Abstrak
Penelitian pada ikan liar cenderung mengabaikan peran faktor abiotik dalam membentuk perilaku yang terkait dengan keberanian dan eksplorasi. Kelalaian ini dapat terjadi karena variasi skala kecil di lingkungan abiotik tampaknya tidak mungkin memengaruhi perilaku tersebut. Kami menantang asumsi ini melalui penelitian dalam sistem guppy Trinidad ( Poecilia reticulata ). Kami mulai dengan mengukur bagaimana perilaku yang terkait dengan keberanian (waktu di tempat berlindung dan waktu membeku di lapangan terbuka) dan perilaku yang terkait dengan eksplorasi (jumlah kotak kisi yang dilintasi di lapangan terbuka) bervariasi untuk guppy di dalam dan di antara 15 kolam di dua aliran sungai, di mana semua kolam di dalam aliran sungai berada dalam jarak 150 m satu sama lain. Perilaku yang diukur sedikit berbeda antara aliran sungai, namun mereka bervariasi secara dramatis di antara kolam di dalam aliran sungai dan di antara individu di dalam kolam, sehingga menggambarkan bagaimana perilaku tersebut dapat terstruktur pada skala spasial yang sangat kecil. Selanjutnya, kami menilai bagaimana variasi perilaku yang diamati dapat dijelaskan oleh atribut tingkat individu (jenis kelamin dan massa tubuh) dan faktor abiotik tingkat kolam (misalnya suhu dan oksigen terlarut). Atribut tingkat individu hanya menjelaskan sedikit tentang variasi perilaku, meskipun ikan guppy yang lebih kecil menunjukkan perilaku yang sedikit lebih berani. Namun, faktor abiotik di antara kolam cukup informatif. Sebagai contoh yang jelas, ikan guppy dari kolam dengan oksigen terlarut yang lebih sedikit menunjukkan perilaku yang lebih berani dan (mungkin) eksplorasi yang lebih besar. Hasil kami menyoroti pentingnya faktor abiotik dalam membentuk perilaku bahkan pada skala spasial yang kecil.
1. PENDAHULUAN
Dengan pengakuan bahwa variasi fenotipik intraspesifik dapat memiliki efek ekologis dan evolusi yang kuat (Des Roches et al., 2018 ; Hendry, 2017 ), para peneliti semakin tertarik untuk memahami penyebab dan konsekuensi dari variasi antar-individu. Sebagian besar penelitian yang menyelidiki variasi intraspesifik tersebut difokuskan pada perilaku, dengan dua perilaku, keberanian (pengambilan risiko) dan eksplorasi (pergerakan dalam lingkungan baru) (Réale et al., 2007 ), telah menerima perhatian khusus (misalnya Brown et al., 2007 ; Chapman et al., 2011 ; Reader, 2015 ). Perilaku ini dapat memengaruhi proses ekologi mulai dari perolehan informasi dan sumber daya yang berubah (misalnya Patrick et al., 2017 ) hingga keberhasilan dalam interaksi perkawinan (misalnya Ariyomo & Watt, 2013 ; McCowan et al., 2014 ) dan kelangsungan hidup (misalnya Ballew et al., 2017 ; Rödel et al., 2015 ). Di antara individu, keberanian dan eksplorasi dapat sangat bervariasi, terutama di antara individu dari populasi yang berbeda (misalnya Magnhagen et al., 2012 ; Rudh et al., 2013 ). Dengan demikian, menyelidiki bagaimana perilaku berani dan eksploratif dikaitkan dengan faktor-faktor yang berbeda dalam dan di antara populasi dapat memberikan wawasan tentang penyebab perilaku yang penting secara ekologis dan evolusi ini.
Hampir semua pekerjaan yang menyelidiki penyebab variasi dalam perilaku berani dan eksploratif difokuskan pada menghubungkan perilaku ini dengan atribut tingkat individu (misalnya jenis kelamin, ukuran atau ‘tipe’ perilaku individu, yaitu, nilai sifat perilaku rata-rata setiap individu) atau komponen biotik lingkungan yang dialami oleh individu, seperti jumlah dan jenis konspesifik atau heterospesifik (misalnya Archard & Braithwaite, 2011 ; Brown & Braithwaite, 2004 ; Burns et al., 2016 ; Harris et al., 2010 ; Ingley et al., 2014 ; Magnhagen et al., 2012 ; Moran et al., 2016 ; Piyapong et al., 2010 ; Rasmussen & Belk, 2017 ; Ward et al., 2007 ). Konsensus dari kumpulan besar karya ini adalah bahwa atribut tingkat individu dan faktor lingkungan biotik dapat dengan kuat membentuk perilaku berani dan eksploratif di dalam dan di antara populasi alami. Namun, dalam setiap studi tersebut, sejumlah besar perbedaan perilaku yang tidak dapat dijelaskan ada di antara populasi yang berbeda dalam lingkungan biotik yang sama (misalnya Bell et al., 2010 ; Magurran & Seghers, 1994 ). Misalnya, ikan guppy Trinidad ( Poecilia reticulata ) yang dikumpulkan dari situs dengan rezim predasi dan parasitisme yang sama menunjukkan variasi perilaku antar-sungai yang substansial, termasuk dalam keberanian (Jacquin et al., 2016 ). Variasi yang tidak dapat dijelaskan ini menunjukkan bahwa faktor-faktor lain, di luar komponen biotik yang diukur, merupakan kontributor penting terhadap perilaku. Mungkin kita harus melihat lebih dekat pada faktor-faktor abiotik.
