukkan bahwa makanan inulin 1,0% menghasilkan laju pertumbuhan spesifik, pertambahan berat, dan tingkat kelangsungan hidup tertinggi. Peningkatan aktivitas enzim antioksidan dan imun menunjukkan peningkatan kondisi kesehatan pada kadar inulin optimal, dibuktikan dengan penurunan kandungan malondialdehid dan potensiasi enzim seperti superoksida dismutase dan lisozim. Analisis mikrobioma melalui sequencing 16S rRNA menunjukkan pergeseran signifikan dalam populasi bakteri usus, dengan peningkatan bakteri menguntungkan seperti Xanthomarina dan pengurangan Vibrio dan Flavobacteria patogen dalam kelompok yang diberi suplemen inulin. Inulin makanan yang lebih tinggi meningkatkan keragaman mikroflora usus secara signifikan pada kelompok T2. Analisis metabolomik menunjukkan pengayaan jalur yang berbeda dan regulasi metabolit diferensial, yang menghubungkan asupan inulin dengan perubahan signifikan dalam metabolisme lipid, termasuk jalur seperti sekresi empedu, metabolisme asam arakidonat, biosintesis hormon steroid, dan transporter ABC. Lebih jauh lagi, analisis korelasi dan pengayaan KEGG mengungkapkan interaksi rumit dalam jaringan metabolisme udang, yang menunjukkan mekanisme kompensasi dan regulasi dalam menanggapi suplementasi inulin. Sebagai kesimpulan, inulin makanan secara substansial meningkatkan kesehatan dan pertumbuhan L. vannamei , kemungkinan dengan memodulasi mikrobiota usus dan jalur metabolisme. Studi ini menyoroti potensi inulin sebagai suplemen makanan yang signifikan untuk meningkatkan kemanjuran akuakultur udang.
1. PENDAHULUAN
Udang putih Pasifik ( Litopenaeus vannamei ) adalah salah satu spesies udang terpenting dalam akuakultur global, dengan nilai ekonomi yang signifikan (Venugopal & Gopakumar, 2017 ; Xie et al., 2019 ; Yu et al., 2019 ; Zhang et al., 2014 ). Suhu merupakan faktor lingkungan penting yang memengaruhi pertumbuhan dan kelangsungan hidup udang dalam ekosistem perairan (Cottin et al., 2016 ; Estrada-Cárdenas et al., 2021 ; Peng et al., 2015 ). Meskipun budidaya L. vannamei tersebar luas di wilayah tropis dan subtropis, stres suhu tetap menjadi tantangan yang signifikan, terutama di daerah dengan fluktuasi suhu yang besar. Stres suhu dapat berdampak signifikan pada fungsi fisiologis udang, termasuk berkurangnya kekebalan tubuh, laju pertumbuhan yang lebih lambat, dan meningkatnya kematian (Abdelrahman et al., 2019 ; Liu et al., 2019 ; Wang et al., 2019 ). Respons stres ini tidak hanya memengaruhi kesehatan udang tetapi juga berdampak buruk pada efisiensi ekonomi industri akuakultur. Oleh karena itu, menemukan metode yang efektif untuk mengurangi dampak stres suhu tinggi pada L. vannamei telah menjadi titik fokus penelitian saat ini.
Dalam beberapa tahun terakhir, telah terjadi peningkatan minat dalam penggunaan prebiotik sebagai aditif pakan dalam akuakultur. Prebiotik merujuk pada senyawa, substrat, gula rantai panjang, nutrisi, atau serat yang berfungsi sebagai makanan bagi mikroorganisme yang bermanfaat dalam sistem pencernaan inang (Davani-Davari et al., 2019 ). Prebiotik telah menunjukkan berbagai efek menguntungkan ketika digunakan sebagai aditif pakan, termasuk peningkatan pertumbuhan (Li et al., 2019 ; Ramos et al., 2017 ), peningkatan respons imun (Li et al., 2021 ; Selim & Reda, 2015 ), peningkatan resistensi penyakit (Abdel-Latif et al., 2022 ; Li, Liu, et al., 2018 ; Li, Xu, et al., 2018 ; Ylmaz et al., 2022 ), dan mitigasi stres abiotik pada spesies akuakultur (Hoseinifar et al., 2014 ). Inulin memainkan peran penting dalam modulasi mikrobiota usus, yang penting untuk menjaga kesehatan udang dan memperkuat resistensi terhadap patogen (Li, Liu, et al., 2018 ; Li, Xu, et al., 2018 ). Lebih jauh lagi, penelitian telah menunjukkan bahwa inulin, ketika disertakan dalam pakan pada tingkat mulai dari 0,25% hingga 2%, menunjukkan sifat antimikroba dan imunomodulatori pada berbagai spesies akuatik, termasuk ikan mas rumput, salmon Atlantik, dan nila (Gibson et al., 2010 ; Ringø et al., 2010 ; Yukgehnaish et al., 2020 ). Mengingat efek buruk stres suhu pada udang putih Pasifik, mengeksplorasi potensi inulin sebagai aditif prebiotik untuk meringankan efek ini menjadi sangat penting.
