ABSTRAK
Memahami hubungan biologis antara populasi sangat penting untuk pengelolaan dan konservasi satwa liar. Studi genetik memainkan peran utama dalam mengkarakterisasi hubungan ini, tetapi biasanya memerlukan rezim pengambilan sampel yang terstratifikasi untuk menilai tingkat spasial dan kekuatan aliran gen, dan pengaruh relatif jenis kelamin dan ontogeni pada pola konektivitas. Namun, ini dapat menjadi tantangan dalam beberapa sistem studi, khususnya pada spesies laut besar seperti hiu, di mana studi genetik sering kali bergantung pada oportunistik dan/atau pengambilan sampel yang dilakukan pada skala spasial yang besar. Kami menunjukkan pentingnya pengambilan sampel terstratifikasi untuk mengidentifikasi struktur genetik yang sebelumnya tidak terdeteksi pada hiu macan ( Galeocerdo cuvier ) di lepas pantai Australia timur, tempat panmixia sebelumnya telah dilaporkan. Kami melakukan analisis genomik populasi pada 414 hiu macan, yang mewakili jantan dan betina dan tahap juvenil-subdewasa dan dewasa, dan 21 lokasi yang mencakup sekitar 3000 km dari Australia timur dan kawasan Indo-Pasifik. Mirip dengan penelitian sebelumnya, kami menunjukkan kurangnya struktur genetik keseluruhan di seluruh area pengambilan sampel; Namun, analisis kami menunjukkan bukti autokorelasi spasial dan penataan genetik lokal pada hiu macan betina muda-subdewasa. Hasil ini menunjukkan potensi pengaruh jenis kelamin dan ontogeni pada pola struktur genetik populasi dan konektivitas pada hiu macan Australia. Kami membahas temuan ini dalam konteks habitat penting yang mendukung populasi hiu macan dan risiko melebih-lebihkan kekuatan hubungan biologis di antara populasi hiu jika tidak ada pola pengambilan sampel yang tepat.
1 Pendahuluan
Memahami pola konektivitas di seluruh rentang spesies adalah fundamental untuk menginformasikan pengelolaan keanekaragaman hayati (Hirschfeld et al. 2021 ; Hohenlohe et al. 2021 ). Secara spesifik, memahami batas spasial populasi sangat penting untuk memperkirakan ukuran populasi lokal dan struktur demografi (Clark et al. 2024 ), mengidentifikasi habitat utama untuk perlindungan (Carr et al. 2017 ), dan memprediksi potensi jangkauan spasial dari peristiwa gangguan (Whiterod et al. 2016 ). Informasi ini juga penting untuk menilai ketahanan spesies terhadap perubahan lingkungan, karena pencampuran genotipe di seluruh gradien lingkungan dapat memengaruhi kemampuan populasi alami untuk beradaptasi dengan tantangan lingkungan baru (Hoffmann dan Sgrò 2011 ; Miller et al. 2020 ). Studi genetik memainkan peran sentral dalam mengkarakterisasi pola spasial konektivitas biologis antara populasi melalui perkiraan aliran gen dan struktur genetik. Namun, estimasi aliran gen dan struktur genetik di antara populasi dapat menyesatkan jika tidak ada pengambilan sampel spasial yang tepat atau saat menggabungkan hewan dengan jenis kelamin dan tahap kehidupan yang berbeda untuk tujuan analitis (Phillips et al. 2021 ). Untuk menghindari masalah ini, rezim pengambilan sampel berstrata, yang melibatkan pengambilan sampel berulang pada skala spasial yang berbeda dan lintas jenis kelamin dan tahap kehidupan, sering kali diperlukan untuk menguji bukti struktur genetik pada skala spasial lokal dan luas, dan untuk mengevaluasi pengaruh jenis kelamin dan ontogeni pada dinamika konektivitas (Waples 1998 ; Holmes et al. 2017 ; Phillips et al. 2021 ). Akibatnya, studi yang didasarkan pada rezim pengambilan sampel oportunistik berisiko melebih-lebihkan kekuatan konektivitas di seluruh rentang spesies.
Pengambilan sampel spesies yang sangat mobile dan terdistribusi luas dalam mode replikasi spasial (yaitu, pengambilan sampel transek) dan biologis (yaitu, lintas jenis kelamin dan tahap kehidupan) untuk studi genetik populasi pada dasarnya menantang. Misalnya, spesies hiu pelagis besar sering kali sangat dispersif, sering berpindah antara habitat pesisir, neritik, dan samudra, dan sulit untuk diambil sampelnya karena sifatnya yang mobile, sulit ditangkap, dan sering kali menyendiri (Hirschfeld et al. 2021 ; Phillips et al. 2021 ). Studi genetik populasi telah dilakukan pada sekitar 140 spesies hiu hingga saat ini, banyak di antaranya menunjukkan tingkat aliran gen yang tinggi dan kurangnya struktur genetik pada skala spasial yang luas baik di dalam maupun di antara cekungan laut (Tabel 1 ; Hirschfeld et al. 2021 ; Phillips et al. 2021 ). Namun, sebagian besar studi genetik pada hiu, khususnya spesies pelagis besar, bergantung pada oportunistik dan/atau pengambilan sampel yang dilakukan pada skala spasial yang besar (Tabel 1 ). Rezim pengambilan sampel seperti ini sering kali tidak memiliki sensitivitas untuk memeriksa kemungkinan autokorelasi spasial dan struktur genetik lokal (Whiterod et al. 2016 ; Schmidt-Roach et al. 2021 ; Bertram et al. 2022 ), dan untuk memperhitungkan perbedaan penyebaran antara jenis kelamin dan tahap kehidupan yang berbeda (Phillips et al. 2021 ). Akibatnya, ada kemungkinan bahwa hubungan antara populasi telah dilebih-lebihkan pada beberapa spesies hiu karena rezim pengambilan sampel yang tidak tepat, dan bahwa perbedaan dalam penyebaran dan struktur genetik antara jenis kelamin dan tahap kehidupan yang berbeda telah diabaikan. Informasi tersebut adalah kunci untuk menginformasikan manajemen yang bertujuan untuk mengidentifikasi dan melindungi habitat penting (misalnya, habitat pembibitan atau perlindungan; Barnett et al. 2019 ; Heupel et al. 2019 ; De Wysiecki et al. 2023 ) dan memahami faktor biologis penting yang mendukung kelangsungan hidup populasi lokal, seperti filopatri reproduksi, yang telah ditunjukkan pada banyak spesies hiu (Klein et al. 2019 ; Mourier dan Planes 2013 ; Tillett et al. 2012 ).
