
Abstrak
- Taktik reproduksi alternatif (ART) menggambarkan strategi alternatif yang tidak dapat dibalikkan atau fleksibel yang mengamankan pembuahan. Misalnya, beberapa jantan mempertahankan wilayah dengan betina sementara yang lain mencoba perkawinan diam-diam. Seringkali, ART dianggap bergantung pada status dan dijelaskan oleh perbedaan dalam massa atau kemampuan bersaing. Namun, sebagian besar penelitian tentang ART hanya memperkirakan efek kebugarannya, mengabaikan jantan yang tidak pernah bereproduksi dan menganggap status (misalnya berat) sebagai satu-satunya mediator ART.
- Kami menggunakan 244 tikus jantan, Mus musculus domesticus , dari populasi semi-alami, untuk mendeskripsikan ART pada Mus Musculus untuk pertama kalinya. Kami mengikuti tikus jantan sepanjang hidup mereka dan mengkategorikan mereka sebagai hewan teritorial atau pengembara selama beberapa interval bulanan, setelah memvalidasi metode kami dalam menetapkan taktik dengan data spasial terperinci.
- Kami menyelidiki apakah pilihan taktik dapat diulang, apakah berbagai faktor sosial dan/atau intrinsik memprediksi pilihan taktik dan transisi antartaktik, dan menguji kebugaran serta perbedaan fisiologis antar ART.
- Pilihan taktik dapat diulang, tetapi jantan dapat berganti taktik secara fleksibel. Pilihan taktik dikaitkan dengan massa, usia, rasio jenis kelamin operasional, dan ukuran populasi. Teritorial memiliki kemungkinan reproduksi yang lebih tinggi, tetapi indeks gonadosomatik yang lebih rendah.
- Hasil kami mengungkap komponen kepribadian ART, mengonfirmasi kebugaran rata-rata yang sama di antara taktik dan menyarankan pilihan taktik sebagai keputusan multifaset di bawah berbagai tekanan selektif.
1. PENDAHULUAN
Pada banyak spesies, jantan menggunakan taktik reproduksi alternatif (yaitu sifat perilaku/morfologi/fisiologi yang berbeda; ART) untuk memaksimalkan kebugaran mereka (Brockmann, 2001 ; Brockmann & Taborsky, 2008 ; Dougherty et al., 2022 ; Gross, 1996 ; Kustra & Alonzo, 2020 ; Schradin & Lindholm, 2011 ; Stockley et al., 1994 ; Taborsky et al., 2008 ). Misalnya, pada chamois Alpen, Rupicapra rupicapra , beberapa jantan mempertahankan area dari penyusup (yaitu pemegang wilayah), sementara yang lain (nonteritorial) mencoba memasuki wilayah jantan lain tanpa izin (Corlatti et al., 2012 ; Taborsky et al., 2008 ). ART dapat bersifat non-reversibel, dikodekan secara genetik atau fleksibel (diulas dalam Gross, 1996 ), dan berevolusi jika ada manfaat kebugaran yang terkait dengan mengejar alternatif (Taborsky et al., 2008 ). Misalnya, tikus belang jantan ( Rhabdomys pumilio ) dapat beralih dari teritorial menjadi pengembara (Schradin & Lindholm, 2011 ). ART yang fleksibel secara fenotip juga dideskripsikan pada hewan pengerat lain, seperti tikus padang rumput ( Microtus ochrogaster , Shuster et al., 2019 ) atau tupai tanah ( Urocitellus columbianus , Balmer et al., 2019 ).
Secara umum, ART lebih sering terdeteksi pada jantan karena variabilitas yang lebih besar dalam keberhasilan reproduksi jantan (Taborsky et al., 2008 ) dan investasi jantan yang lebih tinggi dalam sifat reproduksi yang dapat dieksploitasi oleh spesies sejenis yang mengejar taktik alternatif (Clutton-Brock, 2007 , 2009 ; Queller, 1997 ; Taborsky et al., 2008 ; Taborsky & Brockmann, 2010 ). Taktik yang fleksibel (umum pada ikan, Karkarey et al., 2017 ; Taborsky & Brockmann, 2010 ) dapat berevolusi karena ketidakstabilan dalam ketersediaan pasangan spasial dan temporal (Brockmann & Taborsky, 2008 ; Wolff, 2008 ), karena risiko persaingan atau predasi. Misalnya, kepadatan betina yang tinggi atau rasio jenis kelamin operasional (OSR) yang condong ke jantan lebih mendukung pengembaraan di antara kelompok betina, dan kepadatan populasi yang rendah atau OSR yang condong ke betina lebih mendukung monopoli betina (Wolff, 2008 ), seperti yang terlihat pada lembu kesturi jantan Ovibos moschatus (Forchhammer & Boomsma, 1998 ; Wolff, 2008 ).
Ekspresi diferensial ART jantan dianggap bergantung pada kondisi atau status (Brockmann, 2001 ; Gross, 1996 ; Johnson & Brockmann, 2012 ; Schradin & Lindholm, 2011 ; Shuster & Wade, 2003 ) atau bergantung pada aktivitas betina (Buzatto & Machado, 2008 ; Schradin & Lindholm, 2011 ; Stockley et al., 1994 ; Taborsky et al., 2008 ), ukuran populasi (Radwan, 1993 ; Radwan et al., 2014 ) dan kepadatan (Karkarey et al., 2017 ; Wolff, 2008 ) atau ekologi lokal (misalnya Monroe et al., 2016 ). Secara umum, kemampuan bersaing individu sering kali merupakan proksi yang baik dari pilihan taktik untuk ketergantungan kondisi dan/atau status (Stockley et al., 1994 ), dari artropoda (Brockmann, 2001 ; Hunt & Simmons, 2001 ; Perdigón Ferreira & Lüpold, 2022 ) hingga ikan (Taborsky, 2008 ) dan primata (Setchell, 2008 ). Secara keseluruhan, ART terdiri dari strategi alokasi diskret di antara spesies yang sama: beberapa individu (biasanya jantan) secara langsung mengakses heteroseksual (misalnya sifat seksual sekunder atau pertahanan harem) dan yang lain mengeksploitasi upaya spesies yang sama dan mendapatkan akses langsung ke gamet (taktik ‘borjuis’ dan ‘parasit’; Taborsky, 1997 ), yaitu taktik alternatif ‘yang terbaik dari pekerjaan yang buruk’ (Dawkins, 1980 ) dari perkawinan oportunistik.