Untuk memberikan pemahaman yang komprehensif tentang potensi inulin dalam mengurangi stres suhu, penelitian ini tidak hanya akan menilai dampak aditif pakan inulin pada pertumbuhan, kelangsungan hidup, dan status kesehatan udang putih Pasifik dalam kondisi suhu tinggi, tetapi juga mengintegrasikan pengurutan mikrobiota usus dan profil metabolomik yang komprehensif untuk analisis gabungan. Penelitian ini bertujuan untuk mengevaluasi dampak aditif pakan inulin pada pertumbuhan, kelangsungan hidup, dan status kesehatan udang putih Pasifik dalam kondisi stres suhu tinggi, yang bertujuan untuk menawarkan strategi yang efektif untuk akuakultur udang. Penelitian ini bertujuan untuk memberikan landasan teoritis yang penting dan panduan praktis untuk mengoptimalkan pengelolaan akuakultur udang dan meningkatkan keuntungan akuakultur.
2 BAHAN DAN METODE
2.1 Desain Eksperimen
Kami memperoleh 240 udang muda yang sehat dari Hongfeng Aquaculture, Zhousan Ltd., yang berlokasi di Zhoushan, provinsi Zhejiang, Tiongkok. Udang-udang muda ini, dengan ukuran rata-rata (0,26 ± 0,02 g, 0,42 ± 0,07 cm), dialokasikan secara acak ke dalam 12 tangki plastik (100 L), dengan 20 udang per tangki. Pemberian pakan dilakukan tiga kali sehari pada pukul 7:00, 13:00, dan 19:00, berlanjut selama 8 minggu. Sebelum setiap sesi pemberian pakan, sisa atau pakan yang tidak dimakan, feses, dan bulu dikeluarkan dari tangki pada pagi dan sore hari. Pemantauan harian parameter kualitas air, termasuk suhu, oksigen terlarut, dan pH, dilakukan menggunakan pemeriksa kualitas air HQ40d (perusahaan HACH, Loveland, AS). Kadar amonia dan nitrit dinilai selama uji pemberian pakan menggunakan metode spektrofotometri. Suhu air dipertahankan pada 32 ± 1°C, kadar oksigen terlarut antara 6,3 dan 7,4 mg L −1 , pH pada 8,1 ± 0,2, salinitas pada 21‰, total amonia–nitrogen pada 0,05 mg L −1 , dan konsentrasi nitrit di bawah 0,02 mg L −1 .
2.2 Persiapan dan pemberian pakan
Penyiapan pakan udang mengacu pada Gao et al. Inulin diperoleh dari Shenzhen IWOWN Technology Co., Ltd. Pakan Eksperimen disiapkan pada gradien konsentrasi 0% (kontrol), 0,5% (T 1 ), 1,0% (T 2 ), dan 1,5% (T 3 ), berturut-turut. Formulasi dan perkiraan komposisi pakan disediakan dalam Tabel 1 . Selama percobaan, udang diberi makan tiga kali sehari (pukul 7:00, 13:00, dan 19:00), dengan takaran 5%–7% dari berat badan basahnya, selama 8 minggu. Pompa air sirkulasi dihentikan selama 1 jam selama pemberian pakan. Setelah 1 jam, sisa pakan dibuang, dan jumlah pemberian disesuaikan sedikit di atas rasa kenyang berdasarkan jumlah pakan yang tidak dimakan di dalam tangki.
| Bahan-bahan | Kelompok perawatan | |||
|---|---|---|---|---|
| angka 0 | 0,5% | 1,0% dari | 1,5% | |
| Bahan-bahan (%) | ||||
| Tepung ikan | 36 | 36 | 36 | 36 |
| Bungkil kedelai | 20 | 20 | 20 | 20 |
| Bubuk ragi | 4 | 4 | 4 | 4 |
| Lesitin kedelai | 6 | 6 | 6 | 6 |
| Tepung rumput laut | 2.5 | 2.5 | 2.5 | 2.5 |
| Minyak ikan | 6.5 | 6.5 | 6.5 | 6.5 |
| Monokalsium fosfat | 2.5 | 2.5 | 2.5 | 2.5 |
| Campuran Vitamin | 2.6 | 2.6 | 2.6 | 2.6 |
| b Campuran mineral | 3 | 3 | 3 | 3 |
| 50% Kolin klorida | 3 | 3 | 3 | 3 |
| taurin | 1.9 | 1.9 | 1.9 | 1.9 |
| Tepung gandum | 12 | 11.5 | 11.0 | 10.5 |
| c.Inulin | angka 0 | 0.5 | 1.0 | 1.5 |
| Komposisi proksimal (% bahan kering) | ||||
| Protein kasar | 42.3 | 42.2 | 41.9 | 42.1 |
| Lipid kasar | 9.1 | 9.0 | 9.2 | 9.2 |
| Abu | 12.3 | 12.5 | 12.6 | 12.4 |
| Kalsium | 2.2 | 2.4 | 2.3 | 2.4 |
Premiks vitamin (kg −1 makanan): VA, 400.000 IU; VD, 40.000 IU; VB 2 , 0,5 g; VE, 30 g; VB 6 , 0,4 g; VB 12 , 0,012 g; VB 1 , 0,02 g; VK, 0,04 g; asam folat, 0,13 g; VH, 0,01 g; α-tokoferol, 3000 IU; mio-inositol, 8 g; kalsium pantotenat, 0,76 g; asam nikotinat, 0,2 g. b Campuran mineral (kg −1 makanan): ZnSO 4 · 7H 2 O, 10 g; FeSO4•7H 2 O, 20 g; MnSO 4 •H 2 O, 3 g; MgSO 4 •7H 2 O, 12 g; Ca(HPO 4 ) 2 •2H 2 O, 600 g; CuSO 4 •5H 2 O, 3 g; CaCO 3 , 215 g; KCL, 90 g; KI, 0,04 g; NaCl, 40 g. c Setiap campuran dibuat dengan menambah jumlah inulin dan mengurangi jumlah tepung gandum. Komposisi premiks inulin per kilogram dari diet 1 hingga 4 adalah sebagai berikut (g kg −1 premiks): inulin 0, 5, 10, dan 15 g; tepung gandum 120, 115, 110, dan 105 g.
