
Abstrak
- Dalam perubahan iklim, pergeseran rezim gangguan mengharuskan pemahaman tentang hubungan antara regenerasi biomassa dan fungsi tanaman untuk menilai ketahanan hutan. Di hutan tropis, pemulihan biomassa didorong oleh dua mekanisme: perekrutan tanaman dan perkecambahan batang. Meskipun perekrutan tanaman telah dipelajari dengan baik, dasar fisiologis dan fungsional perkecambahan masih belum dieksplorasi. Studi ini mengatasi kesenjangan ini dengan memperkenalkan laju pertumbuhan perkecambahan relatif sebagai metrik baru dan mengevaluasi hubungannya dengan strategi fisiologis, menawarkan wawasan tentang bagaimana sifat perkecambahan selaras dengan trade-off fungsional untuk meningkatkan model pemulihan biomassa global dalam berbagai rezim gangguan.
- Dengan menggunakan hutan kering tropis Karibia, di mana perkecambahan mendominasi regenerasi pascagangguan, kami menyelidiki peran kontinum cepat-berisiko versus lambat-aman dalam ekonomi sumber daya pohon, pertumbuhan, kelangsungan hidup, dan kinerja perkecambahan. Kami menganalisis 10 produktivitas dan sifat hidrolik di 44 spesies hutan kering tropis Puerto Rico. Hubungan antara strategi sumber daya, pertumbuhan dewasa, kelangsungan hidup, dan perkecambahan dianalisis lintas spesies dan dengan kontrol filogenetik untuk menilai signifikansi evolusi.
- Kontinum cepat-berisiko vs. lambat-aman muncul sebagai trade-off utama dalam ekosistem ini. Ciri-ciri produktivitas dan ketahanan kekeringan memprediksi pertumbuhan pohon dewasa tetapi tidak kelangsungan hidup. Khususnya, pertumbuhan berkorelasi positif dengan kerentanan terhadap kegagalan hidrolik, menekankan perlunya mempertimbangkan kembali margin keamanan hidrolik (HSM) di hutan tempat resprouting mengurangi kehilangan batang akibat kekeringan, yang mempersulit penilaian tradisional tentang mortalitas dan pemulihan biomassa. Kinerja resprouting berkorelasi dengan trade-off cepat-berisiko/lambat-aman, dengan spesies yang tumbuh cepat dan menghindari kekeringan menunjukkan resprouting yang lebih kuat, menyoroti strategi penggunaan air sebagai hal yang penting untuk regenerasi pasca-gangguan. Selain itu, hubungan positif antara fosfor daun, pertumbuhan dewasa dan pertumbuhan resprouting dipengaruhi oleh nenek moyang yang sama, yang menunjukkan bahwa ciri-ciri seperti asosiasi ektomikoriza dan fiksasi nitrogen, yang lazim di hutan tropis yang miskin nutrisi, dapat mendukung hubungan ini.
- Berdasarkan temuan kami, kami menghubungkan keberhasilan tunas baru dengan strategi fisiologis pohon di mana pemisahan produktivitas dan ciri hidrolik di antara spesies kemungkinan menentukan respons berbeda terhadap angin dan kekeringan yang membentuk rezim gangguan alami di hutan kering tropis Karibia.
Baca Ringkasan Bahasa Sederhana gratis untuk artikel ini di blog Jurnal.
1. PENDAHULUAN
Perubahan iklim memengaruhi rezim gangguan di daerah tropis (Johnstone et al., 2016 ) yang meningkatkan kebutuhan untuk meningkatkan prediksi tentang bagaimana tanaman merespons peristiwa gangguan. Resprouting adalah sifat terkait gangguan di antara banyak tanaman tropis yang menggambarkan pembentukan tunas baru untuk menggantikan hilangnya biomassa hidup. Secara keseluruhan, literatur resprouting sangat berfokus pada perbedaan biner (yaitu resprouter vs. nonresprouter) dalam ekosistem yang didominasi api, sebuah fenomena yang tercermin dengan baik dalam model vegetasi (Hanbury-Brown et al., 2022 ). Namun, perspektif terbatas ini menghambat kemampuan kita untuk memprediksi regenerasi pasca-gangguan di banyak ekosistem tropis non-pirik yang tersebar luas di mana resprouting juga terjadi dan yang mewakili tempat perlindungan penting bagi keanekaragaman hayati (Armenteras et al., 2021 ; Vesk & Westoby, 2004 ). Lebih jauh lagi, dalam lingkungan yang berbeda, resprouting kemungkinan dikaitkan dengan serangkaian sifat yang berbeda (Pausas et al., 2016 ). Misalnya, resprouter bervariasi antara sabana Afrotropis dan beberapa sabana Neotropis dalam sifat respons kebakarannya: dari mengoptimalkan regenerasi pasca-kebakaran melalui bank pohon muda yang persisten dan perlindungan melalui kulit kayu yang sangat tebal yang juga melindungi tunas batang dari panas (Pausas, 2015 ). Dengan demikian, pertimbangan korelasi sifat yang lebih luas ini sangat penting untuk memajukan prediksi respons vegetasi pada skala yang lebih besar di mana variasi dalam kinerja resprouting (misalnya inisiasi tunas, pertumbuhan, kelangsungan hidup) dapat dengan yakin dikaitkan dengan sindrom fungsional kritis dalam ekosistem.
Spektrum ekonomi cepat-lambat, kerangka kerja berbasis sifat, menawarkan lensa yang berharga untuk memprediksi respons tanaman terhadap gangguan di ekosistem tropis non-pirik (diulas oleh Oliveira et al., 2021 ). Kerangka kerja ini mengkarakterisasikan kontinum strategi ekonomi sumber daya di antara spesies, mulai dari ‘strategi produktif’ yang difokuskan pada perolehan sumber daya yang cepat (misalnya luas daun spesifik [SLA] yang lebih tinggi dan kandungan nutrisi daun, yang meningkatkan fotosintesis dan pertumbuhan) hingga ‘strategi konservatif’ (misalnya SLA yang lebih rendah dan kandungan nutrisi daun) yang menekankan konservasi sumber daya dan pengembalian yang lebih lambat. Strategi-strategi ini mengatur trade-off dalam pertumbuhan dan kelangsungan hidup dewasa di berbagai bioma tropis (Poorter, Kitajima, et al., 2010 ; Poorter, McDonald, et al., 2010 ). Bagaimana trade-off pertumbuhan-kelangsungan hidup ini berhubungan dengan kinerja resprouting masih belum jelas. Resprouting—strategi persistensi yang bergantung pada cadangan yang tersimpan—secara konvensional telah dikaitkan dengan sifat-sifat lambat, dengan asumsi bahwa mengalokasikan sumber daya ke cadangan membatasi pertumbuhan yang cepat (Bond & Midgley, 2001 ). Namun, penelitian sebelumnya telah mengaitkan kinerja resprouting yang kuat dengan strategi cepat dan lambat (misalnya Paz et al., 2018 ; Poorter, Kitajima, et al., 2010 ; Poorter, McDonald, et al., 2010 ), dengan beberapa penelitian menunjukkan bahwa hal itu mewakili sumbu sifat yang independen (Heineman et al., 2021 ). Khususnya, kekurangan yang jelas dari pekerjaan sebelumnya adalah karakterisasi strategi cepat-lambat hanya dengan satu atau dua sifat (yang paling umum adalah luas daun dan kepadatan kayu tertentu).
Karena kekeringan semakin diterima sebagai penentu utama kematian pohon di daerah tropis (Allen et al., 2015 ), ada kebutuhan khusus untuk mengungkap hubungan antara sifat hidrolik yang membentuk strategi ketahanan kekeringan dan pertunasan kembali. Sifat seperti kepadatan kayu dan efisiensi hidrolik telah terbukti menjadi kunci dalam hal ini, dengan kayu yang lebih padat dan efisiensi hidrolik yang berkurang terkait dengan ketahanan kekeringan dan pertumbuhan konservatif, sementara kepadatan kayu yang lebih rendah dan efisiensi hidrolik yang lebih tinggi mendukung penggunaan sumber daya yang cepat tetapi meningkatkan kerentanan kekeringan. Sifat stomata dan hidrolik, seperti konduktansi stomata maksimum, margin keamanan hidrolik, dan potensi air daun maksimum, juga mencerminkan trade-off ini, menyeimbangkan penggunaan air dan perolehan karbon dengan risiko kekeringan. Penyelarasan antara ketahanan kekeringan dan trade-off pertumbuhan-kematian telah dicirikan sebagai spektrum cepat-berisiko/lambat-aman, yang menghubungkan penutupan stomata yang disebabkan oleh kekeringan dengan keterbatasan perolehan CO 2 (Brodribb & McAdam, 2017 ). Atas dasar itu, kerangka kerja cepat-berisiko/lambat-aman menawarkan jalan yang menjanjikan untuk menguji ketahanan hutan tropis terhadap gangguan, terutama dalam sistem di mana pertumbuhan kembali memainkan peran penting.