Dalam studi ini, kami menyelidiki apakah tikus rumah jantan ( Mus musculus domesticus ) mengikuti ART dan apakah beberapa faktor sosial dan intrinsik dapat memengaruhi ekspresi dan transisi antara taktik pada tingkat individu. Tikus rumah menyediakan model yang sangat baik untuk mempelajari ART dalam konteks kompetisi reproduksi. Pertama, taktik pengembangbiakan alternatif yang bergantung pada kondisi dan kepadatan telah ditetapkan pada tikus betina, yang menggunakan kombinasi pengembangbiakan soliter dan komunal untuk membesarkan anak-anak (Ferrari et al., 2019 , 2022 ). Namun, ART jantan belum pernah dideskripsikan sebelumnya pada tikus jantan meskipun ada interaksi agresif jantan dewasa untuk perolehan teritorial (Gray & Hurst, 1997 ), akses ke sumber daya, peluang kawin atau perlindungan dari predator dan kondisi lingkungan (Kaufmann, 1983 ; Ord, 2021 ). Akibatnya, perilaku ini dapat menyebabkan spesies yang sama berkeliaran, mungkin mencoba perkawinan diam-diam. Selain itu, rasio ayah ganda meningkat hingga ~40% pada tikus (Schradin et al., 2010 )—rata-rata, 21% dari anak tikus merupakan keturunan dari banyak anak (Porwal et al., 2023 )—angka yang relatif tinggi jika dibandingkan dengan hewan pengerat lainnya (diulas dalam Solomon & Keane, 2008 ) dan mamalia lainnya secara umum (diulas dalam Cohas & Allainé, 2009 ). Ini berarti bahwa tikus teritorial kehilangan ayah dan menyiratkan bahwa tikus jantan yang mengikuti strategi reproduksi alternatif (selain perolehan teritorial) mungkin ada dan mencuri ayah.
Di sini, kami mengukur taktik jantan (pengembara & teritorial) di dua generasi tikus rumah jantan yang hidup semi-alami dalam empat populasi yang direplikasi dan hingga 11 bulan per individu ( N = 244). Kami juga mengukur pertumbuhan dan reproduksi jantan ini dalam kehidupan dewasanya. Kami kemudian membandingkan dua metode untuk menilai ART jantan: (1) kami mengukur di mana jantan ditangkap per bulan (di dalam kotak sarang—disebut teritorial selanjutnya—atau berkeliaran) dan (2) membandingkan data tersebut dengan data antena RFID yang lebih rinci (dari dua populasi independen) yang mengukur lokasi jantan selama 2 bulan terus menerus (König et al., 2015 ). Dengan menggunakan data ini kami memeriksa apakah taktik jantan dapat diulang, sesuatu yang sepengetahuan kami belum pernah diuji sebelumnya pada mamalia dalam konteks ART dengan juga memantau semua jantan (untuk studi jangka panjang serupa yang mengukur ART, lihat Balmer et al., 2019 ; Fasel et al., 2016 ). Yang penting, kami mengharapkan jantan menunjukkan taktik plastik karena kondisi lingkungan dan sosial bervariasi terus menerus, secara spasial dan temporal, dalam populasi tikus rumah (DeLong, 1967 ; Singleton et al., 2005 ; Wilson et al., 2018 ). Selanjutnya, kami mengeksplorasi apakah ART menghasilkan kebugaran yang berbeda (Brockmann & Taborsky, 2008 ; Gross, 1996 ) dan mengukur aspek lingkungan sosial jantan (OSR dan ukuran populasi), serta faktor intrinsik seperti massa atau usia, untuk menguji korelasinya dengan pilihan ART. Kami memperkirakan pilihan taktik bergantung pada massa individu karena jantan yang lebih berat seharusnya lebih kuat dan memiliki kemampuan bersaing yang meningkat (Schradin, 2019 ), sehingga bersifat teritorial. Kemudian, kami memperkirakan pilihan taktik bergantung pada usia, dengan kemungkinan teritorialitas meningkat ketika jantan tumbuh kuat tetapi kemudian secara bertahap menurun karena kemampuan yang menurun untuk mengalahkan spesies yang sama saat jantan menjadi lebih tua; dan pada jumlah betina reseptif, yang seharusnya mendorong apakah jantan bertahan atau berkeliaran untuk mendapatkan kesempatan kawin. Terakhir, kami memperkirakan ART berkorelasi dengan ukuran populasi karena peningkatan populasi pasti akan menghasilkan penipisan kotak sarang yang tersedia (atau lebih umum, teritorial; dijelaskan di Bagian 2.1) dengan lebih cepat.), dan karenanya harus memprediksi probabilitas yang lebih tinggi untuk berkeliaran. Kami juga memodelkan probabilitas untuk beralih antara teritorialitas ke jelajah dan sebaliknya, serta rasio bahaya dari setiap taktik, menggunakan model multi-negara. Di sini, kami menetapkan bahwa beralih dari teritorialitas ke jelajah, teritorialitas ke kematian dan jelajah ke kematian bergantung pada intensitas persaingan intraseksual, wilayah yang tersedia, dan usia individu: persaingan yang lebih tinggi (peningkatan OSR atau ukuran populasi) harus mengarah pada tingkat cedera atau kematian yang lebih tinggi, dan pria yang lebih tua harus memiliki kemampuan bersaing yang menurun dan dengan demikian lebih rentan terhadap pemindahan dari wilayah atau mati (dari salah satu taktik). Selain itu, kami memperkirakan usia, tingkat persaingan pria-pria dan ukuran populasi memengaruhi transisi dari jelajah ke teritorialitas jika pengembara muda yang tumbuh lebih tua menjadi lebih kompetitif dan jika lebih sedikit individu dan pesaing (pria lain) dalam populasi meningkatkan peluang untuk perolehan teritorial. Terakhir, kami meneliti apakah ART yang berbeda terkait dengan perbedaan fisiologis seperti jumlah sperma, panjang tubuh, dan rasio testis terhadap massa tubuh karena teori memprediksi bahwa pria yang berkeliaran mengalami peningkatan risiko persaingan sperma dan dengan demikian lebih banyak berinvestasi pada sifat sperma (Dougherty et al., 2022 ; Kustra & Alonzo, 2020 ).
2 METODE
2.1 Hewan dan perumahan
Tikus berasal dari populasi liar yang diambil sampelnya di wilayah Cologne/Bonn di Jerman ( N = 18 pasangan pembiakan asli; 50°45′ N–51° N, 6°45′ E–7° E). Tikus pendiri ( N = 160, 80 jantan, 80 betina) untuk populasi semi-alami didistribusikan ke empat kandang replikasi (masing-masing 19,6 m 2 ), dengan 20 jantan dan 20 betina sebagai pendiri setiap populasi.
Bagi tikus rumah, lingkungan alami adalah lumbung atau tempat berlindung manusia (König & Lindholm, 2012 ) dan, untuk meniru lingkungan seperti itu, setiap kandang semi-alami dilengkapi dengan berbagai bahan bersarang dan 12 kotak sarang. Kotak sarang ini berfungsi sebagai lokasi utama bagi keluarga tikus untuk hidup, berkembang biak, dan membesarkan keturunan dan merupakan inti dari wilayah jelajah yang secara aktif dipertahankan oleh tikus jantan dewasa (yaitu pemegang wilayah) (Gambar S1 ). Sementara wilayah jelajah biasanya lebih besar (Gambar S1 ), kotak sarang secara aktif dipertahankan terhadap penyusup. Makanan dan air disediakan sepuasnya dan didistribusikan secara merata di seluruh kandang di sembilan tempat makan. Kondisi seperti itu mirip dengan yang ditemukan pada populasi tikus rumah alami Eropa (Anderson, 1982 ).