Beberapa penelitian telah menganalisis hubungan antara faktor abiotik dan keberanian atau perilaku eksploratif, dan beberapa penelitian tersebut menemukan pengaruh yang kuat (misalnya suhu, sulfida, oksigen terlarut, kecepatan aliran air; Culumber, 2020 ; Riesch et al., 2009 ; Sommer-Trembo et al., 2017 ; Tang & Fu, 2021 ). Dengan demikian, faktor lingkungan abiotik dapat berkontribusi terhadap variasi yang tidak dapat dijelaskan dalam lingkungan yang secara biotik serupa. Yang menarik, faktor abiotik dapat bervariasi pada skala spasial yang berkisar dari kecil (misalnya dua kolam yang dipisahkan oleh riffle dangkal di dalam aliran sungai) hingga besar (misalnya dua aliran sungai yang berbeda), namun studi relevan dari alam biasanya hanya berfokus pada skala yang lebih besar (misalnya di antara populasi: Sommer-Trembo et al., 2017 ; Tang & Fu, 2021 ) atau secara sengaja membesar-besarkan faktor abiotik untuk melampaui kondisi lingkungan yang tidak terganggu (misalnya tekanan termal atau salinitas; Culumber, 2020 ; Leite et al., 2019 , 2022 ). Akibatnya, kita masih memiliki sedikit pemahaman tentang bagaimana variasi abiotik alami pada skala spasial kecil dapat berkontribusi pada variasi perilaku. Untuk mengatasi kesenjangan penelitian ini, kami berusaha menjelaskan variasi perilaku melalui pengukuran faktor abiotik dalam lingkungan yang secara biotik serupa. Artinya, kami secara sengaja mengendalikan beberapa sumber variasi biotik yang ‘diketahui penting’ untuk menilai apakah faktor abiotik dapat mengatasi variasi yang tidak dapat dijelaskan.
1.1 Studi kami
Kami fokus pada perilaku berani dan eksploratif dari ikan guppy Trinidad Poecilia reticulata (Peters 1859) yang terungkap melalui pengamatan dalam uji lapangan terbuka, uji yang mapan untuk mengukur variasi dalam perilaku ini pada ikan guppy (Burns, 2008 ) dan spesies ikan lainnya (Toms et al., 2010 ). Beberapa penelitian yang mengukur perilaku berani dan eksploratif melakukannya dengan tujuan mempelajari perbedaan perilaku yang dapat diulang, yang sering dipandang sebagai ‘kepribadian’ atau ‘temperamen’; Réale et al., 2007 ). Meskipun penelitian sebelumnya tentang perilaku ini pada ikan guppy sering menemukan bahwa mereka dapat diulang (Tabel S1 ), minat kami adalah menjelaskan variasi dalam perkiraan titik ‘perilaku’ bukan dalam upaya untuk menyimpulkan variasi dalam ‘kepribadian’. Artinya, ukuran perilaku berani dan eksploratif kami mungkin mencakup efek perbedaan perilaku antar individu, serta respons plastik dalam individu terhadap lingkungan tempat perilaku tersebut dikumpulkan, yang mungkin kemudian juga dipengaruhi oleh proses eksperimen (misalnya stres dan penanganan).