Pola alokasi karbon, yang dibentuk oleh tekanan selektif dan rezim gangguan, sangat memengaruhi demografi dan kinerja resprouting. Dalam sistem yang rawan kebakaran, tanaman yang resprouting dapat mengalokasikan sumber daya di bawah tanah sebagai respons terhadap kejadian dengan tingkat keparahan tinggi, yang membatasi pertumbuhan dan reproduksi di atas tanah dibandingkan dengan nonsprouters (Pausas et al., 2016 ). Di hutan tropis bebas kebakaran, di mana kejadian destruktif seperti itu lebih jarang terjadi, resprouting lebih bergantung pada fungsi batang, seperti penyimpanan karbohidrat non-struktural, menampung jaringan pembentuk kuncup dan subsidi fotosintesis untuk pertumbuhan resprout (Herrera-Ramírez et al., 2021 ; Pfanz et al., 2002 ; Van Bloem et al., 2003 ). Strategi yang kontras ini menunjukkan bahwa alokasi karbon dalam sistem tropis non-pirik mungkin berbeda secara substansial dari sistem yang rawan kebakaran, dengan implikasi untuk trade-off pertumbuhan-kematian. Selain itu, ekosistem Mediterania menunjukkan bahwa spesies yang tumbuh kembali dengan rasio akar:pucuk yang lebih tinggi—yang dikaitkan dengan pertumbuhan yang lebih lambat—sering kali menunjukkan sifat-sifat seperti efisiensi hidrolik yang lebih rendah dan berkurangnya resistensi kavitasi xilem, yang mencerminkan strategi toleransi kekeringan (Hernández et al., 2011 ; Pratt et al., 2007 ; Vilagrosa et al., 2014 ). Pola Mediterania tampak kontras dengan pengamatan dari hutan tropis (baik hutan lembap maupun kering) di mana penyimpanan umumnya dikaitkan dengan strategi penghindaran kekeringan untuk memungkinkan spesies gugur daun akibat kekeringan yang tumbuh lebih cepat untuk bertahan hidup dalam periode saat tidak berdaun (Poorter, Kitajima, et al., 2010 ; Poorter, McDonald, et al., 2010 ). Namun, peran penyimpanan menjadi rumit karena pemahaman kita yang buruk tentang karbohidrat non-struktural di akar di hutan tropis relatif terhadap sistem Mediterania. Secara keseluruhan, rezim gangguan yang dialami di hutan tropis diperkirakan akan menentukan pola alokasi C dengan cara yang memengaruhi demografi dan ketahanan kekeringan spesies yang tumbuh kembali.
Kami berusaha untuk mendapatkan wawasan tentang ketahanan hutan kering tropis (TDF) dengan menilai bagaimana kinerja spesies yang tumbuh kembali berhubungan dengan spektrum ekonomi cepat-berisiko/lambat-aman. Kami menggunakan data dari komunitas TDF yang beragam secara filogenetik di Karibia (Lampiran S1 ) karena dominasi pohon yang tumbuh kembali sebagai mekanisme regenerasi dalam menanggapi angin dan kekeringan (Murphy & Lugo, 1986 ; Van Bloem et al., 2003 ). Di seluruh makalah ini kami menggunakan istilah ‘kinerja tumbuh kembali’ untuk menggambarkan dua proses yang mencirikan potensi regenerasi spesies melalui pengembangan tunas baru di pangkal pohon (yaitu basal atau akar). Istilah ‘inisiasi kuncup’ menggambarkan inisiasi kuncup segera setelah gangguan sementara ‘pertumbuhan tunas kembali’ menggambarkan kemampuan untuk meningkatkan panjang tunas dalam jangka waktu tertentu. Agen yang berbeda bertanggung jawab untuk memengaruhi keberhasilan setiap proses. Misalnya, gangguan dapat memengaruhi inisiasi tunas dengan merusak jaringan meristematik atau mengurangi sumber daya yang diperlukan sementara kondisi abiotik akan memengaruhi pertumbuhan tunas kembali dengan menentukan ukuran kumpulan sumber daya dan pola alokasi sumber daya (Pausas et al., 2016 ). Dalam pengalaman kami, tunas kembali biasanya terjadi setelah efek struktural utama pada batang (misalnya pembakaran, pemotongan atau efek angin) tetapi beberapa tunas kembali terjadi pada tingkat latar belakang (Van Bloem et al., 2003 ) mungkin lebih terkait dengan kekeringan. Semua spesies dapat bertunas dari batang, tetapi tidak dari akar (pengamatan pribadi) dan tunas dominan (yang terpanjang yang dihasilkan dari suatu kelompok) cenderung berada di atau dekat permukaan tanah (yaitu pengisap akar atau di bawah 10 cm di batang) (Van Bloem et al., 2007 ). Hanya 7 tahun setelah badai besar, lebih dari 13% tunas yang berkembang sudah setinggi subkanopi, sementara mortalitas pohon yang bertunas sekitar 2% lebih rendah daripada pohon yang tidak bertunas. Dengan demikian, tingkat pertumbuhan dan tingkat kematian yang lebih rendah pada batang yang bertunas kembali menunjukkan bahwa bertunas kembali bukan sekadar cara yang lebih lambat bagi pohon untuk mati (Van Bloem et al., 2007 ).
Untuk penelitian yang disajikan di sini, kami bertanya: (a) Apakah spesies TDF menunjukkan trade-off dalam strategi penggunaan sumber daya yang menunjukkan kontinum cepat-berisiko/lambat-aman? Kami memperkirakan trade-off cepat-berisiko/lambat-aman membentuk sumbu utama variasi sifat di antara spesies pohon dalam studi TDF kami. (b) Apakah pertumbuhan dan kelangsungan hidup spesies dewasa berkorelasi dengan sifat ekonomi cepat-berisiko/lambat-aman? Kami berhipotesis pertumbuhan yang lebih cepat dan kelangsungan hidup yang lebih rendah di antara pohon-pohon yang memperoleh sumber daya dengan ketahanan kekeringan yang lebih rendah dan strategi hidrolik yang ‘lebih berisiko’ (yaitu potensi air daun maksimum yang lebih rendah, margin keamanan hidrolik yang lebih rendah, kepadatan kayu yang lebih rendah dan efisiensi hidrolik yang lebih tinggi) sedangkan spesies konservatif dengan toleransi kekeringan yang lebih besar (yaitu potensi air daun maksimum yang lebih tinggi, margin keamanan hidrolik yang lebih tinggi, kepadatan kayu yang lebih tinggi dan efisiensi hidrolik yang lebih rendah) akan memiliki kelangsungan hidup yang lebih besar tetapi tumbuh lebih lambat. (c) Untuk spesies yang dapat bertunas kembali, bagaimana strategi cepat-berisiko/lambat-aman mereka, bersama dengan sifat pertumbuhan dan kelangsungan hidup mereka, dikaitkan dengan kinerja bertunas kembali mereka? Kami berhipotesis bahwa spesies yang tumbuh lebih cepat dan terhindar dari kekeringan (yaitu potensi air daun maksimum yang lebih rendah, konduktansi stomata musim kemarau yang lebih rendah, margin keamanan hidrolik yang lebih rendah, dan efisiensi hidrolik yang lebih tinggi) akan menunjukkan kinerja resprouting yang lebih unggul (misalnya pertumbuhan resprout yang lebih cepat atau inisiasi tunas yang lebih besar). Spesies ini memiliki kecenderungan lebih besar untuk menjadi gugur daun akibat kekeringan dan karena itu memiliki cadangan yang lebih besar untuk bertahan hidup pada periode tanpa daun dan mendorong resprouting. Sebaliknya, spesies yang tahan kekeringan (yaitu potensi air daun maksimum yang lebih tinggi, konduktansi stomata musim kemarau yang lebih tinggi, margin keamanan hidrolik yang lebih tinggi, dan efisiensi hidrolik yang lebih rendah) mungkin berinvestasi lebih sedikit dalam penyimpanan, karena mereka sering mempertahankan sebagian besar kanopi mereka selama musim kemarau. Prediksi ini mengasumsikan penyimpanan akar minimal, faktor yang kurang dipahami di hutan tropis.
2 BAHAN DAN METODE
2.1 Lokasi penelitian
Kami melakukan penelitian kami di Hutan Negara Guánica seluas 4500 ha, TDF semi-gugur di barat daya Puerto Riko (17°58′ LU, 65°30′ BB). Suhu tahunan rata-rata adalah 25,1°C dan curah hujan tahunan rata-rata 910 mm ±179 SD (basis data CHIRPS diperoleh dari earthengine.google.com ), meskipun variasi curah hujan yang cukup besar terjadi dalam dan antar tahun (Murphy & Lugo, 1986 ). Musim bersifat bimodal, terbagi antara musim dingin (Desember–Maret) dan pertengahan musim panas (Juni–Agustus) musim kemarau. Tanah sebagian besar adalah mollisol pada batu kapur yang relatif basa (pH 7,8). Bioavailabilitas P yang rendah dianggap sebagai faktor pembatas pada produktivitas tanaman (Murphy & Lugo, 1986 ). Banyak spesies pohon memiliki kecenderungan kuat untuk bertunas kembali, terutama setelah badai yang kuat (Van Bloem et al., 2003 ). Struktur hutan dicirikan oleh kerapatan batang yang relatif tinggi (rata-rata 14.000 batang/ha dengan diameter batang minimal 2,5 cm; Murphy & Lugo, 1986 ), dengan sekitar 42% pohon berbatang ganda—refleksi jelas dari mekanisme resprouting yang kuat di antara spesies (Van Bloem et al., 2003 ). Kebakaran bukan merupakan hal endemik di Hutan Guánica (Murphy & Lugo, 1986 ; Wolfe et al., 2014 ).