Kami memantau perkembangan populasi setiap 4-5 minggu. Perkembangan populasi dan kepadatan di kandang semi-alami meniru populasi tikus yang berada dalam pengamatan jangka panjang dengan kemungkinan penyebaran (König & Lindholm, 2012 ). Selama pemantauan ini, kami menangkap semua tikus di dalam kandang, mengukur massa tubuh, memeriksa tanda gigitan baru dan mengambil sampel jaringan individu baru (berat >10 g) untuk penugasan induk. Hewan baru (>10 g) menerima tag RFID PIT (Planet ID, 1,4 × 9 mm) untuk pengenalan permanen. Setiap kali populasi tertentu mencapai setidaknya 80 keturunan yang diberi chip, kami membuang generasi yang lebih tua (biasanya setelah 8-10 bulan) untuk menjaga kepadatan di bawah daya dukung. Karena alasan ini, pengamatan kami dibatasi hingga usia maksimum 11 bulan. Beberapa hewan yang dibuang tersebut di-eutanasia untuk mendapatkan karakteristik fisiologis, misalnya melakukan penghitungan sperma.
2.2 Penugasan taktik reproduksi
Selama pemantauan populasi bulanan rutin kami, setiap jantan dewasa diberi taktik berdasarkan lokasinya (visualisasi pada Gambar S2 ). Ketika memasuki kandang, beberapa tikus (semuanya dapat diidentifikasi melalui chip mereka) berada di dalam kotak sarang atau memasuki satu kotak sarang, sementara yang lain tetap di luar karena mereka tidak mencoba memasuki kotak sarang atau karena mereka segera diusir ketika mencoba melakukannya. Kotak sarang kemudian ditutup (yaitu identitas hewan di dalam kotak sarang diketahui) dan semua hewan (di dalam kotak sarang dan semua hewan di luar) yang berada di kandang dikeluarkan, diidentifikasi, dan ditimbang (hingga desimal terdekat, misalnya 28,4 g). Karena kami dapat mengidentifikasi semua tikus secara individual, kami dapat menyimpulkan kelangsungan hidup berdasarkan ada/tidaknya hewan; dengan demikian, kami juga memiliki perkiraan bulanan untuk kepadatan populasi dan karakteristik individu seperti kelangsungan hidup.
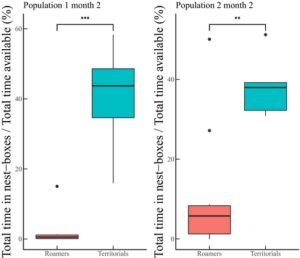
Bahasa Indonesia: Jika seekor jantan dewasa berada di dalam kotak sarang sebagai satu-satunya jantan dewasa di sana, ia diidentifikasi sebagai pemegang teritori, dan jika di luar kotak sarang, sebagai pengembara. Kami mengecualikan pengamatan dari bulan-bulan ketika lebih dari satu jantan dewasa (>4 minggu) ditemukan di kotak sarang tertentu karena tidak jelas jantan mana yang memonopoli kotak sarang (tetapi data ini mencakup sebagian kecil dari ukuran sampel awal: ~10%; N = 199/1699; lihat Gambar 1 ). Untuk memvalidasi bahwa ‘gambaran’ lokasi jantan selama pemantauan benar-benar mencerminkan perbedaan yang berarti dalam perilaku jantan sepanjang waktu, kami menjalankan eksperimen tambahan dengan 107 jantan selama 2 bulan dalam dua populasi tambahan (populasi ini hanya digunakan untuk memvalidasi penugasan taktik kami dan jantan bukan bagian dari 244 individu yang digunakan dalam semua analisis lainnya; lihat Gambar 1 ). Selama percobaan validasi ini, kami memasang antena RFID di pintu masuk semua kotak sarang untuk merekam lokasi dan durasi tinggal di dalam kotak sarang. Kami menghitung jumlah waktu yang dihabiskan setiap jantan dewasa di dalam kotak sarang dan mengkorelasikannya dengan penugasan taktik selama pemantauan (Gambar 2 ).
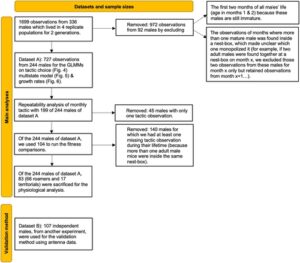
2.3 Kumpulan Data
Kami menggunakan dua kumpulan data independen, seperti yang ditampilkan di bawah ini (Gambar 1 ): satu untuk memvalidasi penugasan taktik kami sebagai metode yang bermakna secara biologis untuk menetapkan taktik; dan satu lagi untuk semua analisis ART lainnya.
2.4 Kebugaran
Kami menggunakan 17 penanda mikrosatelit untuk menentukan garis keturunan dan menetapkan kebugaran menggunakan prosedur yang diadaptasi dari Linnenbrink et al. ( 2013 ). DNA diekstraksi dari klip telinga, diperkuat menggunakan kit PCR Multiplex (QIAGEN) dan sampel dijalankan pada ABI 3730 Sequencer (Applied Biosystems). Kami menggunakan GeneMarker (V2.6.4) untuk mengidentifikasi alel dan Colony [©COLONY Zoological Society of London (ZSL)] untuk menetapkan garis keturunan berdasarkan kemungkinan maksimum setiap pasangan parental potensial. Kami mengkondisikan penugasan garis keturunan kami pada asumsi berikut: reproduksi seksual, sistem perkawinan poligami, kemungkinan perkawinan sedarah, dan semua hewan hadir bulan sebelum pengambilan sampel (yaitu ketika juvenil lahir) sebagai kemungkinan parental. Secara keseluruhan, estimasi kebugaran kami mengukur jumlah keturunan yang bertahan hidup hingga setidaknya 10 g (Gambar S4 ).
Bahasa Indonesia: Untuk mengeksplorasi perbedaan antara ART dalam keberhasilan reproduksi, kami mengukur kebugaran dalam empat cara berbeda untuk 104 jantan (50 yang bereproduksi dan 54 yang tidak pernah bereproduksi). Kami fokus pada 104 (dari 244) jantan yang dapat kami tetapkan pilihan taktiknya setiap bulan tanpa ambiguitas dan yang kami miliki semua observasi taktik bulanannya sepanjang hidup mereka. Dengan kata lain, kami mengecualikan 140 jantan yang setidaknya satu penugasan taktiknya tidak pasti (dijelaskan dalam Gambar 1 ). Dengan melakukan itu, kami mengurangi gangguan untuk prediktor kebugaran kami [rumus (1)]. Kami mengukur kebugaran menggunakan: (1) variabel biner (0/1) yang menggambarkan reproduksi yang berhasil atau tidak; (2) jumlah total keturunan setiap jantan (kebugaran absolut); (3) jumlah keturunan yang dihasilkan setiap jantan selama hidupnya dibagi dengan jumlah rata-rata keturunan semua jantan di kandang semi-alami itu (kebugaran relatif); (4) jumlah total keturunan setiap jantan dibagi dengan rentang hidupnya (didefinisikan sebagai kebugaran/usia di bagian Pemodelan ). Metrik 3 mengidentifikasi kebugaran relatif setiap jantan berdasarkan taktik yang diikuti setiap jantan sepanjang hidupnya. Metrik 4 ‘menetapkan stratifikasi’ individu berdasarkan hasil keturunan dan kelangsungan hidup mereka. Kami menggunakan individu yang bereproduksi dengan sukses, bersama dengan yang tidak, karena mengabaikan yang terakhir pasti akan meningkatkan estimasi kebugaran rata-rata serta mengurangi varians kebugaran untuk setiap taktik (Shuster, 2011 ).