| Jenis | Cekungan samudra | Struktur dalam/antar cekungan samudra | Tes untuk struktur genetik lokal | Uji penyebaran berdasarkan jenis kelamin | Uji perbedaan ontogenetik | Referensi |
|---|---|---|---|---|---|---|
| Hiu macan ( Galeocerdo cuvier ) | Indo-Pasifik, Atlantik | Dalam = Tidak; Diantara = Ya/Tidak a | TIDAK | TIDAK | TIDAK | Holmes dkk. ( 2017 ), Pirog, Jaquemet, dkk. ( 2019 ), Sortir dkk. ( 2021 ), Bernard dkk. ( 2021 ) |
| Sevengill hidung lebar ( Notorynchus cepedianus ) | Atlantik Selatan, Oseania, Pasifik Timur | Dalam = Tidak; Diantara = Ya | TIDAK | TIDAK | TIDAK | Schmidt-Roach dan kawan-kawan ( 2021 ) |
| Pemburu paus perunggu ( Carcharhinus brachyurus ) | Indo-Pasifik | Dalam = Tidak; Diantara = Ya | TIDAK | Ya | TIDAK | Junge dan kawan-kawan ( 2019 ) |
| Hiu banteng ( Carcharhinus leucas ) | Hindia Barat, Pasifik, Atlantik | Dalam = Tidak; Diantara = Ya/Tidak a | TIDAK | Ya | TIDAK | Pirog, Ravigne, dkk. ( 2019 ) |
| Ikan sirip putih samudra ( Carcharhinus longimanus ) | India, Atlantik | Dalam = Tidak; Diantara = Tidak | TIDAK | TIDAK | TIDAK | Sreelekshmi dan kawan-kawan ( 2020 ) |
| Hiu paus ( Rhincodon typus ) | Indo-Pasifik, Atlantik Utara | Di dalam = Ya; Di antara = Ya | TIDAK | TIDAK | TIDAK | Vignaud dan kawan-kawan ( 2014 ) |
| Hiu rubah bermata besar ( Alopias superciliosus ) | Atlantik, India | Dalam = Tidak; Diantara = Tidak | TIDAK | TIDAK | TIDAK | Morales dan kawan-kawan ( 2018 ) |
| Ikan mako sirip pendek ( Isrus oxyrinchus ) | Indo-Pasifik, Atlantik Utara | Dalam = Tidak; Diantara = Tidak | TIDAK | Ya | TIDAK | Corrigan dan kawan-kawan ( 2018 ) |
| Hiu putih ( Carcharodon carcharias ) | Indo-Pasifik | Di dalam = Ya; Di antara = Ya | TIDAK | TIDAK | TIDAK | Blower dan kawan-kawan ( 2012 ) |
a Holmes et al. ( 2017 ) memberikan bukti adanya struktur genetik yang signifikan antara populasi hiu macan dari cekungan Samudra Atlantik dan Indo-Pasifik, tetapi tidak antara hiu dari cekungan Samudra Hindia atau Pasifik. Pirog, Jaquemet, et al. ( 2019 ) mendeteksi diferensiasi genetik yang lemah antara Samudra Hindia Barat dan Samudra Pasifik Barat. Bernard et al. ( 2021 ) dan Sort et al. ( 2021 ) menemukan diferensiasi genetik antara Atlantik dan Indo-Pasifik. Pirog, Ravigné, et al. ( 2019 ) menemukan diferensiasi genetik antara hiu banteng dari Atlantik Barat dan hiu banteng dari Samudra Pasifik Barat dan Hindia, tetapi tidak di dalam cekungan Samudra Hindia Barat dan Samudra Pasifik Barat.
Dalam studi ini, kami menyelidiki efek dari rezim pengambilan sampel pada pola struktur genetik pada hiu macan ( Galeocerdo cuvier ). Hiu macan adalah predator puncak laut dengan distribusi sirkumglobal yang meliputi perairan laut beriklim sedang dan tropis (Simpfendorfer et al. 2001 ; Dicken et al. 2017 ). Beberapa studi telah mengindikasikan adanya struktur genetik populasi yang signifikan antara populasi hiu macan dari cekungan Samudra Atlantik dan Indo-Pasifik, tetapi kurangnya struktur genetik keseluruhan dan potensi panmixia pada skala spasial yang luas dalam cekungan ini (Holmes et al. 2017 ; Pirog, Jaquemet, et al. 2019 ; Bernard et al. 2021 ; Sort et al. 2021 ). Namun, studi genetik hiu macan sebagian besar terbatas pada pendekatan pengambilan sampel nonstratifikasi (Carmo et al. 2019 ; Pirog, Jaquemet, et al. 2019 ; Bernard et al. 2021 ). Bahasa Indonesia: Sampai batas tertentu, studi telemetri mendukung gagasan panmixia dalam cekungan samudra karena bukti penyebaran jarak jauh pada beberapa hiu macan (Lea et al. 2015 ; Lipscombe et al. 2020 ). Namun, variabilitas tinggi dalam pola pergerakan, baik di dalam maupun di antara tahap kehidupan, terjadi di antara hiu macan yang ditandai di lokasi yang sama (Meyer et al. 2009 ; Fitzpatrick et al. 2012 ; Holmes et al. 2014 ; Werry et al. 2014 ; Ajemian et al. 2020 ; Barnett et al. 2022 ; Niella et al. 2022 ). Akibatnya, ada kemungkinan bahwa sejumlah kecil migran efektif per generasi bertanggung jawab untuk mendorong pola homogenitas genetik pada skala spasial yang luas dalam cekungan samudra, dan bahwa kekuatan konektivitas biologis di antara populasi hiu macan telah dilebih-lebihkan. Baru-baru ini, McClain et al. ( 2022 ) memberikan dukungan untuk argumen ini dengan menunjukkan struktur genetik yang signifikan di antara hiu macan muda-subdewasa di Samudra Atlantik barat laut, tetapi tidak ada struktur genetik di antara populasi dewasa. Temuan ini konsisten dengan studi telemetri dari wilayah tersebut, yang menunjukkan hiu subdewasa dan dewasa yang besar sangat mobile, dan hiu muda dan subdewasa kecil menunjukkan pola tempat tinggal (Sulikowski et al. 2016 ; Ajemian et al. 2020 ). Namun, karena keterbatasan pengambilan sampel, McClain et al. ( 2022 ) tidak dapat menguji penyebaran yang bias jenis kelamin atau mengukur tingkat spasial aliran gen, yang menyoroti perlunya pendekatan pengambilan sampel yang lebih komprehensif.