2.2 Spesies dan sifat-sifatnya
Izin kerja lapangan diberikan oleh E Martínez, Pejabat Pengelolaan Hutan Guánica. Kami memilih sifat berdasarkan karakteristik tanaman yang paling efektif mewakili spektrum antara strategi ‘cepat-berisiko’ dan ‘lambat-aman’ (Oliveira et al., 2021 ). Kumpulan data kami yang terdiri dari 10 sifat menggabungkan data yang diterbitkan sebelumnya (Vargas et al., 2021 ) dengan pengukuran yang baru dikumpulkan (Tabel 1 ). Jumlah spesies dengan data sifat yang tersedia bervariasi antara 19 dan 44, selain dari margin keamanan hidrolik (HSM), yang datanya hanya tersedia untuk enam spesies (Lampiran S2 memberikan rincian ketersediaan spesies berdasarkan sifat). Ke-44 spesies tersebut mencakup >90% dari semua individu ≥2,5 cm dbh di hutan (Murphy & Lugo, 1986 ). Data sifat baru dikumpulkan antara tahun 2017 dan 2018, bertepatan dengan musim hujan (Agustus–November) kecuali sifat yang menggambarkan perilaku stomata ( ψ m , Δ ψ l , dan g smax ), yang dikumpulkan selama musim kemarau sebelum daun gugur. Luas daun spesifik dan nutrisi daun dikumpulkan dari daun dewasa yang tidak rusak dari kanopi luar yang terkena sinar matahari dari 5–12 individu. Daun direhidrasi semalaman sebelum pengukuran massa segar menggunakan neraca presisi (± 0,001 g). Luas permukaan daun termasuk tangkai daun diperoleh menggunakan pemindai alas datar dan ImageJ (Schneider et al., 2012 ; https://imagej.net/ ). Selanjutnya, sampel dikeringkan dalam oven pada suhu 75°C selama 72 jam sebelum massa kering diukur. Kandungan nitrogen daun (LNC) dan kandungan fosfor daun (LPC) diukur dari sampel daun massal menggunakan pencernaan hidrogen peroksida asam nitrat sebelum analisis ICP-OES (ICP-6500 Duo, Thermo Fisher Scientific, Pittsburg, PA, AS). Kepadatan kayu (WD) ditentukan menggunakan inti yang diekstraksi pada dbh dari lima individu per spesies. Individu pohon berkisar antara 2,5 hingga 15 cm dbh. Setelah kulitnya dikupas, inti (berkisar antara panjang 6,8 hingga 43,8 mm, diambil dari ujung empulur) direndam dalam ruang vakum selama 1 minggu untuk memastikan hidrasi penuh. Kami mengukur volume inti menggunakan metode perpindahan air (Ilic et al., 2000 ) setelah itu sampel dikeringkan dalam oven selama 96 jam pada 100 °C dan ditimbang.
| Sifat | Singkatan. | Satuan | N spp. | Definisi/rumus | Sumber data |
|---|---|---|---|---|---|
| Ciri-ciri ekonomi sumber daya | |||||
| Luas daun spesifik | Persyaratan Layanan | cm2 gram -1 | 44 | Luas permukaan/massa kering daun | Data yang baru dikumpulkan, musim hujan 2017–18 |
| Kandungan Nitrogen Daun | LNC | % | 36 | Persentase Nitrogen dalam Massa Kering Daun | Data yang baru dikumpulkan, musim hujan 2017–18 |
| Kandungan fosfor daun | LPC | mgg -1 | 43 | Kandungan fosfor berdasarkan massa dalam massa kering daun | Data yang baru dikumpulkan, musim hujan 2017–18 |
| Kepadatan kayu | WD | gram cm3 | 43 | Massa kayu kering/volume segar. | Data yang baru dikumpulkan, musim hujan 2017–18 |
| Potensi air daun minimum musim kemarau | ψ m | -MPa | 31 | Potensi air daun minimum tercatat antara pukul 12:00 dan 14:00 | Data yang baru dikumpulkan, musim kemarau 2017–18 |
| Kisaran diurnal potensi air daun pada musim kemarau | Δ ψ L | -MPa | 31 | ψ m — potensi air daun sebelum fajar yang tercatat antara pukul 04:00 dan 06:00 | Data yang baru dikumpulkan, musim kemarau 2017–18 |
| Konduktivitas hidrolik spesifik xilem asli | K S | kg m 2 s -1 MPa -1 | 19 | Efisiensi transportasi air melalui xilem batang pada kapasitas lapangan | Data yang diarsipkan sebelumnya (Vargas et al., 2021 ) |
| Nilai Huber | TTV | tanpa kesatuan | 19 | Luas penampang cabang gubal/total luas daun cabang | Data yang diarsipkan sebelumnya (Vargas et al., 2021 ) |
| Konduktansi stomata maksimum pada musim kemarau | g kecil | mmol m -2 s -1 | 31 | Konduktansi stomata daun maksimum pada bulan Januari 2017 | Data yang baru dikumpulkan, musim kemarau 2017–18 |
| Margin keamanan hidrolik | HSM | -MPa | 6 | ψ m — tekanan xilem yang menyebabkan hilangnya konduktansi hidrolik sebesar 50% ( P 50 ) | ψ m —data yang baru dikumpulkan, musim kemarau 2017–18; P 50 —data yang diarsipkan sebelumnya (Vargas et al., 2021 ) |
| Pertumbuhan dan kelangsungan hidup pohon | |||||
| Laju pertumbuhan diameter pohon dewasa | GR | cm tahun -1 | 32 | Kemiringan regresi dbh terhadap tanggal pengukuran yang sesuai | Data yang baru dikumpulkan, 1992–2019 |
| Tingkat kelangsungan hidup pohon dewasa | Bahasa Indonesia: SR | % tahun −1 | 32 | Angka kematian tahunan (MR) = log e N 0 − log e N 1 /waktu (tahun)
Tingkat kelangsungan hidup tahunan (SR) = 100 × e −MRx1 |
Data yang baru dikumpulkan, 1998–2019 |
| Tumbuh kembali | |||||
| Inisiasi tunas kembali | Nomor RS | tanpa kesatuan | 35 | Jumlah tunas baru yang dihasilkan dalam 119 hari pertama | Kebakaran—data yang baru dikumpulkan, musim kemarau 2012; Badai—data yang sebelumnya diarsipkan (Van Bloem et al., 2003 ); Penebangan habis—data yang sebelumnya diarsipkan (Dunewitz, 1985 ) |
| Laju pertumbuhan relatif panjang tunas kembali | RSUD | cm × cm −1 hari −1 | 35 | (ln H 2 − ln H 1 )/( t 2 − t 1 ) dimana H 1 dan t 1 adalah tinggi dan waktu pada saat munculnya tunas baru dan H 2 dan t 2 adalah tinggi dan waktu munculnya tunas baru pada umur 119 hari. | |
Catatan : Tabel ini mencakup jumlah spesies yang dianalisis untuk setiap sifat ( N spp.), definisi sifat dan sumber data. SLA yang lebih tinggi, kandungan nutrisi daun dan K tetapi HV yang lebih rendah merupakan indikasi strategi yang mendukung penggunaan sumber daya yang cepat tetapi meningkatkan kerentanan kekeringan (yaitu spesies yang ‘cepat-berisiko’). WD yang lebih tinggi, g smax musim kemarau , Δ ψ L dan HSM tetapi lebih negatif ψ m , terkait dengan ketahanan terhadap kekeringan dan pertumbuhan konservatif (yaitu spesies yang ‘lambat-aman’).
Toleransi kekeringan ditentukan menggunakan potensi air daun minimum ( ψ m ), kisaran diurnal potensi air daun (Δ ψ l ) dan konduktansi stomata musim kemarau maksimum ( g smax ; Bhaskar & Ackerly, 2006 ; Hoshika et al., 2018 ). Nilai yang lebih negatif dari ψ m dan Δ ψ l musim kemarau dan nilai g smax yang lebih besar menunjukkan penggunaan air yang lebih liberal dalam menanggapi kondisi pengeringan. ψ m berkorelasi baik dengan ukuran toleransi kekeringan yang lebih langsung termasuk resistensi emboli ( P 50 ; berpasangan r = 0,95, p < 0,001) dan titik kehilangan turgor daun ( ψ TLP ; berpasangan r = 0,63, p < 0,001) di antara subset spesies pohon kami yang lebih kecil (Allerton, tidak dipublikasikan ). Selama 15 hari berturut-turut pada bulan Maret 2017, kami mengukur potensi air daun ( ψ l ) dan konduktansi stomata daun ( g s ) pada interval dua jam antara pukul 04:00 dan 16:00 pada lima individu per spesies. Semua individu ditemukan di sepanjang jalan setapak hutan, dengan hanya daun yang sehat dan berkembang penuh yang dipilih untuk pengukuran. ψ l individu adalah tekanan yang diperlukan yang diterapkan pada satu daun yang terpapar sinar matahari untuk mendorong air keluar dari pembuluh xilem di ujung tangkai daun yang dipotong (Ruang tekanan Model 1000, Instrumen PMS). g s untuk interval tertentu adalah rata-rata dari tiga daun yang terpapar sinar matahari menggunakan porometer kondisi stabil SC-1 (Decagon). Setelah menghitung rata-rata di lima individu, hasilnya adalah nilai tunggal untuk ψ l dan g s per interval per spesies. ψ m adalah ψ l pada tengah hari (12:00–14:00; dalam semua kasus ψ l minimum ). Δ ψ l dihitung sebagai ψ m − predawn ψ l (04:00–06:00) sementara g smax adalah g s maksimum yang dicapai pada setiap interval setiap hari. Nilai ψ m dan Δ ψ l dibuat positif dengan mengalikan dengan −1.