Kami menggunakan rumus berikut sebagai prediktor (dalam bagian efek tetap dari rumus) dalam semua analisis, yang memiliki kebugaran sebagai respons:
![]()
di mana R berarti pengembara dan T teritorial. Metrik ini, yang mengukur rasio berapa kali setiap jantan tertangkap sebagai pengembara dengan total berapa kali ia tertangkap (karena seekor jantan dapat ditangkap sebagai teritorial atau pengembara), memungkinkan kami untuk memperhitungkan keputusan taktik seumur hidup masing-masing individu.
2.5 Korelasi dan tingkat transisi ART
Dengan mempertimbangkan korelasi antara ART dengan aspek intrinsik atau lingkungan, serta probabilitas transisi antara taktik, kami pertama-tama menggunakan OSR, sebagai rasio jantan dewasa terhadap betina yang aktif secara seksual. Kemudian, kami memasukkan dalam analisis usia dan massa sebagai proksi kondisi dan status, serta ukuran populasi untuk memahami apakah ART bergantung pada kepadatan. Terakhir, kami mengukur 83 jantan yang ditangkap secara acak (dari 244; 66 diidentifikasi sebagai pengembara dan 17 sebagai teritorial bulan sebelum analisis ini), panjang tubuh, berat testis dan persentase massa tubuh testis yang ditempati (yaitu indeks gonadosomatik, GSI, lihat Informasi Pendukung; Tomkins & Simmons, 2002 ), karena perbedaan dalam aspek fisiologis ini akan menunjukkan strategi alokasi yang berbeda terhadap sifat reproduksi yang mahal. Kami menghitung sperma menggunakan versi modifikasi dari prosedur yang dijelaskan oleh Wang ( 2002 ). Secara khusus, epididimis diiris dalam larutan penyangga fosfat, suspensi sperma diinkubasi dalam thermomixer selama 10 menit pada suhu 40°C untuk membunuh sperma, dan 10 mL suspensi sperma yang diencerkan dengan perbandingan 1:20 atau 1:40 digunakan untuk mendapatkan jumlah sperma sekitar 3–10 per kotak dalam ruang Burker. Kemudian, jumlah sperma dihitung dua kali di bawah mikroskop pada perbesaran 40x dengan filter PH2 dalam 25 kotak.
2.6 Data antena dan analisis statistik
Pada populasi independen kami, antena RFID beresolusi tinggi dipasang di pintu masuk setiap sarang (AniLoc, FBI Systems GmbH, Jerman), dan secara otomatis merekam waktu yang dihabiskan setiap tikus di setiap sarang (König et al., 2015 ). Data dikumpulkan menggunakan program OLCUS IDE (FBI Systems GmbH, Jerman).
Semua analisis, termasuk analisis data antena, dilakukan menggunakan R versi 4.2.2.
2.7 Pengulangan dan pemodelan
Untuk menentukan seberapa banyak variasi dalam pilihan jantan untuk menjadi teritorial atau pengembara disebabkan oleh perbedaan antara individu, kami menghitung pengulangan pilihan taktik bulanan menggunakan paket R rptR (Stoffel et al., 2017 ). Estimasi yang relatif tinggi (estimasi rata-rata untuk perilaku adalah ~37%, Bell et al., 2009 ) akan menunjukkan bahwa beberapa jantan secara konsisten lebih cenderung menjadi teritorial/pengembara dibandingkan dengan spesies yang sama. Jantan dengan hanya satu observasi dikecualikan (kami berakhir dengan ukuran sampel N = 199) dan kami mengasumsikan struktur kesalahan binomial. Kami memasukkan identitas jantan sebagai efek acak dan usia sebagai efek tetap untuk mengecualikan kontribusinya pada estimasi kami. Permutasi dan bootstrapping ditetapkan pada 1000, dan interval kepercayaan pada 95%.
Untuk mengkorelasikan ART dengan keberhasilan reproduksi, kami meregresikan empat metrik kebugaran yang dijelaskan dalam 2.4 (dalam empat model) dengan rasio waktu setiap jantan ditangkap sebagai pengembara terhadap total waktu ditangkap (rumus a) untuk jantan dengan semua pengamatan bulanan ( N jantan = 104). Pertama-tama kami memodelkan probabilitas reproduksi sebagai respons 0/1 (0 = tidak ada reproduksi; 1 = reproduksi) menggunakan regresi logistik (statistik paket R). Kemudian, kami memodelkan kebugaran (jumlah individu keturunan) menggunakan model binomial negatif dua bagian yang diubah nol (
disertakan prediktor untuk nilai >0 dan untuk bagian binomial; paket pscl, Zeileis et al., 2008 ), karena banyak jantan tidak bereproduksi dan untuk mengoreksi penyebaran berlebih. Terakhir, kebugaran relatif dan kebugaran/usia adalah variabel respons dalam dua model linier umum (statistik paket R) dengan asumsi distribusi gamma terpotong nol. Yang penting, dua metrik terakhir adalah nol-inflasi tetapi kami mengecualikan nol karena kami mempertimbangkannya (nol) untuk probabilitas reproduksi dan kebugaran absolut.
Kami menggunakan model efek campuran umum (glmm; paket R lme4, Bates et al., 2015 ) dengan taktik bulanan sebagai respons binomial (teritorial/pengembara, 727 observasi dari N laki-laki = 244) untuk mengidentifikasi korelasi ART. Untuk memilih prediktor terbaik dari sekumpulan kandidat (prediktor ini dipilih karena mereka secara konsisten dijelaskan untuk memengaruhi ekspresi ART, lihat Pendahuluan), kami menggunakan AICc (Tabel S1 ). Secara khusus, kami mengeksplorasi apakah interaksi massa dengan usia memiliki daya penjelas, apakah OSR harus digunakan atau jumlah perempuan aktif secara seksual dan laki-laki dewasa sebagai variabel individu, jika ukuran populasi dan jumlah subdewasa meningkatkan daya penjelas suatu model (penjelasan terperinci dalam Tabel S1 ). Akhirnya, model kami menyertakan massa dan usia untuk menguji apakah status atau usia berkorelasi dengan taktik, masing-masing, OSR (jantan dewasa/betina yang aktif secara seksual) untuk menguji apakah ketersediaan pasangan memengaruhi ART dan ukuran populasi (berskala, yaitu dikurangi rata-rata dan dibagi dengan simpangan baku) untuk menguji apakah ART bergantung pada kepadatan. Efek acak menyertakan ID individu yang disarangkan dalam ID kandang. Model terbaik berkinerja lebih baik daripada model nol dengan hanya intersep ( p < 0,001).