2.3 Pengambilan sampel
Pada akhir uji coba pemberian pakan selama 8 minggu, udang diambil sampelnya setelah periode puasa selama 24 jam. Pengukuran panjang udang (FBL) dan berat (FBW) dilakukan, dan kinerja pertumbuhan serta tingkat kelangsungan hidup dihitung. Sampel hepatopankreas, usus, dan otot dari setiap kelompok dikumpulkan untuk analisis aktivitas enzim. Jaringan usus dari kelompok kontrol dan kelompok dengan kinerja pertumbuhan terbaik dikumpulkan untuk analisis metabolomik dan mikroflora usus. Semua jaringan yang dikumpulkan segera disimpan pada suhu -80°C hingga digunakan lebih lanjut.
Kinerja pertumbuhan dan tingkat kelangsungan hidup dihitung menggunakan persamaan berikut:

2.4 Uji biokimia
Aktivitas enzim dianalisis mengikuti prosedur Lv et al. ( 2021 ). Sampel hepatopankreas, usus, dan otot beku dihomogenisasi dalam 10 volume (v/w) larutan penyangga fosfat dingin (Procell Life Science & Technology Co., Ltd., Wuhan, Tiongkok), dan disentrifugasi pada 3000 g selama 20 menit pada suhu 4°C. Setelah menghilangkan endapan, supernatan segera digunakan untuk analisis aktivitas enzim antioksidan dan imun. Total kandungan protein dalam homogenat jaringan diukur dengan kit uji protein biru cemerlang Coomassie (Jiancheng, Ltd., Nanjing, Tiongkok) mengikuti protokol pabrik. Semua uji dijalankan dalam tiga sampel replikasi. Kandungan superoksida dismutase (SOD), katalase (CAT), malonaldehida (MDA), lisozim (LZM), alkali fosfatase (AKP), asam fosfatase (ACP), alanin aminotransferase (ALT) dan aspartat aminotransferase (AST) dalam supernatan dianalisis dengan kit enzim (Jiancheng, Ltd., Nanjing, Cina) mengikuti petunjuk produsen dan dibaca pada spektrofotometer (Bio-RAD, AS).
2.5 Analisis mikrobiota usus
Untuk menganalisis mikrobiota usus dalam penelitian ini, protokol berikut diadopsi, mengacu pada metode yang dijelaskan oleh Lv et al. ( 2021 ). Tiga udang digunakan per replikasi, dari mana usus dikeluarkan dengan hati-hati. Isi usus kemudian diekstraksi, dikumpulkan, dan disimpan dalam tabung sentrifus polipropilena steril pada suhu −80°C untuk ekstraksi DNA berikutnya. DNA mikroba diisolasi dari sampel-sampel ini menggunakan EZNA Soil DNA Kit (D3396-01; Omega Bio-tek, Inc., AS), mengikuti pedoman produsen. Kuantifikasi konsentrasi DNA dicapai dengan menggunakan fluorometer Qubit 2.0 (Invitrogen, AS), dan penilaian integritas DNA dilakukan melalui elektroforesis gel agarosa 2%. Evaluasi lebih lanjut dari konsentrasi DNA menggunakan NanoDrop (ND-2000, Thermo Fisher Scientific, Tiongkok). DNA yang diekstraksi diawetkan pada suhu −80°C hingga pemrosesan lebih lanjut. Untuk amplifikasi PCR, Trans Start FastPfu DNA Polymerase (TransGen Biotech, China) digunakan dalam volume reaksi 20 μL, yang terdiri dari 4 μL buffer FastPfu 5x, 2,0 μL dNTP (masing-masing 2,5 mM), 0,8 μL setiap primer (5 μM), 0,4 μL FastPfu Polymerase, dan 10 ng DNA cetakan. Protokol PCR melibatkan denaturasi awal pada suhu 95°C selama 5 menit, diikuti oleh 27 siklus pada suhu 95°C selama 30 detik, 55°C selama 30 detik, dan 72°C selama 45 detik, diakhiri dengan ekstensi akhir pada suhu 72°C selama 10 menit. Ini dilakukan pada siklus termal ABI GeneAmp 9700 (Applied Biosystems, AS). DNA yang diperkuat diverifikasi menggunakan elektroforesis gel, dengan pita target dipotong dan diukur menggunakan kit uji DNA Qubit 2.0 (Invitrogen, AS). Produk PCR yang dimurnikan kemudian diurutkan pada platform Illumina HiSeq (Illumina, AS) oleh Origin-gene Bio-pharmaceutical Technology Co., Ltd., Tiongkok. Data sekuensing melibatkan pembacaan ujung berpasangan yang disusun menurut kode batang unik, diproses dengan menghilangkan kode batang dan urutan primer, dan difilter kualitasnya menggunakan perangkat lunak Usearch. Urutan yang lolos pemeriksaan kualitas dikelompokkan ke dalam unit taksonomi operasional (OTU) menggunakan ambang batas kesamaan 97%. Klasifikasi taksonomi dilakukan dengan pengklasifikasi RDP pada tingkat kepercayaan 0,80. Analisis keanekaragaman mencakup penilaian keanekaragaman alfa dan beta. Keanekaragaman alfa dihitung dengan indeks seperti Chao1, ACE, Shannon, dan Simpson, menggunakan perangkat lunak QIIME, dan hasilnya divisualisasikan menggunakan perangkat lunak R. Keanekaragaman beta dieksplorasi melalui analisis komponen utama (PCA), yang menggunakan jarak Bray-Curtis dan Weighted Unifrac untuk memeriksa variasi antara komunitas mikroba. Taksa bakteri yang terwakili secara berbeda dikategorikan menggunakan analisis LEfSe.