Di sini, kami membangun temuan McClain et al. ( 2022 ) dengan menilai ulang status genetik populasi hiu macan dari Australia timur dan kawasan Indo-Pasifik, di mana penelitian sebelumnya telah menunjukkan kurangnya struktur genetik di seluruh wilayah berdasarkan pada rezim pengambilan sampel geografis yang luas dan penggunaan penanda genetik modern (yaitu, polimorfisme nukleotida tunggal, SNP) dan tradisional (yaitu, mikrosatelit) (Bernard et al. 2016 ; Holmes et al. 2017 ). Sampel jaringan dari lebih dari 400 hiu dikumpulkan di 21 lokasi pengambilan sampel di sepanjang Australia timur dan kawasan Indo-Pasifik. Kami membandingkan pola struktur genetik di antara jenis kelamin dan tahap kehidupan menggunakan desain pengambilan sampel yang direplikasi secara spasial (mencakup gradien lintang ~3000 km) dan penanda SNP yang berasal dari sekuensing representasi genomik yang dikurangi. Hasil kami menyoroti pentingnya rejim pengambilan sampel hierarkis dan berulang, yang memberikan wawasan baru tentang kekuatan aliran gen dan konektivitas di antara hiu macan di wilayah tersebut. Secara khusus, hasil kami menunjukkan bias penyebaran yang berkaitan dengan jenis kelamin dan kemungkinan ontogeni, serta potensi residensi dan penataan genetik lokal. Temuan ini menantang gagasan panmixia pada hiu macan dari wilayah Indo-Pasifik dan menyoroti risiko melebih-lebihkan kekuatan konektivitas di antara populasi hiu jika tidak ada rejim pengambilan sampel yang tepat.
2 Metode
2.1 Pengambilan sampel
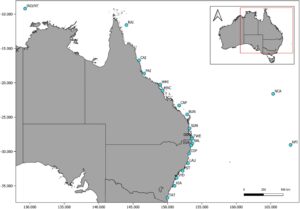
Sampel jaringan dikumpulkan dari 414 hiu macan antara tahun 2015 dan 2021, yang mewakili individu dengan berbagai jenis kelamin, kelas ukuran, dan 21 lokasi berbeda (minimal lima sampel per lokasi) yang mencakup lebih dari 3000 km Australia timur dari Merimbula (−36.9° S, 149.9° E) di selatan New South Wales hingga Pulau Raine (−11.6° S, 144° E) di Queensland Utara, dan Pulau Norfolk (−29° S, 167.9° E), serta Kaledonia Baru (−21.6° S, 165.4° E), dan Indonesia/Wilayah Utara (−9.2° S, 129.3° E; Gambar 1 ). Hiu ditangkap menggunakan metode kail dan tali (Lipscombe et al. 2020 ; Tate et al. 2021 ), atau pada drumline dan/atau jaring mitigasi hiu (Holmes et al. 2017 ). Hiu hidup ditandai secara fisik, dan semua metadata yang direkam mencakup lokasi penangkapan, jenis kelamin, dan ukuran tubuh. Jaringan otot dan/atau potongan sirip dikumpulkan dari setiap hiu, segera diawetkan dalam etanol, dan disimpan pada suhu −20°C hingga diperlukan untuk analisis genetik. Pendekatan pengambilan sampel yang direplikasi secara spasial yang digunakan di sini dirancang untuk tujuan pengujian bukti struktur genetik skala luas serta struktur genetik lokal dan autokorelasi spasial. Pengambilan sampel spasial yang direplikasi dari jantan dan betina, dan tahap kehidupan dewasa dan juvenil-subdewasa, juga disertakan untuk memungkinkan kami menguji bias jenis kelamin dan ontogenetik dari penyebaran dan struktur genetik di seluruh distribusi pengambilan sampel (Tabel 2 ; Tabel S1 ). Namun, pengambilan sampel kami bias terhadap hiu remaja-sub-dewasa dan mungkin disebabkan oleh pengumpulan sampel yang sebagian besar terjadi di habitat pesisir dekat pantai dan selektivitas alat tangkap (Holmes et al. 2012 ).