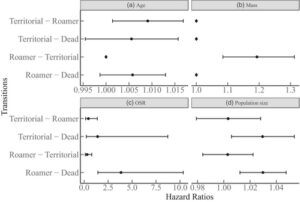
Terakhir, kami menggunakan model Markov multi-state (msm: R package msm, Jackson, 2011 ) untuk mengeksplorasi probabilitas transisi antara ART, dan rasio bahaya masing-masing (Tabel S2 ; Gambar S5 ), sebagai fungsi kovariat untuk 244 laki-laki. Probabilitas transisi untuk setiap taktik dimodelkan dengan interval kepercayaan 95%, dengan memperhitungkan identitas dan usia individu. Kami memperbolehkan laki-laki untuk bertransisi bebas antara taktik untuk mencocokkan data observasional (Gambar S3 ), dan kematian adalah kondisi penyerapan. Kami menggunakan kovariat yang sama seperti di atas dalam glmm untuk memprediksi intensitas transisi (Tabel S2 ). Usia, OSR, dan ukuran populasi memprediksi transisi antara teritorialitas ke jelajah dan antara teritorialitas ke kematian. Kami berasumsi ini karena kami memperkirakan teritorial akan tergusur/mati seiring bertambahnya usia, sebagai indikasi penurunan kemampuan bersaing, dan lebih mungkin tergusur/mati saat kompetisi intraseksual (OSR dan ukuran populasi yang lebih tinggi) meningkat. Usia, OSR, dan ukuran populasi juga dikaitkan dengan transisi antara pengembaraan hingga kematian karena kami memperkirakan pengembara yang lebih tua akan kurang kompetitif, dan lingkungan yang kompetitif akan meningkatkan angka kematian. Terakhir, massa, OSR, dan ukuran populasi memodelkan transisi antara pengembaraan hingga teritorialitas, dengan asumsi pengembara yang bertambah berat, atau mengalami lebih sedikit persaingan, akan mencoba mengklaim kotak sarang.
2.8 Korelasi intrinsik
Untuk membandingkan ciri fisiologis antara ART, kami menggunakan uji-t untuk membandingkan massa antara taktik per bulan, berat testis dan panjang tubuh (keduanya diukur hanya sekali ketika hewan secara acak dikeluarkan dari populasi dan dikorbankan) karena semuanya mengikuti distribusi Gaussian; dan uji Wilcoxon rank-sum untuk rasio testis terhadap massa tubuh (GSI) dan jumlah sperma karena keduanya tidak (sekali lagi keduanya diukur hanya sekali, ketika hewan secara acak dikeluarkan dari populasi dan dikorbankan). Yang penting, untuk GSI, kami menjalankan ANCOVA (Tomkins & Simmons, 2002 ) untuk mengeksplorasi pengaruh taktik bulanan pada berat testis dengan mengendalikan efek massa, sehingga menguji homogenitas lereng regresi antara taktik bulanan dan massa (untuk detailnya lihat Informasi Pendukung).
2.9 Pernyataan etika
Semua hewan ditangani dan prosedur dilakukan sesuai dengan pedoman nasional. Prosedur disetujui berdasarkan lisensi V244-12767/2019 dan V244-31223/2019(62–5/19) oleh Ministerium für Energiewende, Landwirtschaftliche Räume und Umwelt, Kiel. Pemeliharaan tikus telah disetujui dan dikontrol secara teratur oleh Veterinäramt Plön dengan izin: 1401-144/PLÖ-004697.
3 HASIL
Dalam eksperimen validasi kami menggunakan data antena, diferensiasi yang jelas di antara jantan yang sebelumnya kami klasifikasikan sebagai teritorial atau pengembara muncul: teritorial menghabiskan sebagian besar waktu mereka di kotak sarang sementara pengembara berada di luar dan hanya ‘mengunjungi’ kotak sarang untuk sebagian kecil dari total waktu mereka (uji Wilcoxon rank-sum, Gambar 2 : grafik kiri p < 0,001; grafik kanan p < 0,01). Oleh karena itu, data ini mencerminkan penggunaan kotak sarang yang lebih rinci oleh jantan mengikuti taktik yang berbeda. Akibatnya, kami menggunakan lokasi jantan selama setiap pemantauan untuk menggambarkan taktik bulanan karena mencerminkan aktivitas di dalam dan di luar kotak sarang.
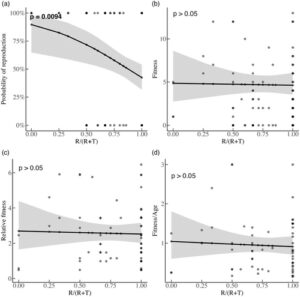
Taktik tersebut cukup dapat diulang ( R = 0,18, CI = 0,067–0,3, p < 0,001). Dari 199 (dari 244 yang tersedia; Gambar 1 ) laki-laki yang diuji untuk pengulangan, 9 selalu teritorial, 61 selalu berkeliaran dan 129 berpindah setidaknya sekali. Dari yang terakhir, 24% (31 laki-laki) berpindah lebih dari sekali antara teritorialitas dan berkeliaran (Gambar S3 ). Distribusi kebugaran di antara jantan yang kami miliki semua pengamatannya (104 dari 244 yang tersedia; Gambar 1 ) sangat miring dengan setengahnya tidak bereproduksi, dan dengan jantan yang bereproduksi memperoleh antara 1 dan 13 keturunan (rata-rata = 2,7 ± 3,6; Gambar S4 ). Kami menemukan bahwa jantan lebih sering berlokasi sebagai pengembara (lebih tinggi nilai) lebih kecil kemungkinannya untuk bereproduksi (logistik & model dua bagian/rintangan: β = −2,4, SE = 0,92, p = 0,009; Gambar 3 ). Namun, tidak memprediksi kebugaran untuk bagian hitungan model rintangan ( β = −0,04, SE = 0,20, p > 0,05), yaitu kebugaran rata-rata tidak berubah berdasarkan pilihan ART seumur hidup. Saat menguji efek
pada kebugaran relatif dan kebugaran/usia, kami sekali lagi tidak menemukan signifikansi (kebugaran relatif GLM: β = −0,06, SE = 0,30, p > 0,05; kebugaran/usia GLM: β = −0,12, SE = 0,33, p > 0,05). Yang penting, roamer menghasilkan keturunan (61/104 jantan selalu roamer dan 22 dari 61 ini berhasil bereproduksi) di beberapa titik selama hidup mereka.
Dengan berfokus pada semua jantan yang tersedia (244), yaitu menambahkan ke 104 di atas yang tidak memiliki pengamatan (140), tidak mengubah hasil ini: dari 244, 19 selalu teritorial, 78 selalu berkeliaran dan 147 mengganti taktik setidaknya sekali. Kebugaran rata-rata tidak berbeda secara signifikan antara kategori karena teritorial memiliki 2,47 ± 2,99 (rata-rata & galat standar) keturunan, pengembara 1,91 ± 3,16 dan pengalih 2,45 ± 3,64.