2.6 Analisis metabolit
Analisis metabolom yang tidak ditargetkan mengikuti metode Lv et al. ( 2024 ). Ekstrak sampel dianalisis menggunakan sistem LC/MS (UPLC, ExionLC AD, https://sciex.com.cn/ ; MS, Sistem QTRAP®, https://sciex.com/ ) untuk analisis metabolik. Operasi analitis dilakukan pada kolom Waters ACQUITY UPLC HSS T3 C18, yang dijaga pada suhu stabil 40°C. Sistem beroperasi dengan laju alir 0,4 mL/menit dan volume injeksi 2 μL. Fase mobil adalah campuran gradien air dan asetonitril, masing-masing mengandung 0,1% asam format. Ionisasi elektrospray (ESI) dilakukan dalam mode ion positif dan negatif, dengan pengaturan yang dioptimalkan secara hati-hati untuk suhu sumber ion, tegangan semprotan ion, dan aliran gas. Pemrosesan data dan kuantifikasi metabolit difasilitasi oleh perangkat lunak Analyst versi 1.6.3. Penyaringan untuk metabolit diferensial (DM) dilakukan dengan kriteria yang ketat, diikuti oleh analisis pengayaan jalur KEGG. Selain itu, jaringan korelasi dibentuk antara gen yang diekspresikan secara diferensial (DEG) dan DM menggunakan koefisien korelasi Pearson.
2.7 Analisis Statistik
Analisis varians satu arah (ANOVA) dilakukan menggunakan perangkat lunak SPSS versi 26.0 untuk mengevaluasi perbedaan kinerja pertumbuhan dan aktivitas enzim di antara perlakuan. Perbandingan post hoc dilakukan dengan uji rentang berganda Duncan, dengan menetapkan ambang signifikansi pada p < 0,05. Data disajikan sebagai mean ± SD. Representasi grafis dibuat menggunakan Prism 8.0, dengan semua nilai dinyatakan sebagai mean ± SD.
3 HASIL
3.1 Kinerja pertumbuhan
Tabel 2 menunjukkan kinerja pertumbuhan udang setelah 8 minggu. Udang dalam kelompok diet 1,0% menunjukkan FBL yang secara signifikan lebih besar dibandingkan dengan kelompok kontrol dan kelompok 1,5%, meskipun tidak ada perbedaan yang signifikan dari kelompok 0,5%. FBW secara signifikan lebih tinggi dalam kelompok diet 0,5%, 1,0%, dan 1,5% dibandingkan dengan kelompok kontrol. Kelompok diet 1,0% memiliki WG yang secara signifikan lebih tinggi dibandingkan dengan kelompok kontrol, 0,5%, dan 1,5%. SGRL tidak bervariasi secara signifikan di seluruh kelompok diet pada inklusi 0% dan 0,5%. Namun, pada inklusi 1,0% dan 1,5%, SGRL meningkat secara signifikan. Demikian pula, SGRW menunjukkan tren naik dengan peningkatan inklusi makanan, yang menunjukkan peningkatan yang signifikan. Tingkat kelangsungan hidup juga meningkat secara signifikan dengan konsentrasi yang lebih tinggi, yang menunjukkan peningkatan yang jelas dalam kelompok inklusi 1,0% dan 1,5%.
| Luas Permukaan Tanah (cm) | FBW(g) | Kelompok Kerja (%) | SGRL (%) | SGRW (%) | Tingkat kelangsungan hidup (%) | |
|---|---|---|---|---|---|---|
| 0% (Kon.) | 7,48 ± 0,42 satuan | 2,67 ± 0,45 per menit | 611,23 ± 68,43 per tahun | 1,51 ± 0,10 per menit | 4,63 ± 0,19 per menit | 67,50 ± 6,11 per menit |
| 0,5% | 7,51 ± 0,84 per menit | 2,97 ± 0,34 miliar | 688,19 ± 58,98 per tahun | 1,52 ± 0,21 satuan | 4,81 ± 0,34 inci | 69,33 ± 6,33 per menit |
| 1,0% dari | 7,93 ± 0,51 miliar | 3,20 ± 0,27 miliar | 896,92 ± 106,90 miliar | 1,64 ± 0,14 miliar | 4,95 ± 0,24 miliar | 81,67 ± 4,62 miliar |
| 1,5% | 7,88 ± 0,39 per menit | 3,19 ± 0,22 miliar | 844,73 ± 87,97 miliar | 1,59 ± 0,09 ab | 4,94 ± 0,12 miliar | 77,67 ± 6,11 miliar |
Catatan : Nilai dinyatakan sebagai mean ± SD ( n = 3). Huruf yang berbeda pada baris yang sama menunjukkan perbedaan yang signifikan ( p < 0,05).