| Nama situs | Kode situs | Lintang | Garis bujur | Jumlah n | Remaja-sub-dewasa | Dewasa | ||
|---|---|---|---|---|---|---|---|---|
| Perempuan | Laki-laki | Perempuan | Laki-laki | |||||
| Pulau Raine | RAI | -11.58837 | Nomor telepon 144.03137 | 8 | 4 | angka 0 | 4 | angka 0 |
| Cairns | Bahasa Indonesia: CAI | -16.74665 | 145.87043 | 18 | 7 | 3 | 8 | angka 0 |
| Kepulauan Palm | PAI | -18.65251 | 146.59898 | 12 | 4 | 2 | 4 | 2 |
| Hari Minggu Whitsunday | SIAPA | -20.26174 | 148.93661 | 48 | 7 | 5 | 28 | 8 |
| Mackay | Bahasa Indonesia: MAC | -21.13224 | 149.29475 | 16 | 11 | 4 | 1 | angka 0 |
| Kelompok Capricorn | TOPI | -23.29548 | 151.69477 | 6 | 3 | angka 0 | 3 | angka 0 |
| Bundaberg | SANGGUL | -24.60315 | 152.85571 | 29 | 8 | 12 | 8 | 1 |
| Pantai Sunshine | MATAHARI | -26.64017 | 153.20399 | 23 | 16 | 2 | 5 | angka 0 |
| Pantai Emas | GOL | -27.94874 | 153.55313 | 6 | 2 | 3 | 1 | angka 0 |
| Kepala Wol | DUA KALI | -28.15654 | 153.5787 | 10 | 4 | 5 | 1 | angka 0 |
| Kota Ballina | BAL | -28.8329 | 153.6102 | 77 | 28 | 30 | 14 | 5 |
| Kepala Evans | EVA | -29.1104 | 153.4412 | 25 | 12 | 10 | 3 | angka 0 |
| Pelabuhan Coffs | COF | -30.2922 | 153.154 | 28 | 22 | 5 | angka 0 | 1 |
| Laurieton | Bahasa Indonesia: LAU | -31.71084 | 152.98571 | 6 | 4 | 1 | 1 | angka 0 |
| Pelabuhan Stephens | Waktu Standar Pasifik | -32.80953 | Nomor telepon 152.19961 | 27 | 4 | 5 | 9 | 9 |
| Kota Sydney | Daerah Administratif Khusus Sydney | -33.8438 | 151.322 | 10 | 2 | 2 | 1 | 5 |
| Kota Kiama | KIA | -34.99872 | Nomor telepon 151.02565 | 10 | 7 | 2 | 1 | angka 0 |
| Tatra | TAT | -36.7381 | 149.9972 | 5 | 4 | angka 0 | 1 | angka 0 |
| Indonesia/ NT | IND | -9.19671 tahun | 129.29598 | 3 | 2 | 1 | angka 0 | angka 0 |
| Kaledonia Baru | NCA | -21.61646 | 165.36407 | 23 | 9 | 1 | 12 | 1 |
| Pulau Norfolk | Dana Non-Finansial (NFI) | -29.04203 | 167.89909 | 18 | 1 | 1 | 12 | 4 |
a Indonesia/NT = 5 × jenis kelamin tidak diketahui, Whitsundays = 1 × jenis kelamin tidak diketahui.
2.2 Ekstraksi DNA dan Genotipe SNP
Total DNA genomik diekstraksi dari 10 hingga 15 mg jaringan dari setiap sampel oleh Diversity Arrays Technologies (DArT Pty Ltd., Canberra, Australia) menggunakan NucleoMag 96 Tissue Kit (Macherey-Nagel, Düren, Jerman) yang dipasangkan dengan NucleoMag SEP untuk memungkinkan pemisahan otomatis DNA berkualitas tinggi pada robot pengendali cairan Freedom Evo (TECAN, Männedorf, Swiss). Genotipe Single Nucleotide Polymorphism (SNP) kemudian dilakukan dengan menggabungkan metode pengurangan kompleksitas genom milik DArT dengan sekuensing generasi berikutnya (Wenzl et al. 2004 ; Sansaloni et al. 2011 ; Kilian et al. 2012 ). Pustaka kompleksitas genom yang dikurangi untuk G. cuvier dihasilkan termasuk langkah-langkah pencernaan dan ligasi DNA termasuk kombinasi enzim restriksi PstI/TaqI dengan urutan adaptor kode batang yang memungkinkan pemilihan amplifikasi fragmen restriksi melalui 30 siklus reaksi berantai polimerase (PCR) menggunakan primer DArT PstI (5′-GAT GGA TCC AGT GCA G-3′). Kondisi PCR mencakup denaturasi awal pada suhu 94°C selama 1 menit, diikuti oleh 30 siklus pada suhu 94°C selama 20 detik (denaturasi), 58°C selama 30 detik (annealing), dan 72°C selama 45 detik (ekstensi), dengan ekstensi akhir pada suhu 72°C selama 7 menit. Produk PCR yang diperkuat diurutkan pada platform HiSeq2500 (Illumina, San Diego, AS) menggunakan pengurutan baca tunggal selama 77 siklus, yang menghasilkan rata-rata 3448 juta pembacaan per sampel. Alur analitis DArT (Ren et al. 2015 ; Egea et al. 2017 ) selanjutnya digunakan untuk mendemultipleks urutan DNA dari sampel individual, menyaring urutan berkualitas buruk dengan skor Phred < 30, dan untuk memanggil genotipe SNP. Genotipe menghasilkan total 32.690 lokus SNP dari 407 hiu macan individu dengan rata-rata cakupan urutan 20x. Lokus SNP selanjutnya difilter menggunakan paket dartR versi 1.9.9.1 (Gruber et al. 2018 ), hanya mempertahankan satu SNP per tag, membuang yang sekunder, menerapkan ambang batas tingkat panggilan lokus dan individu hingga 80%, reproduktifitas 90%, dan frekuensi alel minor minimum 5%. Akhirnya, lokus SNP yang menyimpang dari ekspektasi Hardy–Weinberg ( p < 0,05) dihilangkan dari kumpulan data, dan ambang batas jarak hamming 0,2 diterapkan untuk mengendalikan paralog potensial. Setelah penyaringan, total 10.709 lokus SNP dari 407 hiu macan individu dan 21 lokasi pengambilan sampel tetap ada untuk analisis genomik populasi hilir.