Uji perilaku dilakukan pada semua individu yang dikumpulkan dari 15 kolam (yaitu mikrohabitat dengan aliran gen terbatas musiman) yang didistribusikan melintasi dua aliran sungai. Dalam aliran sungai tertentu, kolam dipisahkan oleh jarak sesedikit 0,75 m dan tidak lebih dari 150 m. Kedua aliran sungai berada dalam DAS yang sama (Gambar 1 ), dan secara geografis dekat tetapi mengandung populasi ikan guppy yang secara fenotip dan genetik berbeda (Blondel et al., 2019 ). Untuk fokus pada lingkungan yang secara biotik serupa, kedua aliran sungai berisi populasi ikan guppy ‘predasi rendah’, di mana predator utamanya adalah Anablepsoides hartii yang terbatas pada celah yang hanya memakan ikan guppy kecil (Endler, 1978 ; McKellar et al., 2009 ; Seghers, 1973 ). Selain itu, aliran sungai yang serupa di sungai yang sama dapat memiliki tingkat parasit ikan guppy Gyrodactylus spp yang sangat rendah. (Gotanda dkk., 2013 ; Pérez-Jvostov dkk., 2012 ).
Secara analitis, pertama-tama kami menetapkan bagaimana perilaku berani dan eksploratif bervariasi pada tiga skala spasial. Pertama, kami menyelidiki variasi di antara individu dalam kelompok tertentu. Kedua, kami menyelidiki variasi di antara kelompok dalam aliran tertentu. Ketiga, kami menyelidiki variasi antara kedua aliran. Selanjutnya kami memeriksa faktor-faktor yang terkait dengan variasi perilaku pada skala spasial ini. Dalam kelompok, kami berfokus pada jenis kelamin dan ukuran, dua atribut tingkat individu yang dapat berkontribusi pada variasi perilaku di antara individu. Di antara kelompok, kami berfokus pada beberapa faktor lingkungan abiotik, seperti oksigen terlarut (mg/L) dan suhu (°C), yang secara masuk akal terkait dengan perilaku yang diukur (misalnya Culumber, 2020 ; Sommer-Trembo et al., 2017 ).
2 BAHAN DAN METODE
2.1 Pemilihan kolam dan pengumpulan data ekologi
Studi kami difokuskan pada total 15 kolam yang dipilih di dua aliran sungai (aliran 1, n = 7; aliran 2, n = 8) di Sungai Marianne di Trinidad (Gambar 1 ). Kedua aliran sungai ini mewakili kelompok genetik yang berbeda (Blondel et al., 2019 ), meskipun keduanya masih merupakan bagian dari sungai yang sama. Pekerjaan kami dilakukan selama musim air rendah/kering (Februari 2020), yang memastikan bahwa kolam-kolam tersebut sebagian besar atau seluruhnya terisolasi satu sama lain. Sebelum mengumpulkan ikan guppy, kami mencatat variabel ekologi tingkat kolam yang dapat memengaruhi perilaku mereka. Pertama, untuk setiap kolam, kami menggunakan pita pengukur atau pipa PVC bertanda untuk mengukur lebar maksimum (m), panjang maksimum (m), kedalaman maksimum (m), dan kedalaman rata-rata (m); pengukuran terakhir ini diperoleh dengan melakukan pengukuran kedalaman setiap 50 cm di sepanjang bagian terpanjang dan terluas kolam. Dimensi linier ini kemudian digunakan untuk memperkirakan luas permukaan (panjang × lebar dalam m 2 ) dan volume (luas permukaan × kedalaman rata-rata dalam m 3 ) setiap kolam. Kedua, kami menggunakan probe Yellow Springs Instrument (YSI) (model 10.102.030; Yellow Springs Inc.) untuk mencatat pH, suhu (°C) dan konsentrasi oksigen terlarut (mg/L) air. Kami juga mencatat konduktansi spesifik (μS/cm) dengan YSI, yang memberikan konduktivitas yang distandarisasi oleh suhu. Akhirnya, kami menggunakan densiometer bulat cekung untuk mencatat tutupan kanopi di atas setiap kolam.
Kami kembali ke kolam 72 jam setelah variabel fisik dan ekologi di atas diukur untuk mengumpulkan ikan guppy. Dengan menggunakan jaring kupu-kupu, kami mencoba menangkap semua ikan guppy dari setiap kolam, yang biasanya memungkinkan karena tingkat penangkapan sangat tinggi di kolam kecil di musim kemarau, seperti yang ditunjukkan oleh banyak studi penandaan-penangkapan kembali (misalnya Bryant & Reznick, 2004 ; Reznick et al., 1996 ; Weese et al., 2010 ). Untuk memastikan tingkat penangkapan yang tinggi dalam studi kami, kami menunggu setidaknya 2 menit setelah ikan guppy terakhir yang ditangkap. Jika ada ikan guppy lain yang terlihat, ia ditangkap dan penghitung waktu 2 menit disetel ulang. Proses ini diulang hingga tidak ada lagi ikan guppy yang terlihat. Meskipun ada kemungkinan kami gagal menangkap sejumlah ikan guppy penghuni, setidaknya kami menangkap sebagian besar dari mereka, menghasilkan total 303 ikan guppy dewasa (aliran 1, n = 126; aliran 2, n = 177) (lihat Tabel S2 untuk ukuran sampel spesifik kolam).