Informasi tentang efisiensi hidrolik di lapangan dan kerentanan terhadap mortalitas akibat kekeringan diperoleh dari data yang diarsipkan (Vargas et al., 2021 ) termasuk konduktivitas hidrolik spesifik xilem ( K S ), nilai huber (HV) dan margin keamanan hidrolik (HSM). Lihat Vargas et al. ( 2021 ) untuk protokol pengumpulan lengkap mengenai K S , HV dan P50 . Konduktivitas hidrolik spesifik xilem ( K S ) diukur di laboratorium lapangan untuk lima individu per spesies. Setelah memperkirakan panjang pembuluh, batang baru dikumpulkan dari cabang tajuk berdiameter 1 hingga 2 cm sebelum fajar. Segmen cabang kemudian digunakan untuk mengukur laju aliran melalui xilem dengan mencatat perubahan meniskus melalui pipet presisi tinggi yang dikalibrasi. Secara keseluruhan, 3–4 pengukuran aliran tekanan dilakukan pada ketinggian yang bervariasi untuk mendapatkan nilai head tekanan keseluruhan. Kemiringan linier semua titik data (yaitu konduktivitas hidrolik) kemudian dibagi dengan luas xilem dan panjang segmen cabang. Nilai Huber (HV) adalah rasio penampang melintang gubal terhadap total luas daun distal batang. Cabang terminal yang terbuka dengan diameter sekitar 0,5 cm digunakan dari lima individu per spesies. Luas xilem diukur menggunakan jangka sorong digital di pangkal cabang sementara total luas daun dihitung dari analisis gambar di ImageJ (Schneider et al., 2012 ). HSM daun adalah perbedaan antara ψ m dan potensial air daun di mana xilem daun mengalami 50% kejadian emboli ( P50 ). Nilai HSM yang lebih sempit menunjukkan risiko kegagalan hidrolik yang lebih tinggi di bawah kekeringan ekstrem. Kurva kerentanan dibuat untuk setiap spesies dengan secara bersamaan menggunakan gambar digital resolusi tinggi untuk melacak dehidrasi daun sambil mengukur ψ l selama interval yang ditetapkan sepanjang hari. Fungsi sigmoidal kemudian dipasang untuk memperkirakan P50 . Nilai ψ m yang kami kumpulkan di lapangan kemudian digunakan untuk menghitung HSM untuk setiap spesies.
2.3 Pertumbuhan dan tingkat kelangsungan hidup pohon
Tingkat pertumbuhan (GR) tingkat populasi dan tingkat kelangsungan hidup tahunan (SR) yang spesifik spesies dihitung untuk individu dewasa dari empat plot sensus persegi permanen di Hutan Guánica. Plot-plot ini berkisar dalam ukuran dari 625 hingga 10.000 m 2 dan diberi jarak hingga ~3 km. Semua plot terletak di hutan kering tropis semi-gugur dataran tinggi (TDF) yang matang dengan tanah mollisol pada substrat batu kapur. Di dalam plot-plot ini, masing-masing pohon ditandai, diidentifikasi berdasarkan spesies, dan dipantau untuk mortalitas dan diameter setinggi dada (dbh) selama setiap sensus. Untuk mengurangi efek yang bergantung pada ukuran, tingkat pertumbuhan diameter dihitung untuk individu dalam kisaran dbh terbatas 2,5–15 cm. Selama sensus awal tahun 1992, batang dalam kisaran ukuran ini dicatat. Untuk meningkatkan ukuran sampel, pohon-pohon baru ditambahkan selama sensus berikutnya yang dilakukan setiap 1–8 tahun. Sensus pasca-1992 mengukur batang dalam kisaran dbh 2,5–10 cm untuk lebih mengurangi potensi efek ontogenetik pada pertumbuhan. Distribusi kelas ukuran awal dirinci dalam Murphy dan Lugo ( 1986 ). Laju pertumbuhan dihitung untuk 32 spesies, sementara tingkat kelangsungan hidup ditentukan untuk 30 spesies, dengan ukuran sampel median tujuh individu per spesies (kisaran: 2–138). Untuk menstandardisasi rentang waktu pengukuran di antara plot, kami menggunakan data yang dikumpulkan antara tahun 1992 dan 2019, menghasilkan periode pemantauan 20,4–26,4 tahun untuk pengukuran dbh di seluruh spesies. Mengingat laju pertumbuhan lambat yang khas dari spesies TDF (Murphy & Lugo, 1990 ), prosedur diterapkan untuk memastikan akurasi pengukuran, termasuk: (a) mengukur diameter pada interval 5 cm di lima lokasi batang yang konsisten yang membatasi tinggi dada (1,3–1,5 m) selama setiap sensus menggunakan pita diameter; (b) mengecualikan pengukuran musim hujan untuk menghindari artefak pembengkakan; dan (c) mengabaikan pengukuran dari dua sensus sebelumnya untuk individu yang kemudian mati, karena sulitnya membedakan antara mortalitas dan dormansi. Laju pertumbuhan diperoleh dari kemiringan regresi dbh terhadap waktu, dengan menggabungkan minimal dua pengukuran dbh per individu (mengikuti Poorter, Kitajima, et al., 2010 ; Poorter, McDonald, et al., 2010 ). Tingkat kelangsungan hidup tahunan (SR) dihitung selama rentang waktu 21 tahun (1998–2019) untuk menghindari bias yang disebabkan oleh kekeringan hebat pada tahun 1997. Persamaan berikut digunakan:
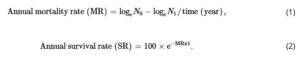
2.4 Kinerja perkecambahan kembali
Data tentang pertumbuhan kembali pohon setelah gangguan dihimpun dari sumber asli dan yang diarsipkan (sebelumnya tidak dipublikasikan), yang mencakup tiga jenis gangguan berbeda yang terjadi di Hutan Guánica: penebangan habis, kebakaran, dan badai. Semua plot gangguan berisi hutan kering tropis dewasa semi-gugur dataran tinggi di atas tanah mollisol dan substrat batu kapur. Jarak plot di seluruh plot gangguan membentang maksimal ~13 km. Data kebakaran berasal dari dua lokasi yang terbakar pada bulan Februari dan Mei 2012, terpisah sejauh ~13 km. Pertumbuhan kembali pohon pertama kali diukur pada bulan Juni 2012, dengan pengukuran kedua dari panjang pertumbuhan kembali terpanjang per individu dilakukan rata-rata 49,5 hari kemudian. Data badai dikumpulkan dari empat plot yang terkena dampak Badai Georges pada bulan September 1998, yang terletak dalam radius 3 km. Sensus pertumbuhan kembali pohon pertama untuk pohon yang rusak akibat badai terjadi 15 minggu pasca-badai (rata-rata 119 hari), dengan pengukuran kedua dilakukan sekitar 630 hari kemudian. Data penebangan habis diperoleh dari lima plot (<100 m terpisah) di mana semua pohon dalam setiap plot 10 × 10 m ditebang di pangkalnya pada bulan Juni 1981. Pertumbuhan kembali pertama kali diukur pada bulan November 1981 (rata-rata 130 hari pasca penebangan habis) dan sensus kedua dilakukan pada bulan Maret 1982, sekitar 135 hari kemudian. Pengujian tambahan menunjukkan kinerja pertumbuhan kembali dapat dipengaruhi oleh tingkat kehilangan biomassa di atas tanah (Lampiran S3 ). Untuk memastikan konsistensi, kami menstandardisasi data dengan memasukkan tunas baru dari lokasi basal dan akar pada pohon yang mengalami kehilangan biomassa di atas tanah ≥60%. Pendekatan ini juga sejalan dengan studi dalam sistem yang rawan kebakaran, di mana pertumbuhan kembali umumnya didefinisikan sebagai pohon yang mampu menghasilkan tunas baru setelah kehilangan biomassa lengkap (100%), yang memungkinkan perbandingan yang lebih kuat.