2.3 Tes untuk Struktur Genetik Populasi Secara Keseluruhan
Frekuensi SNP di semua lokus diperbandingkan antara individu dari setiap lokasi untuk menentukan pola keseluruhan struktur genetik dan konektivitas populasi. Perangkat lunak hierfstat yang diimplementasikan dalam R (Goudet 2005 ) digunakan untuk menghitung ukuran global dan berpasangan dari diferensiasi populasi ( FST ; Weir dan Cockerham 1984 ) pada lokus SNP yang difilter. Analisis variasi molekuler (AMOVA) dilakukan dalam paket R poppr (Kamvar et al. 2014 ) dengan menggunakan FST berpasangan sebagai ukuran jarak, dan mempartisi variasi di antara lokasi sampel dan dalam lokasi sampel, dengan signifikansi ditentukan berdasarkan 999 permutasi. Analisis Diskriminan Komponen Utama (DAPC) dilakukan dengan menggunakan perangkat lunak adegenet (Jombart 2008 ; Jombart dan Ahmed 2011 ) yang diimplementasikan dalam paket R dengan menggunakan fungsi find clusters. Selain itu, paket analisis Bayesian STRUCTURE (Pritchard et al. 2000 ) digunakan untuk menguji keseluruhan struktur genetik populasi. STRUCTURE digunakan untuk mengidentifikasi jumlah klaster populasi yang berbeda, untuk menempatkan individu ke dalam klaster, dan untuk mengidentifikasi migran dan individu campuran menggunakan data genetik saja. Untuk menentukan jumlah klaster populasi ( K ), lima simulasi independen untuk K = 1–21 dengan 10.000 burn-in dan 100.000 iterasi data dijalankan. Analisis dilakukan menggunakan model campuran struktur populasi (yaitu, setiap individu mengambil beberapa fraksi genom mereka dari masing-masing populasi K ) dan frekuensi alel ditetapkan sebagai independen di antara populasi. K yang paling mungkin diperkirakan menggunakan Δ K Evanno (Evanno et al. 2005 ) dalam Structure Harvester (Earl dan von Holdt 2012 ). Terakhir, estimasi keterkaitan di antara semua individu dihitung menggunakan paket R SNPRelate (Zheng et al. 2012 ), dengan kategori keterkaitan (hubungan derajat pertama, kedua dan ketiga) disimpulkan dari koefisien ko-leluhur ( θ ) dan koefisien R 0 dan R 1 yang menyertainya (Tabel S2 ).
2.4 Uji Struktur Genetik Lokal dan Autokorelasi Spasial
Analisis autokorelasi spasial dilakukan dalam GenAlEx 6.51 (Peakall dan Smouse 2006 ), yang menyediakan pengujian penataan genetik lokal dan keterbatasan aliran gen di antara lokasi pengambilan sampel. Analisis dilakukan pada subset acak dari 1000 lokus SNP (karena keterbatasan komputasi program) dengan analisis dilakukan pada semua hiu yang digabungkan dan kemudian jenis kelamin (jantan dan betina) dan tahap kehidupan secara terpisah (remaja-subdewasa dan dewasa). Hiu betina dengan panjang total (TL) > 330 cm dan jantan > 300 cm dianggap dewasa, sedangkan yang lebih kecil diperlakukan sebagai remaja-subdewasa (sebagaimana per L 50 untuk populasi ini, dilaporkan dalam Holmes et al. 2015 ). Kelas jarak untuk analisis ini didasarkan pada opsi “ukuran sampel yang sama”, dengan 10.000 permutasi untuk menguji tingkat signifikansi dan menggunakan opsi uji “multi-pop”. Untuk setiap kelas, permutasi acak di lokasi spasial individu (10.000 permutasi) kemudian digunakan untuk menilai penyimpangan koefisien keterkaitan ( R ) dari 0. Kelas jarak dipilih sehingga berisi lebih dari 100 perbandingan berpasangan, memiliki indeks partisipasi > 50% dan koefisien variasi partisipasi < 1 (Hardy dan Vekemans 2002 ). Koefisien keterkaitan ( R ) dihitung untuk semua pasangan individu, dengan perbandingan berpasangan di delapan kelas jarak mulai dari 0 hingga 700 km. Penyimpangan dari 0 menunjukkan bahwa individu dalam kelas jarak tertentu secara signifikan lebih (nilai positif) atau kurang (nilai negatif) secara genetik mirip daripada yang diharapkan secara acak (Whiterod et al. 2016 ).
3 Hasil
3.1 Struktur Genetik Populasi Secara Keseluruhan
Analisis genomik populasi menunjukkan struktur genetik yang lemah tetapi signifikan di antara lokasi sampel (global F ST = 0,001, 95% CI: 0,001–0,002). Pola ini sebagian besar konsisten di antara hiu yang bervariasi menurut jenis kelamin dan tahap kehidupan, dengan analisis independen yang menunjukkan struktur genetik yang lemah tetapi signifikan pada hiu betina muda-subdewasa (global F ST = 0,004, 95% CI: 0,003–0,004), hiu jantan muda-subdewasa (global F ST = 0,009, 95% CI: 0,007–0,009), dan hiu betina dewasa (global F ST = 0,005, 95% CI: 0,004–0,006). Sebaliknya, kurangnya struktur genetik yang signifikan diamati pada jantan dewasa (global F ST = 0,000, 95% CI: 0,000–0,001), meskipun estimasi ini harus ditafsirkan dengan hati-hati karena keterbatasan pengambilan sampel. Secara keseluruhan, estimasi berpasangan F ST menunjukkan struktur genetik yang lemah antara lokasi pengambilan sampel, dengan hanya 55 dari 210 estimasi berpasangan yang berbeda secara signifikan dari nol (Tabel S2 ). AMOVA tidak menemukan bukti diferensiasi genetik yang signifikan antara lokasi sampel (0,12%, p > 0,001), dengan mayoritas varians dijelaskan oleh variasi genetik antara individu dalam lokasi (99,88%, p < 0,001). Demikian pula, analisis DAPC dan STRUCTURE menunjukkan kurangnya struktur genetik, keduanya mengidentifikasi satu klaster populasi ( K = 1, Δ K = 1; Gambar S1 dan S2 ). Akhirnya, estimasi kekerabatan di antara semua individu yang dihasilkan dalam paket SNPRelate menggunakan beberapa ukuran keterkaitan menunjukkan keterkaitan keseluruhan yang rendah di antara pasangan hiu. Dari total 82.621 ukuran keterkaitan berpasangan, > 99% diidentifikasi sebagai individu yang tidak berkerabat. Secara keseluruhan, hanya enam hubungan tingkat pertama (saudara kandung), lima hubungan tingkat kedua (saudara tiri), dan 16 hubungan tingkat ketiga (saudara seperempat) di antara pasangan hiu yang tercatat (Tabel S3 ). Individu yang berkerabat tidak dikaitkan dengan lokasi pengambilan sampel yang sama.