3.2 Aktivitas enzim
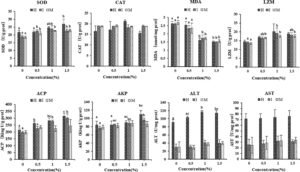
Aktivitas enzim SOD lebih tinggi di jaringan usus dan otot kelompok eksperimen daripada kontrol (Gambar 1 , p < 0,05), dengan kelompok 1,5% menunjukkan aktivitas tertinggi di hepatopankreas. Sebaliknya, aktivitas CAT tidak menunjukkan perbedaan signifikan antar kelompok ( p > 0,05). Kandungan MDA menunjukkan penurunan dengan meningkatnya konsentrasi inulin, menunjukkan pengurangan signifikan pada kelompok 1,0% dan 1,5% dibandingkan dengan kontrol ( p < 0,05). Aktivitas LZM awalnya meningkat dan kemudian menurun dengan konsentrasi inulin, dengan peningkatan signifikan diamati pada kelompok 0,5% dan 1,0% dibandingkan dengan kontrol ( p < 0,05). Baik aktivitas ACP maupun AKP meningkat dengan konsentrasi inulin, menunjukkan peningkatan signifikan pada kelompok 1,0% dan 1,5% relatif terhadap kontrol ( p < 0,05). Demikian pula, aktivitas ALT secara signifikan lebih tinggi pada kelompok inulin 1,0% dibandingkan pada kelompok kontrol ( p < 0,05). Aktivitas AST juga meningkat seiring dengan konsentrasi inulin, mencapai aktivitas puncaknya pada kelompok 1,5%, meskipun perubahan ini tidak mencapai signifikansi statistik ( p > 0,05).
3.3 Hasil mikrobiota usus
Indeks Chao1 dan ACE menunjukkan kekayaan spesies yang lebih tinggi dalam kelompok inulin dibandingkan dengan kelompok kontrol, seperti yang ditunjukkan pada Gambar 2a . Selain itu, indeks keanekaragaman Shannon dan Simpson mengungkapkan bahwa kelompok inulin juga menunjukkan homogenitas spesies yang lebih tinggi, yang menunjukkan keanekaragaman komunitas mikroba yang terdistribusi lebih merata dibandingkan dengan kontrol.
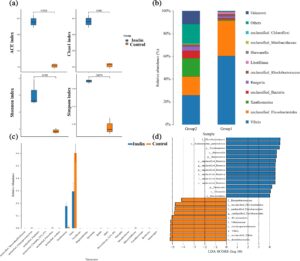
Gambar 2b menyajikan unit taksonomi operasional (OTU) yang paling umum untuk kedua kelompok, dengan klasifikasi tingkat genus yang berasal dari kelimpahan relatif bacaan yang ditetapkan. Pada kelompok kontrol, genus Vibrio dan anggota ordo Flavobacteriales yang tidak ditentukan adalah yang dominan. Sebaliknya, kelompok inulin dicirikan oleh komposisi genus yang bervariasi yang mencakup Vibrio, Flavobacteriales yang tidak ditentukan , Xanthomarina , dan Chloroflexi yang tidak ditentukan . Khususnya, genus Xanthomarina menunjukkan kelimpahan yang jauh lebih tinggi pada kelompok inulin dibandingkan dengan kontrol, seperti yang dikonfirmasi oleh analisis ANOVA (Gambar 2c ).
Perbedaan signifikan dalam populasi mikroba antara kedua kelompok lebih jauh digarisbawahi melalui analisis regresi linear discriminant analysis (LDA), yang dirinci dalam Gambar 2d . Dalam kelompok inulin, berbagai macam genera sangat berlimpah, termasuk Flavobacteriaceae , Xanthomarina gelatinilytica , Xanthomarina , Bacteroidia , Bacteroidota , Bacteria yang tidak ditentukan , Firmicutes , Clostridia , dan Bacteroidales . Sebagai perbandingan, kelompok kontrol menunjukkan kelimpahan relatif yang lebih tinggi dari Ilumatobacteraceae , anggota Flavobacteriales yang tidak ditentukan , Proteobacteria , Vibrionaceae , Gammaproteobacteria , Vibrio , Vibrio yang tidak ditentukan , dan Enterobacterales .
3.4 Hasil Metabolomik
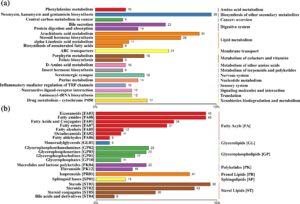
Hasil analisis pengayaan KEGG dari DM ditunjukkan pada Gambar 3 , yang menyajikan 20 jalur teratas dengan nilai q signifikan terendah . Jalur utama yang diperkaya secara signifikan meliputi sekresi empedu, metabolisme asam arakidonat, biosintesis hormon steroid, dan transporter ABC (Gambar 3a ). Kelompok lipid metilasi diferensial (DM) utama dikategorikan ke dalam beberapa kelas: sterol, steroid, dan konjugat steroid dikelompokkan di bawah lipid sterol. Eikosanoid, amida lemak, asam lemak, dan konjugatnya, bersama dengan ester lemak, diklasifikasikan sebagai asil lemak. Isoprenoid diidentifikasi dalam kategori lipid prenol, dan gliserofosfotanolamin, bersama dengan gliserofosfolipid, diklasifikasikan di bawah gliserofosfolipid (Gambar 3b ).