Untuk semua gangguan, data resprouting mencakup jumlah tunas baru ( RS num ; diambil dari sensus pertama pasca-gangguan), dan panjang resprout terpanjang. Untuk setiap individu, tunas baru dihitung menurut protokol yang ada di hutan ini, yang berasal dari tanah dan yang ada di batang ≤30 cm dan ≤45° dari vertikal (Van Bloem et al., 2003 ). Pengukuran berulang pada sensus satu dan dua melacak panjang resprout terpanjang untuk menghitung laju pertumbuhan relatif (tinggi) ( RS RGR ) menggunakan Persamaan (3). Secara total, 1015 individu dari 36 spesies dinilai, dengan 704 menghasilkan resprout basal dan 311 dari akar. Penebangan habis memberikan kontribusi individu dan spesies terbanyak, tetapi kurva rarefaksi menunjukkan spesies yang paling umum ditangkap di antara tiga jenis gangguan (Lampiran S4 ),
![]()
di mana H 1 dan t 1 adalah panjang dan waktu pada inisiasi tunas baru (yaitu sensus pertama) dan H 2 dan t 2 adalah panjang dan waktu tunas baru pada sensus kedua. Panjang tunas baru ditransformasikan ke dalam ln sebelum dirata-ratakan untuk meniadakan bias yang terkait dengan peningkatan varians RS RGR dengan ukuran sampel yang lebih tinggi (Hoffmann & Poorter, 2002 ).
2.5 Analisis Statistik
Data diperiksa untuk memeriksa apakah data tersebut memenuhi asumsi analisis parametrik kami termasuk menggunakan distribusi dan uji Shapiro–Wilks. Untuk variabel yang menunjukkan kemiringan berat atau distribusi non-normal, transformasi log dilakukan. Kovariasi sifat di antara spesies dinilai menggunakan analisis komponen utama (PCA) dan nilai sifat rata-rata berskala. Kesenjangan data diperhitungkan menggunakan nilai dari spesies saudara terdekat (Lampiran S1 untuk pohon filogenetik, Lampiran S2 untuk kesenjangan data). Pertumbuhan dewasa, kelangsungan hidup, metrik resprouting dan kebiasaan daun disertakan a posteriori sebagai variabel ‘tambahan’ terhadap sumbu sifat turunan menggunakan paket R ‘FactoMineR’ (Lê et al., 2008 ). Kami mengecualikan HSM karena data hanya tersedia untuk enam spesies. Hubungan di antara sifat, pertumbuhan dan kelangsungan hidup dewasa, dan kinerja resprouting dinilai menggunakan korelasi berpasangan Pearson. Kami juga menggunakan kontras independen filogenetik (PIC; Felsenstein, 1985 ) untuk menguji hipotesis nol bahwa hubungan lintas spesies tidak ditentukan oleh nenek moyang yang sama. Alasan di sini adalah bahwa sifat-sifat akan cenderung serupa di antara spesies yang berkerabat dekat karena mereka menempati relung yang sama, diwarisi dari nenek moyang yang sama. Data yang diperhitungkan dikecualikan dari korelasi lintas spesies dan filogenetik. Menggunakan korelasi daripada regresi memungkinkan kami untuk membandingkan tanda dan kekuatan uji lintas spesies dan filogenetik. Menggunakan paket R ‘brranching’ (Chamberlain, 2016 ), data filogenetik diimpor dari Phylomatic berdasarkan filogeni angiospermae yang terselesaikan dengan baik (pohon R20120829) oleh Smith et al. ( 2011 ). PIC dihitung menggunakan paket ‘ape’ (Paradis & Schliep, 2019 ). Kami menyelesaikan satu politomi di pohon kami dengan menempatkan simpul baru pada jarak nol dari simpul politomi asli. Asumsi model PIC diperiksa termasuk: (a) kesesuaian dengan distribusi setengah normal; (b) bahwa kontras terstandarisasi dan panjang cabang tidak berkorelasi negatif, menyiratkan panjang cabang mungkin besar secara sewenang-wenang dan (c) variasi sifat yang diamati diprediksi oleh struktur filogenetik di bawah model gerak Brown dari evolusi sifat (Cooper et al., 2016 ). Karena PIC memerlukan rata-rata nol, analisis korelasi dihitung melalui titik asal (Garland, 1992 ). Kami mengurangi kesalahan tipe I dengan mengoreksi nilai- p menggunakan False Discovery Rate (FDR) (Benjamini & Hochberg, 2000 ). Metode koreksi ini memungkinkan nilai ambang batas untuk mendeklarasikan signifikansi meningkat secara monoton, mengurangi kemungkinan kehilangan hasil yang benar-benar signifikan relatif terhadap ambang batas yang tetap dan sangat konservatif termasuk Bonferroni (Pike, 2011) .). Analisis dilakukan di R.3.5.2 (Tim Inti R, 2020 ).
3 HASIL
3.1 Kovariansi sifat-sifat ekonomi sumber daya
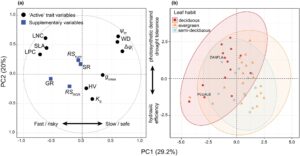
Terdapat kovariasi signifikan di antara ciri-ciri yang berkaitan dengan trade-off cepat-berisiko/lambat-aman, dengan dua sumbu PCA pertama mencakup 49% varians (Gambar 1 ) dan 5 sumbu yang diperlukan untuk menjelaskan 82% (Lampiran S5 ). Komponen utama pertama (PC1) menjelaskan 29,2% varians dan menyatakan trade-off cepat-berisiko/lambat-aman. Nilai PC1 yang lebih rendah berhubungan dengan permintaan fotosintesis yang lebih tinggi termasuk SLA ( ρ = −0,61), kandungan nitrogen daun ( ρ = −0,63) dan kandungan fosfor daun ( ρ = −0,72) sementara nilai yang lebih tinggi termasuk ψ m ( ρ = 0,61), Δ ψ l ( ρ = 0,68), kerapatan kayu ( ρ = 0,60) dan g smax ( ρ = 0,35) menandakan spesies dengan toleransi kekeringan yang lebih besar dan pertukaran gas musim kemarau (Gambar 1 ). Sebaliknya, PC kedua (PC2) menjelaskan 20% varians yang menggabungkan permintaan fotosintesis dan toleransi kekeringan dengan efisiensi transportasi air yang lebih rendah. Nilai PC2 yang lebih tinggi mencakup ψ m ( ρ = 0,64), kerapatan kayu ( ρ = 0,55), kandungan nitrogen daun ( ρ = 0,62) dan SLA ( ρ = 0,45) sementara K S ( ρ = −0,42) menandai nilai yang lebih rendah. PC ketiga (PC3) menjelaskan 13,7% varians dan menyelaraskan spesies di sepanjang sumbu permintaan air dan pertukaran gas musim kemarau, dengan nilai tinggi terkait dengan nilai huber yang tinggi ( ρ = 0,80) dan g smax musim kemarau yang rendah ( ρ = −0,53). Akhirnya, 10% varians dijelaskan oleh PC keempat (PC4) dengan nilai tinggi yang ditandai oleh K S yang tinggi ( ρ = 0,66) dan g smax musim kemarau yang rendah ( ρ = −0,48). Kebiasaan daun berkorelasi lemah dengan PC1 ( R 2 = 0,25; p = < 0,01).
3.2 Korelasi antara sifat-sifat sumber daya ekonomi, pertumbuhan dan kelangsungan hidup pada usia dewasa
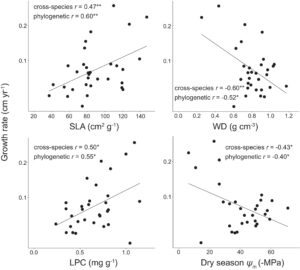
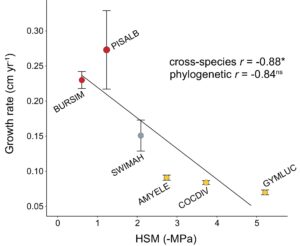
Beberapa sifat yang berkaitan dengan ekonomi karbon dan air dikaitkan dengan pertumbuhan spesies. Hubungan antara produktivitas daun dan pertumbuhan diilustrasikan oleh korelasi positif sedang antara laju pertumbuhan dan SLA ( r = 0,47; p < 0,05; PIC r = 0,60; p < 0,001) dan kandungan fosfor daun ( r = 0,50; p < 0,05; PIC r = 0,55; p < 0,01; Gambar 2 ; Tabel 2 ). Ada tanda-tanda korelasi positif antara laju pertumbuhan dan kandungan nitrogen daun, meskipun ini tidak signifikan pada tingkat lintas spesies ( r = 0,36; p = 0,07; PIC r = 0,46; p < 0,05). Ada bukti bahwa strategi pertumbuhan terkait dengan toleransi kekeringan. Korelasi negatif sedang hingga kuat diamati antara laju pertumbuhan dan kepadatan kayu ( r = −0,60; p < 0,001; PIC r = −0,51; <0,05) bersama dengan laju pertumbuhan dan ψ m ( r = −0,43; p < 0,05; PIC r = −0,40; p < 0,05; Gambar 2 ). Kemungkinan hubungan antara pasokan air dan pertumbuhan ditunjukkan oleh korelasi negatif PIC hanya antara laju pertumbuhan dan nilai huber ( r = −0,42; p = 0,07; PIC r = −0,54; p < 0,05). Tidak ada indikasi bahwa tingkat kelangsungan hidup terkait dengan salah satu sifat ekonomi sumber daya kami di tingkat lintas spesies atau PIC, bahkan sebelum koreksi nilai signifikansi FDR. Lebih jauh, tidak ada bukti langsung untuk trade-off pertumbuhan vs. kelangsungan hidup yang diamati ( r = −0,02; p = 0,85; PIC r = −0,02; p = 0,92; Tabel 2 ). Margin keamanan hidrolik menunjukkan korelasi negatif yang kuat dengan laju pertumbuhan ( r = −0,88, p < 0,05) tetapi korelasi ini tidak signifikan pada tingkat PIC ( r = −0,84; p = 0,07; Tabel 2 ; Gambar 3 ) karena ukuran sampel yang kecil (6 spesies), menghasilkan margin kesalahan yang lebih tinggi dan banyak koefisien korelasi kuat yang secara statistik tidak signifikan.