Pada hari yang sama saat ikan guppy dikumpulkan (aliran 1, 17 Februari 2020; aliran 2, 18 Februari 2020), semua ikan diangkut dalam botol plastik bersih berukuran 2 L dengan mobil ke Stasiun Penelitian Tropis William Beebe di Lembah Arima, tempat mereka ditempatkan di akuarium khusus kolam yang diisi dengan air hujan yang dikumpulkan di stasiun penelitian. Saat berada di laboratorium, ikan guppy diberi makan serpihan ikan (TetraMin Tropical Flakes) setiap hari, air akuarium diolah dengan API Stresscoat dan API Quick Start, dan penggantian air dilakukan setiap beberapa hari. Ikan guppy ditahan dalam kondisi ini setidaknya selama 2 hari, dengan waktu penahanan maksimum 18 hari, sebelum digunakan dalam uji perilaku. Lamanya waktu bervariasi dengan cara ini karena total waktu pemrosesan yang dibutuhkan untuk semua ikan sebelum dilepaskan kembali secara bersamaan ke sungai.
2.2 Uji perilaku
Pengujian perilaku dilakukan di akuarium 72 L (lebar 30,5 cm, kedalaman 38,5 cm, panjang 61 cm) dengan kisi 5 × 5 cm yang digambar di bagian bawah. Kerikil akuarium berwarna merah-coklat dan kuning disebarkan tipis di bagian bawah untuk mendekati beberapa warna substrat alami di sungai Trinidad. Air dipertahankan pada kedalaman 6 cm dan diganti dua kali sehari: sebelum percobaan pertama dan setelah sekitar setengah percobaan selesai. Tangki diterangi dengan cahaya alami dari jendela besar dan pencahayaan dalam ruangan di stasiun lapangan. Tanaman buatan ditempatkan di sudut setiap akuarium untuk memberikan kesempatan bagi ikan untuk mencari perlindungan selama percobaan. Perilaku (detail di bawah) dinilai secara langsung oleh seorang pengamat. Lembaran plastik biru buram ditempatkan di antara tangki dan tubuh pengamat untuk meminimalkan gangguan pada ikan sambil memungkinkan pengamat untuk tetap melihat ikan dari atas lembaran tersebut. Pengujian dilakukan antara matahari terbit dan terbenam agar sesuai dengan periode aktivitas diurnal.
Untuk setiap percobaan, seekor ikan ditempatkan di arena percobaan di dalam wadah plastik bening, yang dibuat dengan merekatkan dua wadah makanan mini bening. Wadah tersebut ditempatkan di sudut akuarium pada sisi pendek yang sama dengan tempat berlindung tanaman (tempat berlindung berada di satu sudut dan wadah ditempatkan di sudut lainnya). Ikan-ikan tersebut dibiarkan di dalam tabung penampung selama periode aklimatisasi selama 3 menit, setelah itu tabung penampung diangkat perlahan dengan tangan oleh pengamat dan percobaan segera dimulai. Setiap percobaan kemudian berlangsung selama 5 menit, selama waktu tersebut keempat perilaku tersebut direkam. Pengukuran perilaku berani kami adalah penggunaan tempat berlindung (waktu, dalam detik, yang dihabiskan ikan di tempat berlindung tanaman buatan), kejadian pembekuan (jumlah kejadian saat ikan bergerak dan kemudian berhenti) dan total waktu pembekuan (waktu, dalam detik, saat ikan tidak bergerak). Pengukuran perilaku eksplorasi kami adalah jumlah kotak yang dilintasi di lapangan terbuka yang baru. Proksi-proksi ini dipilih karena penelitian-penelitian sebelumnya mengonfirmasikan penggunaan proksi-proksi tersebut untuk mempelajari variasi dalam perilaku berani dan eksploratif (misalnya Carlson & Langkilde, 2013 ; Diaz Pauli et al., 2019 ). Kami menggunakan perangkat lunak perilaku BORIS v 7.9.7 (Friard & Gamba, 2016 ) untuk merekam secara langsung waktu yang dihabiskan di tempat perlindungan, total waktu pembekuan, jumlah kejadian pembekuan, dan jumlah kotak kisi yang dilintasi. Setelah menyelesaikan uji perilaku, ikan-ikan tersebut dibius ringan menggunakan tricaine methanesulfonate (MS-222) dan dipindai secara visual untuk mencari ektoparasit di bawah mikroskop bedah Zeiss (kami tidak menemukan apa pun, meskipun Gyrodactylus terkadang dapat terlepas selama penangkapan dan pengangkutan ikan dan penelitian-penelitian lain telah melaporkan Gyrodactylus di Marianne; Gotanda et al., 2013 ). Jenis kelamin dan massa (g) dicatat. Hanya data untuk ikan guppy dewasa yang disertakan dalam naskah ini.