3.2 Struktur Genetik Lokal dan Autokorelasi Spasial
Meskipun bukti terbatas dari keseluruhan struktur genetik, analisis autokorelasi spasial menunjukkan penataan genetik lokal dan keterbatasan aliran gen. Koefisien keterkaitan dihitung untuk semua pasangan individu, yang melibatkan 80.200 perbandingan berpasangan di delapan kelas jarak, mulai dari 0 hingga 700 km. Autokorelasi spasial yang signifikan dan positif diamati hingga 100 km (Gambar S3 ), yang menunjukkan bahwa individu pada skala spasial ini lebih mirip secara genetik daripada yang diharapkan secara acak. Analisis terpisah yang dilakukan untuk jenis kelamin dan tahap kehidupan yang berbeda menunjukkan bahwa sinyal struktur genetik lokal didorong oleh hiu macan betina muda-subdewasa saja. Di sini, autokorelasi spasial yang signifikan dan positif terdeteksi hingga jarak 100 k berdasarkan 11.781 perbandingan berpasangan (Gambar 2 ). Hal ini selanjutnya didukung oleh pola autokorelasi spasial yang signifikan dan negatif pada 200 km, yang menunjukkan hiu pada skala spasial ini kurang mirip secara genetik daripada yang diharapkan dalam skenario perkawinan acak. Di luar 200 km, hiu macan betina muda-subdewasa tampaknya tidak lebih atau kurang mirip secara genetik daripada yang diharapkan secara kebetulan. Sebaliknya, analisis autokorelasi spasial dari hiu macan betina dewasa (6786 perbandingan berpasangan) tidak menunjukkan bukti signifikan dari penataan genetik lokal (Gambar 2 ). Kurangnya autokorelasi spasial juga diamati ketika hiu jantan muda-subdewasa (4371 perbandingan berpasangan) dianalisis secara terpisah, dengan pengecualian satu sinyal signifikan dan positif yang diamati pada 300 km. Namun, ini tidak mungkin bermakna secara biologis karena kurangnya struktur genetik pada kelas jarak yang lebih halus, dan tingkat kesalahan yang relatif tinggi di sekitar rata-rata berdasarkan sejumlah kecil perbandingan berpasangan untuk kelas jarak ini (94 perbandingan berpasangan). Akhirnya, kurangnya autokorelasi spasial juga diamati pada hiu jantan dewasa (630 perbandingan berpasangan), meskipun keluaran ini harus ditafsirkan dengan hati-hati karena ukuran sampel yang relatif kecil (Gambar 2 , Tabel 2 ).

Untuk menguji efek ukuran sampel pada daya untuk menyelesaikan tanda-tanda struktur genetik lokal, analisis diulang menggunakan dataset SNP hiu macan betina muda-subdewasa, subsampel 50 dan 100 hiu tiga kali secara acak dan mengulangi analisis autokorelasi spasial. Dalam semua kasus, ukuran sampel 50 gagal mendeteksi tanda-tanda struktur genetik lokal, sementara ukuran sampel 100 berhasil mendeteksi pola konsisten autokorelasi spasial yang signifikan dan positif hingga jarak 100 km (Gambar S4 ). Temuan-temuan ini memberikan keyakinan pada hasil kami yang menunjukkan kurangnya struktur genetik lokal pada hiu macan betina dewasa (117 individu) dan hiu macan muda-subdewasa jantan (94 individu) tetapi menunjukkan bahwa pengujian untuk struktur genetik lokal pada hiu macan jantan dewasa berdasarkan 36 individu harus diperlakukan dengan hati-hati.
4 Diskusi
Kami menunjukkan pentingnya replikasi rejim pengambilan sampel secara spasial dan biologis (yaitu, lintas jenis kelamin dan tahap kehidupan) saat mengkarakterisasi kekuatan aliran gen dan konektivitas populasi pada spesies hiu pelagis besar. Studi kami menunjukkan struktur genetik yang lemah pada hiu macan dari Australia timur dan kawasan Indo-Pasifik, yang mendukung temuan dari studi genetik sebelumnya yang menunjukkan kemungkinan keberadaan populasi panmiktik tunggal di kawasan tersebut (Bernard et al. 2016 ; Holmes et al. 2017 ). Namun, rejim pengambilan sampel berstrata kami di sepanjang pantai timur Australia memungkinkan deteksi autokorelasi spasial yang signifikan dan struktur genetik lokal pada hiu macan betina muda-subdewasa, yang menunjukkan potensi bias penyebaran antara jenis kelamin dan tahap kehidupan, dan kemungkinan filopatri reproduksi betina. Temuan ini memberikan wawasan baru tentang pengaruh jenis kelamin dan ontogeni pada pola struktur genetik populasi dan konektivitas pada hiu macan Australia timur, serta habitat penting yang mendukung populasi hiu macan. Secara keseluruhan, studi ini menyoroti pentingnya rezim pengambilan sampel yang bertingkat dan risiko melebih-lebihkan kekuatan hubungan biologis di antara populasi hiu jika tidak ada pendekatan pengambilan sampel yang tepat.