| Sifat | Persyaratan Layanan | LNC | LPC | WD 1 | ψ m | Δ ψ l | K adalah sebuah | TTV | g kecil maksimal a | HSM | GR | Bahasa Indonesia: SR | Nomor RS | RSUD |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Ekonomi sumber daya | ||||||||||||||
| Persyaratan Layanan | 0.62*** | 0,48** | -0,40* | 0,08 | -0,17 | -0,26 | -0,07 | -0,34 | -0,82 | 0,60*** | 0,15 | 0.20 | 0.44 | |
| LNC | 0,65*** | 0,58*** | -0,05 | 0,45* | 0,03 | -0,50 | -0,36 | -0,10 | -0,32 | 0,46* | 0,08 | 0.72*** | -0,09 | |
| LPC | 0,48* | 0,74*** | -0,31 | 0.18 | 0.17 | -0,59* | -0,56* | -0,03 | -0,20 | 0,55** | 0,03 | 0,45 | 0.14 | |
| WD sebuah | -0,29 | -0,11 | -0,35 | 0,42* | 0.10 | 0.28 | 0.54 | 0.13 | 0,92* | -0,52* | -0,35 | 0.26 | -0,48* | |
| ψ m | 0,03 | -0,21 | -0,28 | 0,46* | 0,57*** | -0,10 | -0,06 | -0,10 | -0,80 | -0,40* | 0,07 | 0,50* | -0,29 | |
| Δ ψ l | -0,22 | -0,17 | -0,13 | 0.37 | 0,57* | -0,46 | -0,28 | 0.18 | 0.67 | -0,23 | 0,08 | 0.44 | -0,28 | |
| Bahasa Inggris | 0.16 | -0,16 | -0,18 | 0,01 | -0,47 | -0,51 | 0,51* | -0,38 | -0,20 | -0,16 | -0,08 | -0,24 | 0,63** | |
| TTV | 0,03 | -0,11 | -0,44 | 0,25 | 0.14 | -0,01 | 0.22 | -0,60** | -0,21 | -0,54* | -0,04 | -0,16 | 0.40 | |
| g kecil maksimal a | -0,15 | -0,06 | -0,19 | 0.17 | 0.17 | 0.24 | -0,21 | -0,48 | -0,15 | 0,09 | 0.33 | 0,01 | -0,63** | |
| HSM | -0,67 | -0,37 | -0,19 | 0,90* | -0,77 | 0.37 | -0,37 | 0.51 | -0,17 | -0,84 | -0,32 | 0.19 | -0,31 | |
| Pertumbuhan dan kelangsungan hidup orang dewasa | ||||||||||||||
| GR | 0,47* | 0.36 | 0,50* | – 0,60** | -0,43* | -0,35 | 0.14 | -0,42 | -0,03 | -0,88* | -0,02 | 0.34 | 0.13 | |
| Bahasa Indonesia: SR | 0.28 | 0.32 | -0,03 | -0,02 | 0.12 | 0,05 | 0,01 | -0,01 | 0,05 | -0,05 | -0,02 | 0,52** | -0,01 | |
| Kinerja tunas kembali | ||||||||||||||
| Nomor RS | 0,09 | 0.31 | 0.28 | 0.00 | 0.10 | 0,15 | 0,03 | 0.41 | 0.00 | -0,11 | 0.37 | 0.12 | -0,25 | |
| RSUD | 0.24 | 0.23 | 0,54* | -0,48* | -0,33 | -0,23 | 0,49* | 0.18 | -0,43* | -0,32 | 0,41* | -0,01 | 0.10 | |
Catatan : Perbandingan lintas spesies standar berada di bawah diagonal dan korelasi filogenetik berdasarkan kontras independen berada di atas diagonal. Spesies N = 19–44 kecuali batas keamanan hidrolik (HSM) yang mencakup enam spesies. Koefisien yang dicetak tebal menunjukkan hubungan yang signifikan. Nilai- p korelasi : *<0,05, **<0,01, ***<0,001. Beberapa perbandingan dikoreksi menggunakan nilai- p yang disesuaikan FDR (Benjamini & Hochberg, 2000 ).
sebuah Log yang berubah.
3.3 Korelasi antara karakteristik ekonomi sumber daya dan kinerja perkecambahan
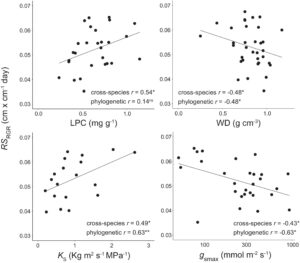
Kami menemukan bukti bahwa kinerja perkecambahan spesies selaras dengan trade-off cepat-berisiko/lambat-aman (Tabel 2 ). Konsentrasi nutrisi daun yang lebih tinggi secara konsisten menghasilkan perkembangan perkecambahan yang lebih besar: kandungan fosfor daun berhubungan cukup dengan laju pertumbuhan relatif perkecambahan ( r = 0,54, p < 0,05; PIC r = 0,14; p = 0,50) meskipun hubungannya tidak independen secara filogenetik (Gambar 4 ). Nilai yang lebih tinggi dari perkembangan tunas kembali juga terdeteksi di antara spesies dengan strategi penghindaran kekeringan dan toleransi struktural yang lebih lemah terhadap stres kekeringan: musim kemarau g smax berhubungan negatif secara moderat dengan laju pertumbuhan relatif tunas kembali ( r = −0,43, p < 0,05; PIC r = −0,63; p < 0,01; Gambar 4 ) sedangkan kepadatan kayu berhubungan cukup dengan laju pertumbuhan relatif tunas kembali ( r = −0,48, p < 0,05; PIC r = −0,48; p < 0,05; Gambar 4 ). Ada juga hubungan positif antara perkembangan tunas kembali dan efisiensi hidrolik, dimana laju pertumbuhan relatif tunas kembali berhubungan cukup positif dengan K S ( r = 0,49, p < 0,05; PIC r = 0,63; p < 0,01; Gambar 4 ). Secara keseluruhan, spesies dengan pertumbuhan cepat memiliki laju pertumbuhan relatif tunas yang lebih besar ( r = 0,41, p < 0,05; PIC r = 0,13; p = 0,55) meskipun temuan ini tidak terlepas dari sejarah filogenetik. Tidak ada sifat ekonomi sumber daya maupun pertumbuhan dewasa atau kelangsungan hidup yang terkait dengan jumlah tunas, meskipun ada indikasi spesies dengan kandungan nitrogen daun yang lebih tinggi memulai lebih banyak tunas ( r = 0,36, p = 0,09; PIC r = 0,72; p < 0,001; Tabel 2 ).
4 DISKUSI
Dalam studi ini, kami menyelidiki hubungan antara strategi fungsional dengan pertumbuhan dan kelangsungan hidup spesies dewasa dan kinerja perkecambahan di hutan kering tropis Karibia yang dipengaruhi oleh berbagai gangguan. Kami menemukan bahwa spesies pohon dapat diselaraskan di sepanjang trade-off fungsional cepat-berisiko vs. lambat-aman, di mana toleransi kekeringan dan penggunaan sumber daya menjelaskan sebagian besar variasi sifat. Khususnya, sementara toleransi kekeringan menunjukkan hubungan dengan tingkat pertumbuhan, dampaknya pada kelangsungan hidup spesies tidak signifikan. Selain itu, kami mengamati bahwa kinerja perkecambahan berkorelasi dengan strategi penggunaan sumber daya yang tinggi, yang menunjukkan bahwa produktivitas daripada persistensi sebagian besar memediasi proses ini di TDF Karibia. Di bawah ini, kami membahas hasil ini dan implikasinya untuk memahami resistensi dan ketahanan hutan tropis terhadap gangguan.