PCA berhasil membedakan antara profil metabolit dari kelompok kontrol dan kelompok yang diberi suplemen inulin, yang mencerminkan dampak signifikan inulin pada metabolisme udang (Gambar 4a ). Plot skor orthogonal projections to latent structures discriminant analysis (OPLS-DA) semakin menonjolkan perbedaan ini, dengan menunjukkan pengelompokan yang jelas sepanjang sumbu x, yang menguatkan bahwa inulin adalah pemberi pengaruh utama dalam variasi metabolik yang diamati antara kedua kelompok (Gambar 4b ). Selain itu, plot substitusi OPLS-DA mengonfirmasi keandalan model, dibuktikan dengan semua titik Q2 merah dalam plot uji substitusi yang tetap lebih rendah daripada titik Q2 merah asli dari kiri ke kanan, sehingga mengindikasikan tidak ada overfitting (Gambar 4c ).

Analisis lebih lanjut tentang perubahan metabolik disajikan dalam Gambar 5a , yang memberi peringkat metabolit berdasarkan Log2 Fold Change antara kelompok-kelompok tersebut. Khususnya, 10 metabolit teratas dengan peningkatan signifikan meliputi Lasinavir, Desglucocheirotoxin, PC (2:0/PGE1), dan lainnya. Sebaliknya, 10 metabolit teratas yang mengalami penurunan meliputi senyawa-senyawa seperti 1α3β,22R-Trihydroxyergosta-5,24E-dien-26-oic acid 3-O-β-D-glucoside 26-O-β-D-glucosyl ester dan oleoyltaurine, antara lain.

Plot gunung berapi menggambarkan pergeseran ini lebih lanjut, dengan mengilustrasikan bahwa dari 3745 metabolit yang berbeda secara signifikan, 2036 mengalami penurunan regulasi yang signifikan, 1709 mengalami peningkatan regulasi, dan 1539 tetap tidak berubah (Gambar 5b ). Lima metabolit kualitatif pertama adalah morfolin, DG (12:0/20:5(7Z, 9Z, 11E, 13E, 17Z)-3OH (5, 6, 15)/0:0), PC(P-18:0/PGE1), 1-heptadekanoil-sn-glisero-3-fosfokolin, dan Fusarokromanon.
Analisis korelasi di seluruh sampel menunjukkan bahwa sebagian besar koefisien korelasi mendekati +1 atau -1, yang menunjukkan bahwa kadar metabolit secara konsisten naik atau turun bersama atau menunjukkan hubungan terbalik yang kuat (Gambar 5c ). Koefisien negatif yang tinggi ini menunjukkan interaksi kompensasi atau regulasi di antara metabolit, di mana peningkatan atau keberadaan satu terkait langsung dengan pengurangan yang lain. Analisis ini menyoroti interaksi metabolik rumit yang dipengaruhi oleh inulin makanan pada udang, yang menunjukkan jalur regulasi dan kompensasi yang kompleks dalam jaringan metaboliknya. Secara khusus, Morpholine dan (S)-Nerolidol 3-O-[aL-Rhamnopyranosyl-(1- > 4)-aL-rhamnopyranosyl-(1- > 2)-[aL-rhamnopyranosyl-(1- > 6)]-bD-glucopyranoside] menunjukkan korelasi positif yang sangat tinggi (0,999979336), yang menunjukkan peran sinergis potensial atau jalur biosintesis bersama. Sebaliknya, Morpholine dan 1-heptadecanoyl-sn-glycero-3-phosphocholine berkorelasi negatif kuat (−0,999972675), yang menunjukkan kemungkinan interaksi antagonis atau kompetitif dalam proses metabolismenya. Fusarochromanone menunjukkan korelasi negatif yang sangat erat dengan hampir semua metabolit lainnya (sekitar −0,999989635), yang menunjukkan perannya yang berbeda atau dampak signifikannya dalam jalur metabolisme tertentu.
3.5 Analisis integrasi mikrobiota dan metabolom
Dalam kategori mikrobioma, data mencakup berbagai macam mikroorganisme, termasuk spesies yang tidak dikultur dan tidak terklasifikasi, serta bakteri yang diidentifikasi secara spesifik seperti Deinococcus , Devosia , dan Methanosarcina (Gambar 6a ). Mikroba khusus ini secara konsisten menunjukkan nilai yang sama dalam Pemuatan 2 (sekitar 0,178), yang menunjukkan bahwa mereka memiliki respons yang sama terhadap suplementasi inulin. Namun, nilai negatifnya dalam Pemuatan 1 dapat menunjukkan korelasi terbalik dengan komponen ini, yang mengisyaratkan interaksi mendasar yang kompleks. Data metabolom dicirikan oleh berbagai senyawa biokimia, termasuk zat kompleks seperti asam 3b,18b-3-Methoxy-11-oxo-12-oleanen-30-oic dan Glycerol 1-(5-hydroxydodecanoate). Metabolit ini menunjukkan variabilitas yang lebih besar baik dalam nilai pemuatan maupun panjang keseluruhannya. Variasi ini menunjukkan bahwa metabolit ini memiliki peran yang beragam dan berpartisipasi dalam berbagai interaksi dalam sistem biologis, yang mencerminkan sifat rumit fungsi dan ketergantungannya. Diagram kord pada Gambar 6b menyajikan pemeriksaan mikroflora yang komprehensif, termasuk strain tertentu seperti Rothia , Halocynthiibacter , dan Mesoflavibacter , serta klasifikasi yang lebih luas seperti Archaea yang tidak terklasifikasi dan Clostridia UCG_014 yang tidak terklasifikasi , yang menunjukkan spektrum penelitian bakteri yang luas. Metabolit yang dianalisis mencakup berbagai zat biokimia yang kompleks, seperti ester glukosa asam absisat, Lividomycin A, dan Isopentyl gentiobioside, yang menunjukkan minat yang luas pada berbagai jalur biokimia dan interaksinya dalam populasi mikroba ini. Mayoritas korelasi yang diamati bersifat positif, sering kali mendekati nilai 1, yang menunjukkan hubungan yang kuat dan langsung antara metabolit tertentu dan kelimpahan atau aktivitas mikroba. Namun, terdapat pengecualian penting, seperti korelasi negatif dengan Coagulin R 3-glukosa, yang menunjukkan bahwa ia mungkin memiliki efek penghambatan atau persaingan pada berbagai mikroorganisme. Selain itu, beberapa interaksi spesifik menonjol karena koefisien korelasinya yang tinggi, yang memperkuat signifikansi biologisnya. Ini termasuk hubungan yang sangat kuat antara Halocynthiibacter dan Annuolide C (CC: 0,997), Archaea yang tidak terklasifikasi dan 9-metoksi-3-metil Isoquinolin-7-on (CC: 0,994), dan Halocynthiibacter dan Daumone (CC: 0,996). Interaksi menarik lainnya melibatkan Mesoflavibacter dan PI(PGE2/22:3(10Z,13Z,16Z)) (CC: 0,991), yang menggarisbawahi relevansinya dalam studi metabolik.