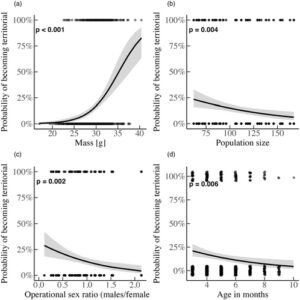
Peluang teritorialitas meningkat hampir 35% (glmm: 95% CI = [1.22, 1.50], p < 0.001; Gambar 4a ; Tabel S3 ) dengan peningkatan massa, dan satu unit peningkatan usia (yaitu 1 bulan) meningkatkan kemungkinan roaming sebesar 22% (95% CI = [0.65, 0.93], p = 0.006; Gambar 4d ). Peningkatan ukuran populasi dikaitkan dengan penurunan 36% dalam kemungkinan teritorialitas (95% CI = [0.47, 0.87], p = 0.004; Gambar 4b ). Terakhir, OSR yang cenderung didominasi laki-laki berkorelasi dengan probabilitas roaming yang 67% lebih tinggi (95% CI = [0,17, 0,67], p = 0,002; Gambar 4c ; OSR: 0,11–2,13).
Hasil serupa diperoleh dari analisis multi-negara (Gambar S5 ). Rata-rata, total lama tinggal yang diharapkan di setiap negara adalah ~55 hari sebagai teritorial dan ~150 sebagai pengembara, sementara teritorial tetap demikian selama ~20 hari berturut-turut dan pengembara sebagai pengembara selama ~65 hari. ART juga berbeda dalam tingkat kelangsungan hidup dasar (dengan semua kovariat pada rata-ratanya; Tabel S2 ). Secara khusus, teritorial memiliki probabilitas kematian yang lebih tinggi setelah 30, 60 dan 180 hari dibandingkan dengan pengembara (lihat Tabel S2 ). Kovariat juga memengaruhi taktik secara berbeda. OSR yang lebih tinggi berkorelasi dengan risiko kematian hampir empat kali lipat untuk pengembara (msm: 95% CI = 1,42–10,33) dan dengan peningkatan risiko kematian untuk teritorial (~40% lebih tinggi, 95% CI = 0,22–8,7; Tabel S2 ; Gambar 5c ). Selain itu, untuk setiap kenaikan berat satu gram, seorang pengembara memiliki kemungkinan ~20% (95% CI = 1,08–1,31; Gambar 5b ) lebih besar untuk menjadi teritorial. Sebaliknya, usia dan ukuran populasi tidak secara signifikan (secara statistik atau biologis; Gambar 5a,d ) memengaruhi tingkat transisi antarnegara.
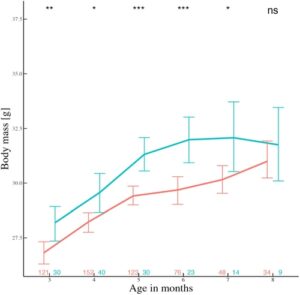
Tikus teritorial lebih berat daripada tikus roamer kecuali pada usia 8 bulan (Gambar 6 , namun perhatikan penurunan ukuran sampel) dan memiliki massa rata-rata keseluruhan yang lebih tinggi [ t (226) = 5,7, p < 0,001;= 30,5 &= 28,9]. GSI berbeda antara ART ( W = 332,5, p = 0,01; Gambar S6 ) dan ini benar bahkan ketika menggunakan ANCOVA dengan berat testis sebagai respons, massa sebagai kovariat dan taktik bulanan sebagai prediktor. Sebaliknya, jumlah sperma tidak berbeda antara ART ( W = 417, p = 0,1). Ada juga korelasi positif signifikan secara keseluruhan antara GSI dan jumlah sperma (rho = 0,32, p = 0,003; Gambar S7 ). Sebaliknya, analisis kami tidak menunjukkan perbedaan antara ART jantan dalam panjang tubuh atau massa testis (Tabel S4 ).
4 DISKUSI
Di sini, pertama-tama kami menemukan bahwa pilihan taktik jantan dapat diulang meskipun faktanya jantan sering berganti taktik. Penerapan taktik juga berkorelasi dengan faktor sosial dan individu, yang juga memengaruhi tingkat transisi di antara keduanya. Terakhir, tikus teritorial lebih mungkin bereproduksi tetapi memiliki nilai GSI yang lebih rendah.
Dalam semua pemantauan, beberapa kotak sarang tidak berpenghuni (observasi pribadi & data antena). Akibatnya, baik jantan berpatroli di lebih dari satu kotak sarang atau beberapa tetap berkeliaran meskipun ada potensi untuk memperoleh kotak sarang. Yang terakhir ini mungkin tampak kontradiktif, karena tampaknya bermanfaat bagi jantan untuk memperoleh kotak sarang tempat betina berkumpul. Mungkin di sini, seperti pada banyak spesies, teritorialitas memerlukan kemampuan bersaing yang tinggi atau pengeluaran energi untuk mencegah spesies yang sama (Ord, 2021 ) dan dapat meningkatkan stres (Hunninck et al., 2020 ), risiko kematian dan cedera. Memang, kadar testosteron berkorelasi positif dengan tuntutan energi (Schradin et al., 2009 ) yang pada gilirannya terkait dengan mempertahankan wilayah (Ord, 2021 ). Atau, jika kualitas pengembara tidak memadai untuk menarik betina maka taktik parasit dapat memberikan manfaat kebugaran bagi jantan tersebut (Luttbeg, 2004 ).
4.1 Pengulangan
Kami menguji apakah pilihan taktik dapat diulang meskipun fakta bahwa pengulangan perilaku kawin jantan kurang mendapat perhatian (Hoefler et al., 2009 ; tetapi lihat Magellan & Magurran, 2007 ; Rushbrook et al., 2008 ). Hasil kami menunjukkan pengulangan sedang (Bell et al., 2009 ), karena ~20% variasi antara jantan dalam pilihan taktik mereka dijelaskan oleh perbedaan antar-individu yang konsisten. Akibatnya, kami melaporkan ART pada tikus rumah memiliki komponen kepribadian; tetapi mereka juga fleksibel karena individu beralih (Gambar S3 ), sesuatu yang dilaporkan di antara hewan pengerat dan mamalia lain (Schradin et al., 2009 ; Schradin & Lindholm, 2011 ; Schradin & Yuen, 2011 ; Shuster et al., 2019 ; Wolff, 2008 ). Dengan kata lain, seiring waktu, beberapa jantan cenderung lebih sering berganti taktik daripada yang lain yang jarang berganti taktik atau tidak sama sekali. Temuan kami bertolak belakang dengan Schradin et al. ( 2009 ), yang melaporkan bahwa tikus belang teritorial jarang berganti taktik, tetapi mereka setuju dengan studi capung jarum ( Calopteryx maculate ; Forsyth & Montgomerie, 1987 ), dan dengan pengamatan lain pada mamalia (diulas dalam Wolff, 2008 ) atau ikan (diulas dalam Taborsky, 2008 ), di mana tikus teritorial dapat beralih ke taktik berkeliaran/menyelinap.