4.1 Cepat-berisiko versus lambat-aman merupakan sumbu variasi dominan di antara spesies
Untuk mendukung hipotesis pertama kami, sumbu PCA primer menyusun spesies di sepanjang kontinum cepat-berisiko vs. lambat-aman. Sementara SLA, LNC, dan LPC yang tinggi mencirikan permintaan fotosintesis yang kuat di antara banyak spesies, kontrol stomata musim kemarau yang ketat dan toleransi yang lebih rendah terhadap kekeringan menggarisbawahi kerentanan mereka terhadap emboli yang disebabkan oleh kavitasi. Sebaliknya, jaringan toleran kekeringan dari spesies yang lambat-aman ditambah dengan sifat sumber daya yang konservatif memungkinkan pertukaran gas yang lebih liberal ketika ketersediaan air menurun. Kompromi serupa yang disorot oleh analisis PIC kami (Tabel 2 ) menunjukkan bahwa mereka tidak terkait dengan sejarah evolusi spesies yang sama. Hubungan antara ekonomi karbon, strategi ketahanan kekeringan, dan keamanan hidrolik bersama-sama menunjukkan sumbu utama variasi fisiologis pohon dalam TDF ini dan memperkuat tren yang konsisten di seluruh bioma hutan tropis (Brodribb et al., 2002 ). Namun, varians yang dijelaskan oleh PC1 (29,2%) mengisyaratkan variasi yang luas dalam sindrom sifat dan keterbatasan generalisasi di seluruh taksa. Sementara PC2 dan PC3 (gabungan varians = 33,7%) juga menghubungkan hidraulik dengan ekonomi C, kombinasi cepat/lambat dan berisiko/aman tumpang tindih secara signifikan. Misalnya, spesies menggabungkan sifat daun yang lebih cepat (yaitu SLA atau LNC yang lebih tinggi) dengan sistem hidraulik yang kurang efisien (yaitu K S yang lebih rendah ) di sepanjang PC2, yang menunjukkan bahwa tanaman ini menggunakan mekanisme lain untuk mempertahankan margin keamanan hidraulik yang positif; misalnya, indeks luas daun yang lebih rendah, akses ke sumber air (misalnya akar yang dalam, kapasitansi tinggi), kemampuan untuk meningkatkan ketahanan terhadap emboli ( P 50 ) dan decoupling atmosfer (misalnya gugurnya daun atau kontrol stomata) telah terbukti terjadi pada pohon-pohon tropis (Oliveira et al., 2021 ; Taylor & Eamus, 2008 ). Selain itu, spektrum luas dalam strategi penggunaan sumber daya telah menjadi ciri komunitas TDF lainnya (misalnya Werden et al., 2018 ).
4.2 Spektrum cepat-berisiko/lambat-aman dihubungkan dengan kinerja orang dewasa
Pertumbuhan tetapi bukan kelangsungan hidup dikaitkan dengan beberapa sifat yang mencirikan spektrum cepat-berisiko/lambat-aman, hasil yang mengejutkan mengingat pertumbuhan dan kematian merupakan trade-off riwayat hidup yang terkenal di hutan tropis. Pertumbuhan cepat cenderung berinvestasi dalam transportasi air yang efisien dan pengembalian jangka pendek yang lebih tinggi, membantu mengimbangi biaya produksi daun yang kaya sumber daya dan padat nutrisi. Namun, strategi seperti itu sering kali menghasilkan risiko kematian yang lebih tinggi. Tidak jelas apakah tingkat kelangsungan hidup spesies yang dihitung dari ukuran sampel yang lebih besar akan memengaruhi hasil meskipun waktu generasi yang lama menimbulkan tantangan untuk mencatat kematian dalam komunitas yang tumbuh kembali. Subset spesies yang lebih kecil ( N = 6; Gambar 3 ) menunjukkan HSM yang lebih rendah untuk pertumbuhan cepat dibandingkan dengan pertumbuhan lambat, hubungan yang terlihat di tempat lain di daerah tropis (Markesteijn et al., 2011 ). HSM yang berkurang menyiratkan risiko disfungsi xilem yang lebih tinggi jika terjadi kekeringan hebat, yang menunjukkan pertumbuhan cepat lebih rentan terhadap kematian akibat kekeringan (Delzon, 2015 ). Mungkin tidak mengherankan, banyak dari tanaman cepat tumbuh kami mengadopsi strategi penghindaran kekeringan termasuk kontrol stomata yang ketat dan peluruhan daun mungkin untuk mengurangi kerentanan ini. Pertukaran HSM-pertumbuhan baru-baru ini diusulkan sebagai sumbu utama variasi sifat hidrolik di daerah tropis, dan penyertaannya dalam model menunjukkan hutan mungkin lebih rentan terhadap kekeringan daripada yang diperkirakan sebelumnya (diulas oleh Oliveira et al., 2021 ). Namun, melewati batas aman mungkin tidak memiliki konsekuensi fatal yang sama untuk pohon yang tumbuh kembali dibandingkan dengan yang tidak tumbuh. Misalnya, kekeringan dapat menyebabkan kematian pemimpin terminal tanpa mempengaruhi sistem akar secara parah yang mengarah pada perkembangan batang baru saat tunas yang tidak aktif tumbuh kembali (Zeppel et al., 2015 ). Dengan demikian, pertumbuhan kembali dapat menantang cara kita mengkarakterisasi kematian pohon dan pemulihan biomassa melalui kerangka kerja pertumbuhan HSM dan dengan demikian, menunjukkan perlunya penilaian menyeluruh tentang peran HSM dalam ketahanan hutan tropis di mana pertumbuhan kembali adalah hal yang umum.
4.3 Perkecambahan yang kuat berhubungan dengan produktivitas daripada persistensi
Resprouting diamati pada semua kecuali dua spesies, membuat perbedaan biner (resprouter/nonresprouter) tidak cukup untuk menangkap variasi yang berarti. Sebaliknya, memeriksa kelimpahan dan tingkat pertumbuhan resprout memberikan wawasan yang lebih dalam tentang perbedaan interspesifik dan strategi fungsional. Sejalan dengan hipotesis ketiga kami, kinerja resprouting yang lebih besar diamati di antara spesies yang berisiko cepat meskipun strategi aman-lambat dikaitkan dengan sifat persistensi (sensu Reich, 2014 ). Secara khusus, spesies dengan pertumbuhan resprout yang lebih cepat berkorelasi dengan pertumbuhan dewasa yang cepat, sifat yang membutuhkan cahaya, peningkatan efisiensi transportasi air, dan pemanfaatan strategi penghindaran kekeringan. Spesies dengan tunas yang tumbuh lebih lambat cenderung menjadi penanam dewasa yang lebih lambat, mungkin karena mereka mengalokasikan sumber daya yang sesuai untuk toleransi kekeringan yang lebih besar (misalnya jaringan daun yang lebih kuat, WD yang lebih tinggi). Meskipun rasio akar:tunas tinggi (~1:1) yang diamati pada tingkat komunitas di lokasi studi kami (Murphy & Lugo, 1986 )—yang menunjukkan bahwa akar kemungkinan berkontribusi pada penyimpanan dan perolehan sumber daya di antara banyak spesies—hubungan antara strategi penghindar kekeringan dan perkecambahan cepat tetap ada. Ini menunjukkan bahwa, bahkan dengan rasio akar:tunas yang tinggi, penghindar kekeringan dapat menyimpan lebih banyak sumber daya secara keseluruhan relatif terhadap penolong kekeringan untuk bertahan hidup pada periode tanpa daun dan bahan bakar regenerasi. Selain itu, perkecambahan yang menghindari kekeringan kemungkinan memiliki SLA yang lebih tinggi, yang memungkinkan perkecambahan daun yang cepat untuk mendukung pertumbuhan kembali. Meskipun korelasi positif ada di antara laju pertumbuhan tunas cepat, LPC, dan GR di antara spesies, kurangnya signifikansi pada tingkat PIC (Gambar 4 ) menyiratkan pengaruh nenek moyang yang sama, yaitu spesies yang terkait erat (misalnya, Polygonaceae, Nyctaginaceae dan Fabaceae; Gambar S1 ) yang dominan di hutan ini. Spesies ini memiliki ciri-ciri yang terbukti mendukung pertumbuhan di hutan tropis yang kekurangan nutrisi, seperti asosiasi ektomikoriza dan fiksasi N (Murphy & Lugo, 1986 ; Soong et al., 2020 ).
Jumlah tunas kembali yang dipancarkan tidak berhubungan dengan sifat ekonomi sumber daya dan mungkin lebih baik dijelaskan oleh bagaimana spesies merespons perubahan keseimbangan hormon yang disebabkan oleh gangguan. Misalnya, angin kencang dan kekeringan mengubah produksi etilen dan rasio auksin terhadap sitokinin dengan cara yang mendorong perkembangan tunas lateral (Kurepa & Smalle, 2022 ). Kurangnya penelitian serupa di daerah tropis telah membuat identifikasi tren umum antara sifat sumber daya dan kinerja tunas kembali menjadi tantangan. Lebih jauh, penelitian yang ada sering kali bertentangan dalam kesimpulan mereka bahwa tunas kembali yang kuat pasca-gangguan telah dikaitkan dengan spesies yang tumbuh lambat (misalnya Poorter, Kitajima, et al., 2010 ; Poorter, McDonald, et al., 2010 ; Zimmerman et al., 1994 ) dan cepat (misalnya Curran et al., 2008 ; Paz et al., 2018 ). Ada harapan umum bahwa kinerja resprouting yang lebih besar akan ditemukan di antara spesies dengan cadangan karbohidrat non-struktural yang besar (NSC; Bond & Midgley, 2001 ). Stok NSC yang besar telah terdeteksi di banyak spesies TDF berbatang besar dan berkayu ringan (Paz et al., 2015 , 2018 ) namun, cadangan NSC bawah tanah yang lebih besar juga telah ditemukan pada spesies berbatang lebat yang tumbuh lebih lambat (Signori-Müller et al., 2022 ). Pada akhirnya, berbagai metodologi yang diadopsi oleh penelitian ini, khususnya berbagai metrik resprouting yang terkait dengan kinerja resprouting, menimbulkan tantangan yang signifikan untuk menarik kesimpulan menyeluruh dan menekankan perlunya prosedur standar untuk maju.