Untuk menentukan validitas proksi perilaku kami, kami menggunakan paket ‘factoextra’ v.1.0.7 di lingkungan R (Kassambara & Mundt, 2020 ; Tim Inti R, 2024 v 4.3.0) untuk melakukan analisis komponen utama (PCA) pada matriks korelasi data perilaku dan empat proksi perilaku (penggunaan tempat perlindungan, jumlah kejadian pembekuan, total waktu pembekuan, dan jumlah kotak yang dilintasi). Komponen utama pertama (PC1) menjelaskan 41% dari total varians dan dikaitkan dengan penggunaan tempat perlindungan (kontribusi 38% untuk PC1), jumlah kejadian pembekuan (kontribusi 31% untuk PC1) dan total waktu pembekuan (kontribusi 31% untuk PC1), yang menunjukkan bahwa PC ini terdiri dari perilaku berani. PC2 menjelaskan 35% dari total varians dan sebagian besar dikaitkan dengan jumlah kotak yang dilintasi (kontribusi 64% untuk PC2), yang menunjukkan bahwa PC ini terdiri dari perilaku eksploratif.
Dalam analisis berikut, alih-alih menggunakan dua PC, kami malah menggunakan skor perilaku mentah sebagai proksi untuk perilaku berani dan perilaku eksploratif. Sebagai proksi untuk perilaku berani, kami membuat variabel baru yang mewakili jumlah waktu yang dihabiskan dalam keadaan beku di lapangan terbuka dan waktu yang dihabiskan di bawah perlindungan. Variabel baru ini diperlukan karena kedua ukuran perilaku ini tidak independen. Misalnya, jika seekor ikan menghabiskan sebagian besar percobaan di bawah perlindungan, yang menunjukkan perilaku yang lebih pemalu, individu yang sama kemungkinan akan memiliki skor waktu beku yang lebih rendah (karena mereka menghabiskan lebih sedikit waktu di lapangan terbuka untuk mengamati perilaku tersebut), yang menunjukkan perilaku yang lebih berani. Sebagai proksi untuk perilaku eksploratif, kami menggunakan jumlah kotak yang disilangkan di lapangan terbuka. Kami memilih perilaku ini karena mudah diubah untuk memenuhi asumsi model linier (lihat di bawah), sedangkan PC tidak mudah diubah untuk memenuhi asumsi linearitas dan penggunaannya dalam model dapat berisiko meningkatkan kesalahan tipe 1. Korelasi Pearson antara kedua proksi perilaku ini adalah r = -0,67. Korelasinya negatif karena nilai yang lebih rendah untuk proksi keberanian kami mencerminkan perilaku yang lebih berani (misalnya lebih sedikit waktu membeku dan berlindung merupakan indikasi perilaku yang lebih berani). Oleh karena itu, individu yang lebih berani juga lebih eksploratif.
2.3 Analisis Statistik
Kami memulai penyelidikan kami tentang struktur spasial perilaku berani (jumlah waktu yang dihabiskan membeku di lapangan terbuka dan di bawah perlindungan tanaman) dan perilaku eksploratori (jumlah kotak yang dilintasi di lapangan terbuka) dengan menggunakan ANOVA bersarang (SPSS v.29.0.0.0) untuk membagi total variasi dalam perilaku berani dan eksploratori di antara aliran dan kolam yang bersarang di dalam aliran. Estimasi eta-kuadrat parsial (proporsi dari total varians, η p 2 ) yang dijelaskan oleh setiap efek acak memungkinkan kami untuk menjawab bagaimana perilaku berani dan eksploratori terstruktur pada skala ini. Selanjutnya kami menyelidiki bagaimana struktur spasial perilaku berani dan eksploratori dapat dijelaskan oleh variabel tingkat individu dan tingkat kolam.