4.1 Bukti Struktur Genetik Skala Halus
Temuan kami konsisten dengan studi sebelumnya yang menunjukkan kurangnya struktur genetik secara keseluruhan di antara hiu macan dari Australia timur dan kawasan Indo-Pasifik (Bernard et al. 2016 ; Holmes et al. 2017 ). Studi pelacakan juga menunjukkan potensi migrasi jarak jauh (lebih dari ribuan kilometer) pada hiu macan dari wilayah ini, yang selanjutnya mendukung gagasan panmixia populasi (Lipscombe et al. 2020 ; Barnett et al. 2022 ). Temuan ini konsisten dengan studi genetik dan telemetri sebelumnya yang dilakukan di Samudra Atlantik, yang menunjukkan kemungkinan terjadinya panmixia populasi (Lea et al. 2015 ; Pirog, Jaquemet, et al. 2019 ; Sort et al. 2021 ; Hammerschlag et al. 2022 ). Namun, kekuatan aliran gen dan konektivitas dalam cekungan samudra masih belum pasti karena bias pengambilan sampel dan keterbatasan dalam sebagian besar studi genetik dan telemetri (Ferreira et al. 2015 ; Carmo et al. 2019 ; Pirog, Jaquemet, et al. 2019 ; Bernard et al. 2021 ; Hammerschlag et al. 2022 ). Studi telah menunjukkan bahwa pola homogenitas genetik dan panmixia dapat didorong oleh tingkat aliran gen yang rendah dan migrasi antargenerasi antara populasi (Wright 1931 ; Whiterod et al. 2016 ). Hal ini berpotensi berlaku untuk hiu macan mengingat tingkat variasi intraindividual yang tinggi dalam perilaku penyebaran, termasuk pola pergerakan skala besar dan terbatas, dan bukti potensi penyebaran yang bias jenis kelamin dan ontogenetik (Driggers et al. 2008 ; Meyer et al. 2009 ; Werry et al. 2014 ; Ajemian et al. 2020 ; Barnett et al. 2022 ; McClain et al. 2022 ; Niella et al. 2022 ). Di sini, kami menunjukkan penataan genetik skala halus pada hiu macan betina muda-subdewasa dari Australia timur. Temuan ini kemungkinan besar didorong oleh proses demografi daripada tekanan seleksi yang bervariasi secara spasial, karena lokus kandidat yang dipengaruhi oleh seleksi terarah sering kali bertahan dalam kelimpahan rendah dan memiliki sedikit efek pada pengujian untuk keseluruhan struktur genetik populasi (Holland et al. 2022 ; Sandoval-Castillo et al. 2018 ). Temuan ini mendukung temuan McClain et al. ( 2022 ) yang baru-baru ini melaporkan struktur genetik yang bergantung pada ukuran pada hiu macan dari Samudra Atlantik barat laut, di mana struktur genetik paling besar terjadi pada tahap kehidupan remaja dan subdewasa tanpa memandang jenis kelamin. Temuan kami didasarkan pada temuan McClain et al. ( 2022)) menunjukkan bahwa jenis kelamin dan ontogeni kemungkinan besar memengaruhi pola struktur genetik dan konektivitas populasi di seluruh rentang spesies.
Pendekatan pengambilan sampel kami memungkinkan kami untuk mendapatkan wawasan tentang pengaruh jenis kelamin dan ontogeni pada koneksi biologis pada hiu macan dari Australia timur dan kawasan Indo-Pasifik. Studi pelacakan sebelumnya dari kawasan yang sama telah menunjukkan tempat tinggal sepanjang tahun pada beberapa betina muda-subdewasa di beberapa bagian Australia timur dan Indo-Pasifik (Werry et al. 2014 ). Namun, sebagian besar studi telah melaporkan pergerakan betina muda-subdewasa sangat bervariasi, dengan beberapa individu melakukan perjalanan lebih dari seribu kilometer dalam ~30 hari dari peristiwa penandaan (Holmes et al. 2014 ; Meyer et al. 2009 ; Niella et al. 2022 ; Papastamatiou et al. 2013 ). Lebih jauh lagi, studi telemetri baru-baru ini menunjukkan tingkat tempat tinggal hiu macan bervariasi di antara individu dan kawasan di Australia timur (Niella et al. 2022 ). Secara khusus, Niella et al. ( 2022 ) melaporkan perbedaan perilaku antara hiu macan dari lokasi penandaan yang berbeda, di mana hiu dari garis pantai bagian utara menunjukkan tingkat residensi lokal yang lebih besar daripada hiu dari pantai tengah di mana pola pergerakannya lebih luas. Temuan kami menunjukkan potensi residensi pada hiu macan betina muda-subdewasa, pola spasial yang tampaknya konsisten di seluruh distribusi sampel kami di Australia timur. Namun, bukti variabilitas antarindividu yang substansial yang berkaitan dengan vagilitas dan residensi menunjukkan bahwa tanda-tanda autokorelasi spasial mungkin didorong oleh hanya sebagian kecil dari subpopulasi hiu macan betina muda-subdewasa. Penelitian lebih lanjut adalah bijaksana untuk membantu memvalidasi temuan ini dan untuk mengukur tingkat residensi pada hiu macan betina muda-subdewasa lebih akurat melalui studi pelacakan yang lebih komprehensif. Upaya tersebut diharapkan dapat memberikan wawasan baru tentang habitat penting yang mendukung hiu macan di Australia timur dan berpotensi lebih luas di seluruh kawasan Indo-Pasifik.
Sebaliknya, hasil kami menunjukkan kurangnya struktur genetik pada skala spasial lokal dan luas pada hiu macan dewasa jantan dan betina dan hiu macan remaja-subdewasa jantan. Meskipun hasil kami yang berkaitan dengan hiu macan dewasa jantan harus ditafsirkan dengan tingkat kehati-hatian tertentu karena keterbatasan pengambilan sampel, studi telemetri di Australia timur dan kawasan Indo-Pasifik (dan dari cekungan samudra lainnya) menunjukkan bahwa hiu macan dewasa jantan dan betina sangat dispersif (Holmes et al. 2014 ; Ferreira et al. 2015 ; Lipscombe et al. 2020 ). Sebaliknya, studi telemetri pada hiu macan remaja-subdewasa jantan kurang direplikasi, meskipun studi tersebut juga menunjukkan pergerakan lepas pantai yang luas dan kemampuan untuk melakukan penyelaman dalam hingga ~1 km (Lipscombe et al. 2020 ). Di Hawaii, Meyer et al. ( 2009 ) juga melaporkan bahwa hiu remaja memiliki jangkauan yang jauh lebih luas daripada hiu dewasa, mungkin didorong oleh perilaku menghindari predasi. Dengan demikian, masih sulit untuk menentukan konsistensi apa pun dalam pendorong pergerakan hiu macan, dan karena itu kami menyarankan agar penelitian di masa depan menggabungkan berbagai lini investigasi termasuk jenis kelamin, ontogeni, dan bahkan perbedaan dalam penggunaan habitat oleh spesies ini (Vaudo et al. 2014 ; Lubitz et al. 2022 ).