Sejauh pengetahuan kami, ini adalah studi pertama di hutan tropis yang menggunakan serangkaian sifat hidrolik yang komprehensif dalam menghubungkan resprouting pasca-gangguan dengan strategi respons kekeringan. Ekosistem Mediterania yang rawan kebakaran telah menghasilkan banyak studi serupa tetapi telah difokuskan pada resprouting sebagai sifat biner (yaitu resprouter vs. nonsprouter). Di sana, resprouter secara konsisten memiliki kayu yang lebih ringan, daun yang kurang kuat dan toleransi kekeringan yang lebih rendah dan resistensi kavitasi (Hernández et al., 2011 ; Pratt et al., 2007 ; Vilagrosa et al., 2014 ) karena struktur akar yang ada menyangga spesies resprouting dari stres kekeringan yang mungkin terjadi di lingkungan pasca-kebakaran yang terbuka. Dalam TDF Karibia, interaksi antara kekeringan dan angin kemungkinan menentukan keberhasilan relatif resprout (yaitu kelangsungan hidup, rekrutmen) dan dengan demikian, spesies yang cepat-berisiko/lambat-aman dapat menempati relung regenerasi yang berbeda. Misalnya, ciri spesies lambat paling cocok untuk tumbuh kembali sebagai respons terhadap kekeringan karena sensitivitas stomata yang lebih rendah, toleransi kekeringan, dan HSM yang lebar mempertahankan penangkapan sumber daya sambil menjadi penyangga terhadap kegagalan hidrolik—asalkan mereka mempertahankan simpanan karbohidrat nonstruktural (NSC) yang cukup. Meskipun kami tidak menilai kelangsungan hidup tunas kembali, hubungan negatif yang diamati antara RS RGR dan WD, yang sangat bervariasi dengan HSM, menyajikan bukti tidak langsung bahwa spesies yang tumbuh kembali lebih lambat cenderung tidak mengalami emboli katastrofik. Selain itu, batang dan akar spesies lambat tropis dapat memiliki karbohidrat nonstruktural yang jauh lebih tinggi (Signori-Müller et al., 2022 ), mediator penting respons stres kekeringan (Tomasella et al., 2020 ). Kepastian apa pun di sini memerlukan data tentang penyimpanan NSC di akar dan kedalaman perakaran—dua komponen penting namun sebagian besar belum terselesaikan di antara spesies pohon tropis. Secara keseluruhan, kelangsungan hidup tunas kembali yang lebih tinggi di antara spesies lambat juga telah diamati di hutan Neotropis lainnya (Corrêa Scalon et al., 2020 ; Poorter, Kitajima, et al., 2010 ; Poorter, McDonald, et al., 2010 ).
Namun, tunas kembali dari spesies yang berisiko cepat dapat berkinerja lebih baik setelah peristiwa badai karena sifat mereka memungkinkan mereka untuk sepenuhnya mengeksploitasi kondisi sumber daya pasca badai (Paz et al., 2018 ; Van Bloem et al., 2003 ). Kinerja tunas kembali juga dapat dibantu di antara spesies yang berisiko cepat setelah peristiwa badai dengan tingkat keparahan rendah jika (a) pohon memprioritaskan remobilisasi sumber daya di atas tanah daripada di bawah tanah, seperti yang diamati di pesisir Afrika Selatan (Nzunda et al., 2008 ). Ini mungkin akan mendukung spesies peluruh kayu lunak berkayu ringan yang cenderung memiliki kolam penyimpanan di atas tanah yang lebih besar (karena persyaratan selama musim dorman) dan (b) jalur yang lebih pendek menuju subsidi fotosintesis untuk tunas kembali yang tumbuh lebih cepat, yang berpotensi mengimbangi setiap pertumbuhan vs. pertukaran penyimpanan sumber daya (Bond & Midgley, 2001 ). Khususnya, di hutan kering Puerto Rico, kerapatan kayu tidak berkorelasi dengan hilangnya biomassa setelah Badai Georges (Van Bloem et al., 2004 ), sebuah pola yang juga diamati dalam penelitian lain (Asner & Goldstein, 1997 ; Putz et al., 1983 ; Zimmerman et al., 1994 ). Ini menunjukkan bahwa sementara kerapatan kayu dapat menunjukkan dukungan struktural, secara umum itu adalah prediktor yang buruk dari kerusakan badai, dan bahwa faktor-faktor lain—seperti remobilisasi dan penyimpanan sumber daya di atas tanah—dapat memainkan peran yang lebih signifikan dalam menentukan keberhasilan resprouting setelah gangguan. Pada akhirnya, hubungan antara resprouting dan strategi cepat-berisiko/lambat-aman dalam TDF bertentangan dengan gagasan bahwa spesies lambat yang cenderung mengembangkan resprouts dengan kecepatan lambat akan ‘memprioritaskan’ keamanan hidrolik di atas pertumbuhan luas penampang (lihat Paz et al., 2018 ) karena kekeringan merupakan stresor kronis dalam sistem ini. Selain itu, jika kekeringan meningkat atau ketersediaan air menjadi kurang dapat diprediksi seperti yang ditunjukkan oleh skenario perubahan iklim di masa mendatang (Taylor et al., 2018 ) maka spesies yang tumbuh lambat dan aman mungkin memiliki posisi yang lebih baik untuk bertahan hidup di ekosistem hutan ini.
Studi ini adalah salah satu yang pertama untuk memeriksa trade-off riwayat hidup yang terkait dengan kemampuan resprouting di antara spesies pohon tropis; namun, lebih banyak data diperlukan untuk menginformasikan prediksi respons hutan Neotropis terhadap perubahan rezim gangguan. Lebih jauh lagi, studi tambahan diperlukan untuk menangkap trade-off penting lainnya yang bertanggung jawab atas dinamika regenerasi, konsisten dengan spektrum cepat-berisiko/lambat-aman yang hanya menangkap ~30% dari variasi di antara spesies kita (PC1; Gambar 1 ). Misalnya, trade-off postur pohon-rekrutmen yang beroperasi ortogonal terhadap trade-off pertumbuhan-kelangsungan hidup telah terbukti secara akurat memprediksi perubahan struktural dan komposisi selama suksesi hutan tropis (Rüger et al., 2020 ) dan studi lain telah menyarankan hubungan antara postur pohon dan kinerja resprouting (Paz et al., 2015 , 2018 ). Selain itu, penelitian lebih lanjut harus mencakup bentuk-bentuk lain dari resprouting di atas tanah yang terjadi lebih jauh di batang (misalnya apikal, epikormik) yang sering kali memberikan kontribusi biomassa yang signifikan selama regenerasi TDF (Souza et al., 2021 ), jika tidak selalu pada kepadatan batang. Fokus kami pada resprouting hipogeal mendasari kesimpulan kami pada resprouting setelah peristiwa gangguan tinggi (misalnya kebakaran, penebangan habis). Baik angin maupun kekeringan umumnya tidak dianggap sebagai gangguan dengan tingkat keparahan tinggi di TDF dan dampaknya dapat mendukung resprouting di atas tanah melalui penyimpanan karbohidrat di atas tanah yang lebih besar (Nzunda et al., 2008 ) atau resprouting dari bawah pemimpin terminal.
5 KESIMPULAN
Kami menunjukkan bahwa cepat-berisiko/lambat-aman merupakan trade-off utama dalam menjelaskan variasi di antara spesies pohon dominan di Caribbean TDF. Trade-off ini dikaitkan dengan variasi dalam pertumbuhan dewasa dan kerentanan hidrolik, yang menyiratkan bahwa pertumbuhan cepat berada pada risiko kematian yang lebih besar akibat stres kekeringan, meskipun kami tidak mendeteksi trade-off pertumbuhan-kelangsungan hidup dalam data kami. Secara keseluruhan, temuan kami menunjukkan bahwa trade-off cepat-berisiko/lambat-aman merupakan mediator penting dari respons gangguan di Caribbean TDF. Meskipun bukan merupakan faktor dalam inisiasi tunas, spesies penghindar kekeringan cepat memang bertunas kembali dengan lebih kuat, sehingga menunjukkan strategi ekonomi hidrolik dan daun memang memengaruhi regenerasi pasca-gangguan. Berdasarkan temuan kami, kami mengusulkan bahwa dinamika bertunas kembali di hutan ini dipengaruhi oleh pergeseran keseimbangan antara dominasi spesies sifat cepat dan persistensi spesies sifat lambat dalam kaitannya dengan frekuensi relatif angin dan kekeringan.