4.2 Kebugaran ART
Kami menunjukkan perbedaan kebugaran antara ART terletak pada reproduksi yang berhasil daripada kebugaran relatif atau absolut. Semakin sering jantan menjadi pengembara, semakin rendah kemungkinannya untuk bereproduksi. Meskipun demikian, karena beberapa pengembara bereproduksi, estimasi kebugaran rata-rata serupa antara jantan yang mengikuti taktik yang berbeda. Secara komparatif, tikus jantan penghuni dan pengembara (Shuster et al., 2019 ) memiliki kebugaran yang sama, dan ART tidak memiliki efek kebugaran pada kepadatan populasi menengah pada tikus belang (Schradin & Lindholm, 2011 ). Oleh karena itu, hasil kami menunjukkan bahwa tidak semua jantan melakukan perilaku paling sukses secara bersamaan (Brockmann, 2002 ). Pada akhirnya, seleksi yang bergantung pada frekuensi dapat menciptakan pola yang mirip dengan pola kita melalui fluktuasi karena musim kawin atau ukuran populasi (Schradin & Lindholm, 2011 ), seperti yang ditunjukkan pada spesies lain (misalnya seleksi yang bergantung pada frekuensi negatif pada ikan Xiphophorus multilineatus , Rios-Cardenas et al., 2018 ).
Persaingan interseksual yang intens mendahului evolusi ART (Shuster & Wade, 2003 ) dan di sini setengah dari jantan yang kami amati tidak pernah bereproduksi (Gambar S4 ). Sementara efek kebugaran ART dalam populasi kami terletak pada kemungkinan reproduksi yang berhasil, varians dalam keberhasilan reproduksi lebih tinggi untuk jantan yang paling sering mengadopsi taktik roaming, tetapi rata-ratanya sama (Gambar 3 ). Dengan kata lain, memasukkan jantan yang tidak pernah bereproduksi dapat menggeser konsekuensi kebugaran ART (Brockmann, 2008 ; Shuster, 2011 ; Shuster et al., 2019 ). Kebugaran rata-rata yang sama dapat dicapai karena di dalam kandang (Porwal et al., 2023 ), serta dalam populasi alami, paternitas ekstra-pasangan tinggi (Schradin et al., 2010 ; Schradin & Lindholm, 2011 ) dan pengembara berkontribusi terhadapnya (Schradin et al., 2010 ). Sementara pendorong paternitas ekstra-pasangan masih belum diketahui (Schradin & Lindholm, 2011 ), itu bisa jadi pilihan betina aktif untuk pengembara (Lehnert et al., 2017 ), paksaan dari pengembara atau keduanya. Atau, tidak adanya pemangsaan dan imigrasi di kandang dapat memengaruhi kebugaran di seluruh taktik, terutama karena risiko pemangsaan harus memengaruhi taktik mana yang dipilih jantan (misalnya pada jangkrik Torsekar & Balakrishnan, 2020 ).
4.3 Korelasi dan tingkat transisi ART
ART jantan sering bergantung pada kondisi atau status (Brockmann, 2001 ; Gross, 1996 ; Johnson & Brockmann, 2012 ; Shuster & Wade, 2003 ), diekspresikan secara berbeda di bawah ukuran populasi yang bervariasi (Radwan, 1993 ; Radwan et al., 2014 ) dan dipengaruhi oleh ketersediaan betina dan OSR (Eberle & Kappeler, 2004 ; Wolff, 2008 ). Di sini, berat yang lebih tinggi meningkatkan kemungkinan jantan menjadi teritorial (Gambar 4a ), meramalkan transisi antara roaming ke teritorialitas (Gambar 5 ), dan teritorial secara konsisten lebih berat daripada roamer (Gambar 6 ). Oleh karena itu, jantan yang lebih berat, seperti pada spesies hewan pengerat lainnya (Schradin et al., 2009 ), mengendalikan sumber daya penting seperti kotak sarang dalam kasus kami. Berat badan, yang mungkin mencerminkan kemampuan, kondisi, dan kekuatan kompetitif, menjelaskan variasi dalam pilihan taktik dan transisi taktik. Temuan kami mengisyaratkan bahwa seleksi yang bergantung pada status berlaku dalam populasi kami meskipun faktanya pilihan taktik tidak memprediksi kebugaran relatif (Schradin & Lindholm, 2011 ). Yang penting, perbedaan massa yang diamati antara ART diminimalkan dari bulan ke-8 (dan seterusnya), mungkin karena jumlah teritorial yang rendah.
Jika perolehan teritorial dikaitkan dengan peningkatan kompetisi (untuk mempertahankan betina atau anak, Schradin et al., 2009 ), maka jantan yang lebih tua mungkin lebih buruk dalam menghalangi sesama spesies (Rathke et al., 2017 ) dan mempertahankan wilayah mereka (Forsyth & Montgomerie, 1987 ). Namun, peningkatan usia tidak memprediksi transisi langsung ke jelajah/kematian, tetapi dikaitkan dengan peningkatan peluang jelajah (Gambar 5a ). Dengan kata lain, kemungkinan jelajah meningkat seiring bertambahnya usia, mungkin karena jantan muda pertama-tama menjadi jelajah, yang juga berarti bahwa jantan muda tidak dapat dengan mudah beralih ke teritorialitas kecuali mereka tumbuh cukup besar (seperti yang dibahas sebelumnya). Selain itu, karena jantan muda biasanya pertama kali menjadi jelajah, persaingan jantan-jantan untuk wilayah tidak cukup kuat untuk menghasilkan risiko kematian yang lebih tinggi untuk kedua taktik, yang menjelaskan mengapa transisi dari kedua taktik ke kematian secara statistik tidak signifikan. Yang penting, karena pengamatan kami dibatasi hingga usia maksimum 11 bulan, penelitian di masa depan dapat secara khusus mencoba memahami bagaimana ART berkembang di tahap kehidupan selanjutnya (namun perlu dicatat bahwa 10 bulan merupakan usia puncak yang dilaporkan dapat dicapai oleh tikus yang hidup di alam: Berry & Jakobson, 1971 ; Morgan & Bellamy, 1975 ).
~25% teritorial yang beralih ke roaming pada titik mana pun mati dalam waktu satu bulan setelah melakukannya, dan risiko kematian secara konsisten lebih tinggi untuk teritorial (Tabel S2 ). Oleh karena itu, transisi dari mempertahankan teritori ke roaming mungkin mencerminkan efek penuaan pada kondisi tubuh atau peningkatan tingkat stres, yang meningkatkan kematian. Dengan kata lain, analisis kami mengungkapkan bahwa beberapa roamer tetap seperti itu sepanjang hidup mereka dan bahwa beberapa teritorial tidak dapat dengan mudah mengatasi kehilangan kotak sarang mereka (seperti yang dijelaskan di atas). Namun, banyak jantan yang beralih (Gambar S3 ), sebuah indikasi bahwa beberapa individu lebih plastis daripada yang lain. Pada gilirannya, ini mungkin menunjukkan adanya tiga taktik (teritorial, roamer & switcher), sesuatu yang dapat dibahas dalam penelitian di masa depan.