4.2 Habitat Penting dan Penelitian Hiu Macan di Masa Depan
Habitat esensial adalah area yang mendukung fungsi-fungsi spesifik selama berbagai tahap riwayat hidup dari siklus hidup spesies, seperti mencari makan, berlindung, atau tujuan reproduksi (Barnett et al. 2019 ; Heupel et al. 2019 ; De Wysiecki et al. 2023 ). Informasi tentang habitat esensial yang mendukung populasi hiu macan Australia timur masih terbatas, termasuk lokasi habitat kelahiran. Di sini, bukti penataan genetik lokal, pergerakan terbatas, dan potensi tempat tinggal pada hiu macan betina muda-subdewasa adalah salah satu indikasi paling awal dari potensi filopatri reproduksi dan area persalinan yang disukai pada hiu macan dari Australia timur. Saat ini, informasi tentang filopatri reproduksi, dan perilaku serta penggunaan habitat hiu macan neonatus dan muda-subdewasa (termasuk penyebarannya dari tempat melahirkan), terbatas untuk wilayah ini (Holland et al. 2019 ). Bukti tempat tinggal hiu macan telah dilaporkan di beberapa bagian belahan bumi utara, dan terkait dengan habitat kelahiran, perilaku perlindungan, mencari makan, dan filopatri reproduksi (Driggers et al. 2008 ; Sulikowski et al. 2016 ; Acuña-Marrero et al. 2017 ; McClain et al. 2022 ; Smukall et al. 2022 ). Lebih jauh lagi, telah dikemukakan bahwa beberapa pulau samudra penting untuk siklus reproduksi hiu macan, termasuk Bahama (Sulikowski et al. 2016 ; Smukall et al. 2022 ), Hawaii (Whitney dan Crow 2007 ; Papastamatiou et al. 2013 ), Kepulauan Galapagos (Acuña-Marrero et al. 2017 ), dan Pulau Cocos (Cambra et al. 2021 ), dan mungkin Pulau Norfolk (Matley et al. 2024 ). Temuan kami menunjukkan kemungkinan filopatri reproduksi betina di wilayah pesisir atau landas kontinen Australia timur, dengan habitat kelahiran yang berpotensi mencakup sebagian besar garis pantai dari perairan tropis hingga sedang. Namun, tidak diketahui apakah persalinan kemungkinan besar terjadi di tempat kelahiran/daerah pembibitan tertentu, atau di daerah umum “yang disukai” yang mencakup puluhan km, yang tampaknya paling mungkin mengingat skala autokorelasi spasial yang diamati dalam penelitian saat ini.
Penelitian tambahan diperlukan untuk memvalidasi temuan ini. Misalnya, menyelidiki pergerakan hewan dan penggunaan habitat dengan menggunakan desain pengambilan sampel berstrata yang sama seperti penelitian saat ini. Meskipun Niella et al. ( 2022 ) tidak mendeteksi efek signifikan dari jenis kelamin atau ontogeni pada pola penyebaran hiu macan di wilayah ini, pengambilan sampel yang lebih banyak diperlukan untuk mengonfirmasi temuan ini. Lebih jauh lagi, perluasan pengambilan sampel untuk mencakup neonatus dan juvenil kecil akan membantu mendeteksi tingkat struktur genetik pada tahap kehidupan sebelumnya dan menentukan kemungkinan lokasi habitat kelahiran di wilayah tersebut. Namun, mengakses dan mengambil sampel rentang ukuran ini tetap menantang karena pertemuan jarang terjadi menggunakan peralatan survei yang umum digunakan (Holmes et al. 2014 ; Niella et al. 2022 ; Tate et al. 2021 ). Sebagai alternatif, telah disarankan sebelumnya bahwa pemasangan pemancar intrauterin (Sulikowski dan Hammerschlag 2023) pada betina yang dikonfirmasi melalui USG untuk hamil (Sulikowski et al. 2024 ) dapat membantu mendeteksi peristiwa dan lokasi beranak, yang dapat membantu pengambilan sampel genetik tahap awal kehidupan di masa mendatang.
4.3 Pentingnya Pengambilan Sampel Berstrata
Studi ini menambah literatur yang berkembang yang mendukung pentingnya rezim pengambilan sampel berstrata ketika menyelesaikan pola struktur genetik populasi dan konektivitas pada elasmobranch. Phillips dkk. ( 2021 ) baru-baru ini menantang integritas studi genetik populasi elasmobranch, dengan menyatakan bahwa ketergantungan pada rezim pengambilan sampel oportunistik yang mengarah pada pengumpulan hewan yang bervariasi dalam jenis kelamin dan tahap kehidupan cenderung mengaburkan sinyal genetik penyebaran yang bias jenis kelamin dan filopatri reproduksi. Studi kami mendukung argumen ini, dengan menunjukkan bahwa pengumpulan sampel dapat mengaburkan pola struktur genetik yang terkait dengan jenis kelamin dan tahap kehidupan tertentu. Lebih jauh, kami menunjukkan pentingnya rezim pengambilan sampel yang direplikasi secara spasial yang memungkinkan pengujian langsung autokorelasi spasial dan memberikan wawasan tentang kekuatan sebenarnya dan batas spasial aliran gen. Akibatnya, ada kemungkinan bahwa kekuatan konektivitas biologis di antara elasmobranch baik di dalam maupun lintas samudra di dunia telah dilebih-lebihkan karena ketergantungan umum studi genetik sebelumnya pada pendekatan pengambilan sampel nonstratifikasi (Tabel 1 ).
Peningkatan desain pengambilan sampel diperlukan untuk studi genetika masa depan pada elasmobranch. Namun, seperti yang dibahas oleh Phillips et al. ( 2021 ), hal ini sering kali memerlukan penelitian yang sangat kolaboratif dan sumber daya yang signifikan. Upaya pengambilan sampel untuk studi saat ini dimungkinkan oleh beberapa program penelitian yang diarahkan untuk mengurangi risiko interaksi manusia-hiu di Australia timur dan kawasan Indo-Pasifik. Namun, jenis pengambilan sampel ini memerlukan tingkat investasi yang tinggi dan mungkin tidak memungkinkan di bagian lain dunia karena kendala logistik dan keuangan, atau dalam sistem studi dengan kelimpahan populasi yang rendah (misalnya, spesies langka dan terancam). Akibatnya, peneliti dan manajer harus berhati-hati ketika menafsirkan pola struktur genetik populasi dan konektivitas pada elasmobranch tanpa adanya rezim pengambilan sampel yang bertingkat, karena hal ini dapat berisiko menyesatkan proses pengambilan keputusan konservasi dan pengelolaan.