Variabel tingkat individu dan tingkat kolam dianalisis dengan model efek campuran linier (LMM) menggunakan paket R ‘lme4’ (Bates et al., 2015 ), dengan perilaku berani atau eksploratif sebagai variabel dependen. Efek tetap dalam model ini mencakup jenis kelamin dan massa, serta efek independen dari beberapa variabel ekologi (diuraikan di bawah). Kami juga menyertakan waktu dalam sehari (pagi/sore) dari pengujian perilaku sebagai kovariat untuk memperhitungkan variasi dalam perilaku yang dapat muncul sepanjang hari karena waktu dalam sehari dapat memengaruhi beberapa perilaku ikan guppy (misalnya O’Neill et al., 2019 ). Kami menyertakan efek acak untuk kolam yang bersarang di dalam sungai untuk memperhitungkan penataan spasial yang tidak independen. Awalnya, karena ikan guppy jantan dan betina berbeda dalam ukuran, kami menyertakan interaksi jenis kelamin berdasarkan massa, tetapi model perilaku tebal tidak konvergen dengan interaksi yang disertakan ini dan interaksi tersebut tidak signifikan dalam model perilaku eksploratif ( χ2 = 0,76, df = 1, p = 0,38) sehingga dihilangkan dari kedua model untuk meningkatkan kesesuaian model (kami malah mempertimbangkan efek independen dari jenis kelamin dan massa) (Engqvist, 2005 ). Variabel lingkungan abiotik dalam model tersebut adalah oksigen terlarut (mg/L), konduktansi spesifik (μS/cm), luas permukaan (m2 ) , suhu (°C), tutupan tajuk (kotak yang tercakup pada densiometer) dan volume kolam (m3 ) . Kami mengecualikan kedalaman rata-rata (m) dan pH karena korelasi yang tinggi (yaitu r > 0,8) dengan volume ( r = 0,93) dan konduktansi spesifik ( r = 0,83), masing-masing (Tabel S3 ). Kami juga mengecualikan kedalaman maksimum (m) karena penyertaannya menghasilkan estimasi varians nol untuk efek acak tingkat kolam (yaitu secara efektif hanya mempertimbangkan efek aliran). Semua prediktor kontinu (dalam model ini dan yang dijelaskan di bawah) distandarisasi melalui konversi ke z -skor sebelum penyertaannya dalam model. Diagnostik model dinilai menggunakan paket R ‘DHARMa’ (Hartig, 2022 ). Perilaku tebal ditransformasikan menjadi akar pangkat tiga dan perilaku eksploratori ditransformasikan menjadi akar pangkat dua untuk memenuhi asumsi linearitas. Tidak ada asumsi lain yang dilanggar. Menggunakan paket R ‘car’, kami menghitung nilai p untuk setiap LMM dengan jumlah kuadrat tipe II.
Untuk menghitung ukuran efek untuk jenis kelamin dan massa, kami menggunakan paket ‘r2glmm’ (Jaeger, 2017 ) untuk mengekstrak nilai R 2 semi-parsial untuk setiap efek tetap dari LMM. Mirip seperti eta-kuadrat parsial yang berasal dari ANOVA atau model linear (LM), R 2 semi-parsial memperkirakan varians yang dijelaskan untuk setiap prediktor dalam LMM (Jaeger et al., 2017 ), yang memberikan wawasan tentang jumlah variasi dalam variabel respons (yaitu perilaku) yang dijelaskan oleh efek tetap (yaitu atribut tingkat individu).
Kami menggunakan pendekatan yang berbeda untuk menghitung ukuran efek untuk variabel ekologi. Karena tingkat pengumpulan data ekologi (yaitu tingkat kumpulan) berbeda dari tingkat pengumpulan data perilaku (yaitu tingkat individu), penghitungan R 2 untuk variabel ekologi menggunakan LMM di atas menghasilkan ukuran efek yang sangat rendah (lihat materi suplemen untuk estimasi ukuran efek dan plot yang dibangun menggunakan hasil LMM; Gambar S1 dan S2 ). Memang, nilai tunggal sebagai variabel independen tidak dapat menjelaskan variasi dalam variabel dependen; untuk menilai nilai R 2 dari LMM dengan cara yang bermakna secara biologis akan memerlukan pengambilan sampel skala yang lebih halus untuk mengumpulkan data lingkungan mikro untuk setiap individu. Oleh karena itu, daripada menilai seberapa besar variasi dalam perilaku individu dapat dijelaskan oleh efek tetap (seperti di atas), kami malah menilai seberapa besar variasi dalam perilaku tingkat kumpulan dapat dijelaskan oleh efek tetap. Untuk menghitung ukuran efek ini, kami membangun dua model linier (LM). Variabel respons dalam model ini adalah skor rata-rata untuk perilaku berani atau eksploratif untuk kumpulan tertentu ( n = 15). Efek tetap adalah efek independen dari semua variabel lingkungan abiotik yang disertakan dalam model di atas. Untuk mengendalikan variasi antar-aliran, kami juga menyertakan aliran sebagai kovariat tambahan dalam model ini. Kami mengekstraksi eta-kuadrat parsial menggunakan paket R ‘effectsize’ dengan jumlah kuadrat tipe 2 (Ben-Shachar et al., 2020 ). Perilaku eksplorasi ditransformasikan ke dalam logaritma untuk memenuhi asumsi linearitas.