4 DISKUSI
Inulin, fruktan dan serat larut yang banyak bersumber dari akar sawi putih dan tanaman lainnya, telah menunjukkan potensi yang besar sebagai bahan tambahan makanan dalam akuakultur, khususnya karena sifat prebiotiknya (Qin et al., 2023 ). Aplikasi inulin dalam akuakultur terkenal karena kontribusinya terhadap peningkatan kesehatan usus, peningkatan respons imun, dan peningkatan metrik pertumbuhan spesies akuatik (De La Cruz-Marín et al., 2023 ; Todorov et al., 2024 ; Zhang et al., 2023 ).
Peningkatan kinerja pertumbuhan dan parameter kesehatan pada udang yang diberi pakan tambahan inulin, seperti yang diamati dalam penelitian kami, selaras dengan badan penelitian yang lebih luas tentang penggunaan prebiotik dalam akuakultur. Berbagai penelitian juga melaporkan efek positif. Sebuah tinjauan oleh Ringø et al. ( 2010 ) telah menunjukkan bahwa inulin makanan menyebabkan peningkatan kinerja pertumbuhan pada berbagai spesies ikan dengan meningkatkan laju penyerapan nutrisi. Temuan kami menggemakan hasil ini, dengan udang mencatat berat dan panjang tubuh yang lebih tinggi dalam kelompok yang diobati dengan inulin. Mugwanya et al. (2022 ) juga mencatat hasil serupa dalam diet nila yang disuplemen dengan inulin, menggarisbawahi manfaat yang konsisten di seluruh spesies akuatik yang berbeda. Respons yang bergantung pada konsentrasi yang kami amati, khususnya dengan hasil optimal pada tingkat inklusi inulin 1,0%, selaras dengan Zhong et al. ( 2020 ), yang menemukan bahwa kadar serat makanan sedang dapat mengoptimalkan kapasitas pemacu pertumbuhan largemouth bass ( Micropterus salmoides ) tanpa menyebabkan penyerapan nutrisi yang tidak efisien. Analisis imunologi dari percobaan kami menunjukkan peningkatan aktivitas beberapa enzim terkait imun di jaringan udang ketika diberi makan inulin. Khususnya, aktivitas SOD secara signifikan lebih tinggi di usus dan otot, dan aktivitas lanjutan dari asam fosfatase (ACP) dan alkali fosfatase (AKP) diamati dengan meningkatnya konsentrasi inulin. Enzim-enzim ini memainkan peran penting dalam garis pertahanan pertama melawan patogen dengan mengatur stres oksidatif dan meningkatkan respons imun secara keseluruhan (Iddir et al., 2020 ). Pengamatan kami terhadap peningkatan aktivitas enzim SOD dan ACP dengan suplementasi inulin memiliki kesamaan dengan temuan oleh Butt et al. ( 2021 ), yang melaporkan peningkatan aktivitas enzim imun pada udang yang terpapar prebiotik makanan termasuk manan-oligosakarida dan β-glukan. Namun, penelitian kami secara khusus menyoroti peningkatan aktivitas SOD selektif inulin tanpa memengaruhi aktivitas CAT, yang menunjukkan modulasi imun yang lebih bernuansa dibandingkan dengan efek spektrum yang lebih luas yang dicatat oleh Hoseinifar et al. ( 2015 ), di mana beberapa enzim antioksidan meningkat pada ikan mas ( Cyprinus carpio ) yang diberi prebiotik. Perubahan positif dalam aktivitas enzim meningkatkan proses metabolisme dan mendorong respons imun yang lebih baik, yang mendukung peningkatan pertumbuhan yang diamati.
Meta-analisis mengenai efek inulin dalam makanan terhadap profil metabolik dan mikroba pada udang menggarisbawahi dampak yang beragam, meningkatkan pemahaman kita tentang bagaimana intervensi makanan dapat memengaruhi spesies akuakultur pada tingkat biokimia dan mikroba. Integrasi temuan kami dengan temuan dari literatur penelitian yang lebih luas mengungkapkan hubungan yang mendalam antara suplementasi prebiotik, modulasi metabolik, dan peningkatan ekosistem mikroba.