OSR yang condong ke jantan dikaitkan dengan kemungkinan teritorialitas dan kelangsungan hidup yang menurun dalam kedua taktik. Di sini, mungkin saja karena persaingan jantan-jantan meningkat (lebih banyak jantan dewasa dalam populasi; karenanya OSR lebih tinggi), teritori yang tersedia terkuras lebih cepat, membuat pengembaraan tak terelakkan, dan tingkat kematian juga meningkat, dengan efek yang (~tiga kali) lebih kuat di antara pengembara. Oleh karena itu, kami menyarankan bahwa biaya pengembaraan terutama berasal dari persaingan intraseksual yang intens, yang meningkat saat pengembara mengganggu lebih banyak jantan (Jirotkul, 1999a ). Kami percaya bahwa ukuran efek yang relatif lebih kecil dari peningkatan OSR (yang dihasilkan dari lebih banyak jantan dewasa yang hadir) pada teritorial adalah karena jantan ini lebih berat dan paling kompetitif pada titik mana pun, sehingga dapat mengatasi persaingan yang intens. Jika OSR yang lebih tinggi justru disebabkan oleh betina yang kurang aktif secara seksual, ini dapat memilih pengembaraan jantan sebagai cara untuk menemukan betina. Namun, hasil kami tidak menunjukkan tingkat transisi dari teritorialitas ke jelajah saat OSR meningkat; dengan demikian, perubahan dalam OSR terutama mencerminkan fluktuasi dalam persaingan jantan-jantan. Secara komparatif, pada tikus lemur, peralihan antara taktik bergantung pada variasi skala pendek dalam aspek-aspek seperti probabilitas pembuahan, jumlah peluang kawin alternatif, dan rasio jenis kelamin operasional (Eberle & Kappeler, 2004 ).
Peningkatan ukuran populasi berkorelasi dengan kemungkinan yang lebih tinggi untuk berkeliaran tetapi tidak mempengaruhi tingkat transisi langsung. Ini karena ukuran populasi yang lebih tinggi menghabiskan kotak sarang yang tersedia dan jantan baru yang memasuki populasi, karena pengembara tidak dapat menggantikan teritorial. Secara komparatif, kepadatan mempengaruhi ART di seluruh taksa (Brockmann, 2008 ; Forchhammer & Boomsma, 1998 ; Karkarey et al., 2017 ; Wolff, 2008 ) tetapi hanya ketika jumlah jantan melebihi teritori (Forsyth & Montgomerie, 1987 ). Memang, peningkatan kepadatan mengintensifkan persaingan jantan-jantan (dalam penelitian kami ukuran populasi yang lebih tinggi memprediksi kemungkinan tanda gigitan yang lebih tinggi pada tingkat populasi; hasil tidak ditunjukkan), dan ini ditunjukkan secara eksperimental juga (Jirotkul, 1999b ).
Secara keseluruhan, teritorial dan roamer yang menghadapi jumlah persaingan yang sama dan memiliki usia dan berat yang sama berbeda dalam rentang hidup mereka, dengan yang pertama menderita tingkat kematian yang lebih tinggi, mungkin karena biaya yang bergantung pada kondisi dan fisiologis yang terkait dengan mempertahankan wilayah (Ord, 2021 ). Namun, seiring meningkatnya persaingan pria-pria, roamer mati pada tingkat yang lebih tinggi (Gambar 5 ), yang berarti bahwa kepadatan, usia, persaingan pria-pria dan status (berat) memengaruhi peluang menduduki taktik; tetapi hanya dua yang terakhir berinteraksi untuk memengaruhi tingkat transisi (Gambar 5 ).
4.4 Korelasi fisiologis ART
Secara teoritis, jantan harus menyesuaikan ukuran ejakulasi sehubungan dengan risiko dan intensitas persaingan sperma (Montgomerie & Fitzpatrick, 2019 ) dan roamers harus lebih banyak berinvestasi dalam sifat sperma (Dougherty et al., 2022 ; Kustra & Alonzo, 2020 ). Dalam penelitian kami, GSI lebih tinggi untuk roamers meskipun berat testis rata-rata dan jumlah sperma tidak berbeda antara ART, yang terakhir sesuai dengan tinjauan terbaru (Dougherty et al., 2022 ; Kustra & Alonzo, 2020 ; tetapi sebagian besar penelitian vertebrata berasal dari ikan). Hasil penelitian kami menunjukkan bahwa ikan roamer meningkatkan ukuran atau frekuensi ejakulasi (Montgomerie & Fitzpatrick, 2019 ), mirip dengan ikan sneaker yang memiliki testis lebih besar daripada ikan non-sneaker (Dougherty et al., 2022 ; Montgomerie & Fitzpatrick, 2019 ). Ikan teritorial mungkin tidak mampu memenuhi kebutuhan energi yang diperlukan untuk mempertahankan peningkatan ukuran testis saat mereka mengalokasikan energi untuk berpacaran dan mempertahankan wilayah mereka (Montgomerie & Fitzpatrick, 2019 ).
Jumlah sperma dan ukuran tubuh hewan tidak berbeda antara ART jantan. Pada tikus belang, jantan ART berinvestasi secara merata pada sperma (Schradin et al., 2012 ) dan di seluruh taksa, tidak ada perbedaan signifikan pada sifat sperma (kepadatan sperma, volume atau jumlah) (Dougherty et al., 2022 ; Kustra & Alonzo, 2020 ). Dalam penelitian kami, mungkin saja kualitas sperma, komponen cairan mani atau hormon (Kustra & Alonzo, 2020 ), daripada kuantitas sperma, mungkin sedang dalam seleksi, sesuatu yang harus diuji dalam penelitian di masa mendatang. ART jantan dapat berbeda dalam alokasi sperma atau komposisi cairan mani dan bukan pengeluaran sperma (Dougherty et al., 2022 ), sehingga roamer mengalokasikan lebih banyak sperma atau cairan mani per kopulasi untuk meningkatkan kemungkinan pembuahan mereka atau menurunkan kinerja sperma jantan dominan, masing-masing (Dougherty et al., 2022 ). Atau, mungkin saja para pengembara tidak berinvestasi lebih banyak pada sperma karena adanya trade-off (misalnya trade-off energi; diulas dalam Kustra & Alonzo, 2020 ).
5 KESIMPULAN
Secara keseluruhan, di sini kami memberikan bukti kuat bahwa ART pada Mus musculus jantan ada dan bahwa, sementara jantan teritorial memiliki probabilitas reproduksi yang lebih tinggi, ART memiliki kebugaran rata-rata yang sama. Kami selanjutnya menunjukkan ART bervariasi secara signifikan dalam rentang hidup individu sebagai respons terhadap fitur sosial (OSR dan ukuran populasi) dan intrinsik (massa dan usia), dan untuk menunjukkan perbedaan dalam ukuran testis tetapi tidak dalam jumlah sperma. Yang terpenting, hasil kami menunjukkan bahwa tikus jantan menunjukkan ART plastis secara fenotip, yang tetap dapat dijelaskan oleh perbedaan antar individu dari waktu ke waktu (pengulangan). Apakah seleksi seksual lebih banyak bertindak dalam taktik daripada di antara, sebagai akibat dari varians yang lebih tinggi dalam kebugaran di antara jantan yang sebagian besar ditemukan sebagai pengembara, adalah topik yang harus dieksplorasi oleh penelitian di masa mendatang.