Meskipun jenis kelamin dan massa adalah atribut tingkat individu, dan memperkirakan R 2 dari LMM karenanya tepat, kami melakukan analisis sensitivitas untuk memastikan bahwa ukuran efek yang dihitung dari LM tidak berbeda secara substansial dari yang diperkirakan dari LMM. Oleh karena itu, kami juga membangun dua LM tambahan, dengan perilaku tebal atau eksploratif sebagai variabel respons ( n = 303), di mana jenis kelamin, massa, dan aliran adalah satu-satunya efek tetap. Perilaku tebal ditransformasikan akar pangkat tiga untuk memenuhi asumsi kenormalan dan linearitas. Kami kembali mengekstraksi eta-kuadrat parsial menggunakan paket R ‘effectsize’ dengan jumlah kuadrat tipe II. Ukuran efek dari LM ini tidak berbeda secara kualitatif dari yang diekstraksi dari LMM di atas, sehingga hanya ukuran efek dari LMM yang disajikan dalam teks (Tabel S4 ).
2.4 Pernyataan etika
Izin untuk melaksanakan pekerjaan ini datang dari McGill University Animal Care (AUP 8058).
3 HASIL
Saat memeriksa struktur spasial perilaku berani dan eksploratif, variasi yang cukup besar terbukti di antara kumpulan di dalam aliran sungai, tetapi tidak antar aliran sungai, untuk kedua perilaku ( η p 2 untuk kumpulan di dalam aliran sungai: berani = 9,0%, p = 0,01; eksploratif = 12%, p < 0,001; η p 2 untuk aliran sungai: berani = 2,0%, p = 0,50; eksploratif = 0%, p = 0,99) (Tabel 1 dan Gambar 2 ).
| df | F | P | saya p 2 | |||
|---|---|---|---|---|---|---|
| Perilaku yang berani | Mencegat | Hipotesa | 1.00 | 360.64 | 0,03 | 1.00 |
| Kesalahan | 1.00 | |||||
| Sungai kecil | Hipotesa | 1.00 | 0.48 | 0,50 | 0,02 | |
| Kesalahan | Tanggal 21.16 | |||||
| Kolam (aliran sungai) | Hipotesa | Jam 13.00 | 2.24 | 0,01 | 0,09 | |
| Kesalahan | 288.00 | |||||
| Perilaku eksploratif | Mencegat | Hipotesa | 1.00 | 186.140,64 | <0,001 | 1.00 |
| Kesalahan | 2.44 | |||||
| Sungai kecil | Hipotesa | 1.00 | 0.00 | 0,99 | 0.00 | |
| Kesalahan | tanggal 19.06 | |||||
| Kolam (aliran sungai) | Hipotesa | Jam 13.00 | 2.94 | <0,001 | 0.12 | |
| Kesalahan | 288.00 | |||||
Catatan : Kumpulan yang dimasukkan dalam aliran disertakan sebagai efek acak.
Untuk variabel tingkat individu, jenis kelamin menjelaskan 0% varians pada kedua perilaku (Gambar 3 ). Sementara massa juga menjelaskan 0% varians pada perilaku eksplorasi, massa menjelaskan 3% (95% CI 0,00–0,08) varians pada perilaku berani dan individu yang lebih kecil menunjukkan perilaku yang lebih berani ( χ 2 1 = 9,3, p < 0,01) (Tabel 2 dan Gambar 3 ).