Temuan kami menunjukkan komunitas mikroba yang beragam pada udang yang diberi makan inulin, yang mencakup pengurangan yang nyata pada bakteri berbahaya seperti Vibrio dan peningkatan pada bakteri menguntungkan seperti Xanthomarina . Dengan mendorong lingkungan yang kompetitif yang tidak mendukung bakteri patogen, inulin dapat mengubah populasi mikroba secara signifikan, memperkaya mereka yang berkontribusi pada lingkungan usus yang lebih sehat (Wee et al., 2024 ). Perubahan tersebut cenderung meningkatkan resistensi penyakit dan efisiensi pemrosesan nutrisi, yang selanjutnya memengaruhi kinerja pertumbuhan secara menguntungkan. Ini mendukung pernyataan yang dibuat oleh Holt et al. ( 2021 ), yang menunjukkan bahwa inulin dapat mengubah mikrobioma usus untuk mendukung bakteri menguntungkan dan menghambat patogen pada krustasea. Pengurangan Vibrio spp. yang signifikan dan peningkatan Xanthomarina pada udang yang disuplemen inulin menjadi perhatian khusus, karena ini menemukan penguatan dalam karya Wang et al. ( 2020 ), yang membahas peran prebiotik dalam menggeser keseimbangan mikroba usus menuju struktur komunitas yang meningkatkan kesehatan. Kehadiran genus bakteri tertentu seperti Deinococcus dan Devosia , yang terkait dengan kesehatan dan ketahanan terhadap penyakit pada udang, menekankan peran inulin dalam regulasi komunitas mikroba.
Analisis metabolomik studi kami menunjukkan perbedaan signifikan dalam jalur metabolisme utama, terutama yang terkait dengan metabolisme lipid, seperti biosintesis asam empedu, metabolisme asam arakidonat, dan mekanisme transportasi terkait. Hal ini konsisten dengan temuan oleh Defaix et al. ( 2024 ), yang mengamati bahwa serat makanan seperti inulin memberikan efek mendalam pada metabolisme lipid, meningkatkan biosintesis lipid yang bermanfaat sekaligus mengurangi yang berbahaya. Pengelompokan berbeda yang diamati dalam plot PCA dan OPLS-DA selanjutnya mengonfirmasi efek inulin yang nyata pada profil metabolisme, konsisten dengan temuan dari studi lain, seperti yang dilakukan oleh Singh et al. ( 2021 ), yang melaporkan peran multifaset inulin dalam mengubah spektrum metabolit dalam budidaya laut. Khususnya, peningkatan metabolit seperti Desglucocheirotoxin menunjukkan bahwa inulin tidak hanya memengaruhi jalur metabolisme yang umum tetapi juga dapat meluas hingga memengaruhi keberadaan senyawa bioaktif yang dapat memiliki implikasi farmasi.
Analisis integrasi menjelaskan interaksi kompleks antara metabolom dan mikrobiota, terbukti dalam penyelarasan antara metabolit tertentu dan bakteri tertentu. Mayoritas korelasi yang diamati kuat dan langsung, mirip dengan pengamatan dalam penelitian oleh Martinez-Porchas et al. ( 2023 ), yang menyoroti keterkaitan aktivitas mikroba dan profil metabolit dalam lingkungan akuatik. Korelasi negatif yang dicatat, seperti yang terjadi pada Koagulin R 3-glukosa, dapat berperan penting untuk memahami dinamika kompetitif atau penghambatan dalam mikroflora, suatu aspek yang dieksplorasi lebih lanjut oleh Freilich et al. ( 2011 ) dalam analisis mereka tentang persaingan metabolik dalam komunitas mikroba. Lebih jauh, interaksi seperti antara Halocynthiibacter dan Daumone menyoroti adaptasi perilaku mikroba dalam menanggapi tekanan lingkungan atau pergeseran komunitas. Daumone, yang diketahui berperan dalam jalur pensinyalan, dapat diproduksi oleh Halocynthiibacter sebagai sarana komunikasi atau sebagai mekanisme pengaturan untuk memodulasi lingkungan mikroba di sekitarnya. Hal ini menunjukkan bahwa Halocynthiibacter mungkin memiliki jalur metabolisme spesifik untuk mendegradasi, memanfaatkan, atau mensintesis Daumone, yang berpotensi memengaruhi interaksi komunitas yang lebih luas. Hubungan ini menggambarkan tekanan ekologis dan evolusi yang membentuk struktur dan fungsi komunitas mikroba, yang menunjukkan sifat canggih ekologi mikroba.
5 KESIMPULAN
Studi ini, yang dipadukan dengan temuan-temuan komparatif dari literatur terkini, menggarisbawahi potensi signifikan inulin sebagai prebiotik yang secara nyata memengaruhi pertumbuhan, kesehatan, metabolisme, dan keragaman mikroba pada udang. Efek-efek ini dimediasi melalui dampak langsung pada jalur-jalur metabolisme dan efek-efek tidak langsung melalui perubahan-perubahan dalam lanskap mikroba usus dan modulasi aktivitas-aktivitas enzim. Penelitian lebih lanjut harus difokuskan pada perluasan cakupan studi-studi ini ke berbagai spesies akuatik dan mengeksplorasi efek-efek jangka panjang dari suplementasi inulin dalam berbagai kondisi lingkungan.