| Perilaku | Variabel ‘tipe’ | Ketentuan | χ 2 | df | P |
|---|---|---|---|---|---|
| Berani | Faktor tingkat individu | Seks | 0,01 | 1 | 0,94 |
| Massa | 9.26 | 1 | <0,01 | ||
| Faktor tingkat kolam | Oksigen terlarut (mg/L) | 4.73 | 1 | 0,03 | |
| Luas permukaan (m 2 ) | 0.44 | 1 | 0.51 | ||
| Suhu (°C) | 0,79 | 1 | 0.37 | ||
| Konduktansi spesifik (μS/cm) | 4.84 | 1 | 0,03 | ||
| Volume ( m3 ) | 0.52 | 1 | 0.47 | ||
| Penutup kanopi (kotak yang tertutup pada densiometer) | 0,65 | 1 | 0.42 | ||
| Kovariat | Waktu dalam sehari | 4.76 | 1 | 0,03 | |
| Penyelidikan | Faktor tingkat individu | Seks | 0.43 | 1 | 0.51 |
| Massa | 0.64 | 1 | 0.42 | ||
| Faktor tingkat kolam | Oksigen terlarut (mg/L) | 3.55 | 1 | 0,06 | |
| Luas permukaan (m 2 ) | 0.56 | 1 | 0,45 | ||
| Suhu (°C) | 0.31 | 1 | 0,58 | ||
| Konduktansi spesifik (μS/cm) | 0,05 | 1 | 0.82 | ||
| Volume ( m3 ) | 0,04 | 1 | 0.84 | ||
| Penutup kanopi (kotak yang tertutup pada densiometer) | 0.00 | 1 | 0,95 | ||
| Kovariat | Waktu dalam sehari | Tanggal 12.23 | 1 | <0,001 |
Catatan : Kolam yang bersarang di aliran sungai disertakan sebagai efek acak. Perilaku yang dicetak tebal (jumlah waktu yang dihabiskan dalam keadaan beku di lapangan terbuka dan di bawah naungan tanaman) diubah menjadi akar pangkat tiga dan perilaku eksploratif (jumlah kotak yang dilintasi di lapangan terbuka) diubah menjadi akar pangkat dua untuk memenuhi asumsi linearitas.

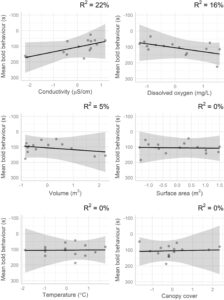
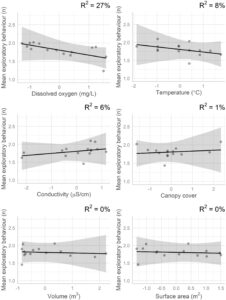
Untuk variabel tingkat kolam abiotik, pertama mengenai perilaku berani, individu di kolam dengan konduktansi spesifik yang lebih tinggi ( χ 2 1 = 4,84, p = 0,03) dan di kolam dengan oksigen terlarut yang lebih rendah ( χ 2 1 = 4,73, p = 0,03) menunjukkan perilaku yang lebih berani. Kami menemukan 22% dari variasi dalam perilaku berani rata-rata tingkat kolam dikaitkan dengan konduktansi spesifik dan 16% dikaitkan dengan oksigen terlarut (Gambar 4 ). Meskipun tidak signifikan secara statistik dalam LMM, volume (m 3 ) juga memiliki efek kecil pada perilaku berani rata-rata tingkat kolam ( η p 2 = 0,05). Aliran menjelaskan 11% dari varians dalam perilaku berani ketika dimasukkan sebagai kovariat dalam model ini. Mengenai perilaku eksplorasi, meskipun hanya signifikan secara marjinal, tren tersebut menunjukkan bahwa individu lebih banyak mengeksplorasi di lingkungan dengan oksigen terlarut yang lebih sedikit ( χ21 = 3,55 , p = 0,06) dan, konsisten dengan temuan ini, oksigen terlarut menjelaskan 27% dari varians dalam perilaku eksplorasi rata-rata tingkat kolam (Gambar 5 ). Suhu dan konduktansi spesifik juga memiliki efek kecil pada perilaku eksplorasi rata-rata tingkat kolam ( ηp2 = 0,08 dan 0,06, berturut- turut ). Aliran menjelaskan 14% dari varians dalam perilaku eksplorasi rata-rata tingkat kolam. Semua variabel ekologi lainnya memiliki efek kecil ( ηp2< 0,02 ) pada perilaku rata-rata tingkat kolam dan tidak ada variabel tingkat kolam lainnya yang memiliki efek signifikan secara statistik pada perilaku (Tabel 2 ). Di antara semua kolam di sungai, rata-rata ± simpangan baku untuk konduktansi spesifik (μS/cm) adalah 106,03 ± 9,48 di sungai 1 dan 84,61 ± 18,30 di sungai 2, rata-rata oksigen terlarut (mg/L) adalah 4,16 ± 0,82 di sungai 1 dan 4,06 ± 0,86 di sungai 2, suhu rata-rata (°C) adalah 22,52 ± 0,17 di sungai 1 dan 22,89 ± 0,13 di sungai 2, dan volume rata-rata (m 3 ) adalah 0,20 ± 0,25 di sungai 1 dan 0,31 ± 0,33 di sungai 2 (Tabel 3 ). Terakhir, waktu dalam sehari juga signifikan dalam kedua model, mengonfirmasi nilai penyertaan istilah ini sebagai kovariat, dengan individu menunjukkan perilaku lebih berani di pagi hari dan perilaku lebih eksploratif di sore hari (Tabel 2 ).